7.2%萎锈灵和40%拌种灵包衣对棉花幼苗冷胁迫生理生化特征的功效比较
2024-02-23周小云张军高龚静云周广威张少民
周小云,雷 斌,张军高,梁 晶,龚静云,周广威,张少民,李 进
(新疆农业科学院核技术生物技术研究所/新疆作物化学调控工程技术研究中心,乌鲁木齐 830091)
0 引 言
【研究意义】棉花是新疆的主要经济作物之一,随着“矮、密、早、膜、滴”植棉技术的推广应用,2022年新疆棉花总产量为539.1×104t,占全国棉花总产量的90.2%[1]。但是,在棉花播种季节中常常遭遇阴雨、寒潮、倒春寒等低温气象灾害,致使地表温度远低于棉花种子萌发和幼苗生长温度的最低点,甚至降低到0℃以下,影响棉花种子萌发,导致每年有不同程度的烂种、烂芽、烂根甚至死苗现象发生,棉花播种后出苗难,一播全苗更难,严重的会导致田间缺苗断垄甚至毁种重播,即使及时补种或移苗补栽,由于生育期不一致,先出苗的长势比补种的长势好,补种的苗往往跟不上群体的发育速度,造成补种的植株弱小生长发育缓慢,即使中后期加强管理也难以弥补前期生长不良造成的损失,小苗无法获得足够的养分,会出现空棵或晚熟,影响棉花产量和品质。因此,播种季节中低温气象灾害对棉花生产影响较大。而种子包衣技术是在浸种和拌种的基础上发展起来的一项植保技术,具有综合防治、低毒高效、省种省药的特点,随着种子萌发、出苗和生长过程,种衣剂里有效成分逐渐被植株根系吸收并传导到幼苗植株各部位后对种子及幼苗根系起到保护作用,对其周围附近病菌及地下、地上害虫起到防治作用。采用种衣剂包衣棉种可有效地解决早春棉花出苗的问题。但是,在低温冷胁迫危害较重情况下,仍然难以确保田间较高保苗率,需要研究提高棉花苗期适应冷胁迫能力的新产品来获得均匀的田间出苗和较高出苗率,保苗率达到计划种植密度。【前人研究进展】萎锈灵(carboxin)和拌种灵(amicarthiazol)两者均是羧基酰胺类杀菌剂,其活性结构相似,均具有丁烯酰胺类结构,两者区别在于萎锈灵的分子结构中含有噁噻因环,而拌种灵的分子结构中含有噻唑基团。由于丁烯酰胺类化合物的脂溶性较高,易于穿透病菌细胞壁进入体内到达作用靶标,因此丁烯酰胺结构在对担子菌真菌具有高效内吸活性方面起着重要的作用[2,3]。另外,与不同病原菌琥珀酸脱氢酶铁-硫蛋白的S3中心的空间构象有关,活性结构结合部位与其中心的氨基酸残基共价结合,导致酶活性中心的构象发生改变,阻止电子从琥珀酸向辅酶Q的传递,进一步阻止其中心的氧化作用,抑制细胞的呼吸作用,致使病原菌的生长受到抑制作用[4,5]。萎锈灵和拌种灵两者在植株体内均具有良好的内吸传导性,对棉花立枯病有较好防治作用[6,7]。但是,萎锈灵包衣棉种不仅能促进棉花生长并具有抗氧化防御机制,还能增强棉苗适应冷胁迫的能力[8,9]。【本研究切入点】萎锈灵和拌种灵均是一种具有内吸作用的杀菌剂,可用于防治麦类锈病、棉花立枯病等,且防效较好,持效性长。但是萎锈灵和拌种灵包衣对棉幼苗冷胁迫生理生化特征的比较研究未见相关报道。需研究比较具有活性结构相似的2种杀菌剂萎锈灵和拌种灵包衣棉种提高棉种适应冷胁迫能力功效。【拟解决的关键问题】比较具有生物活性结构相似的拌种灵与萎锈灵2种杀菌剂包衣诱导棉苗适应冷胁迫的功效,分析2种杀菌剂包衣提高棉花适应冷胁迫的能力差异,研究其生理作用机制,为开发棉花适应冷胁迫环境的包衣剂新产品提供理论依据。
1 材料与方法
1.1 材 料
选用棉花品种为新陆早50号(新疆农业科学院经济作物研究所提供)。
药剂:99.00%拌种灵原药和99.00%萎锈灵原药。(1)基础种衣剂:只包含成膜剂、表面活性剂等种衣剂惰性成分,不包含杀菌剂农药活性成分。(2)拌种灵种衣剂:参考王知惠等[10]在基础种衣剂基础上添加拌种灵,制成拌种灵含量为40%的种衣剂。(3)萎锈灵种衣剂:参考周小云等[8]在基础种衣剂基础上添加萎锈灵,制成萎锈灵含量7.2%的种衣剂。(1)、(2)、和(3)均由新疆农业科学院核技术生物技术研究所生物调控室自行研制。
1.2 方 法
1.2.1 试验设计
挑选籽粒饱满的棉花种子,经1%次氯酸钠消毒后灭菌蒸馏水冲洗干净并晾干。以基础种衣剂包衣处理为对照,将药剂分别添加到基础种衣剂中,制成以上不同的种衣剂,试验共设2个包衣处理:拌种灵种衣剂包衣处理和萎锈灵种衣剂包衣处理;分别按药种重量比1∶100进行包衣为对照,依次记为CK、T1、T2。将包衣的棉花种子放在通风干燥处阴干,进行种子萌发和幼苗培养。
1.2.2 测定指标
1.2.2.1 种子萌发及冷胁迫处理
细砂经水洗后干热灭菌晾至室温,添加13%的灭菌水混匀,装入发芽盒中构建发芽床。每个发芽盒沙床上等距离摆放不同处理的30粒种子;参照李海明等[11]方法在室内人工智能气候箱(4℃/16℃,12 h/12 h)模拟冷胁迫处理环境进行种子萌发。试验过程中,从第4 d开始每48 h观察记录种子萌发情况,第14 d结束萌发,采集生长一致的子叶用于测定相关生理生化指标,从各处理中均随机选出10株棉苗洗净后,经杀青后烘干至质量恒定,电子分析天平称量发芽前种子干重、发芽后的种皮和种壳干重以及幼苗鲜/干质量。并将冷胁迫处理后25℃环境进行恢复生长7 d后,对不同伤害程度的棉花幼苗叶片冷胁迫症状进行调查。
1.2.2.2 计算
参照高荣岐等[12]和施成晓等[13]方法计算发芽势(Germination potential,GP)、发芽率(Germination rate,GR)、发芽指数(Germination index,GI)、活力指数(Vigor index,VI)、鲜重(Fresh weight,FW)、干重(Dry weight,DW)、干物质转移率(Dry matter conversion rate,DMCR)和呼吸消耗量(Respiratory consumption,RC)等种子萌发相关指标。
参照王学奎等[14]编写的《生理生化实验原理和技术》测定棉苗相关生理生化指标,包括相对电导率(Relative conductivity,REC)、丙二醛(Malondialdehyde,MDA)含量、可溶性蛋白(Soluble protein,SP)含量、可溶性糖(Soluble sugar,SS)含量、超氧化物歧化酶(Superoxide dismutase,SOD)活性、过氧化物酶(Peroxidase,POD)活性和过氧化氢酶(Catalase,CAT)活性。
参照王俊娟等[15]方法计算棉苗的冷胁迫指数(Chilling injury index,CI)。
1.3 数据处理
采用Microsoft Excel进行数据分析,采用GraphPad Prism进行绘图。
2 结果与分析
2.1 各处理对冷胁迫下棉花种子萌发活性影响
研究表明,与对照比较,拌种灵和萎锈灵包衣处理的发芽势分别增加5.33%和9.67%,且差异显著;发芽率分别增加5.14%和10.29%,且差异显著;发芽指数分别增加6.00%和12.00%,且差异显著;活力指数分别增加6.67%和13.33%,且差异显著。拌种灵和萎锈灵包衣的棉种萌发活性指标均增加,冷胁迫条件下,拌种灵和萎锈灵包衣后均能提高种子活性,从而提高棉种发芽率。图1
2.2 各处理对冷胁迫下棉种干物质转移的影响
研究表明,与对照比较,拌种灵和萎锈灵包衣处理的鲜重分别增加3.33%和4.33%,且差异不显著;干重分别增加4.00%和8.00%,仅萎锈灵包衣处理差异显著;干物质转移率分别增加4.00%和9.00%,仅萎锈灵包衣处理差异显著;呼吸消耗量分别增加-6.25%和-7.50%,且差异不显著。拌种灵和萎锈灵包衣的棉种干物质转移指标均增加,萎锈灵包衣处理的增加较大,冷胁迫条件下,两药剂包衣处理均能增强种子活性,提高种子干物质转化效率,增加棉花幼苗干物质积累,更有利于幼苗生长发育和适应冷胁迫环境。图2
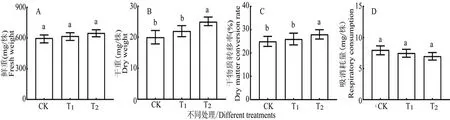
图2 冷胁迫下各包衣处理棉种干物质转移相关指标的比较
2.3 各处理对冷胁迫下棉花幼苗细胞膜系统的影响
研究表明,与对照比较,拌种灵和萎锈灵包衣处理的REC分别增加-3.75%和-12.5%,仅萎锈灵的差异显著;MDA含量分别增加-3.33%和-11.11%,仅萎锈灵包衣处理差异显著。2处理REC和MDA含量均有下降,萎锈灵包衣处理的下降较大,萎锈灵处理较拌种灵处理更有助于避免冷胁迫对棉花幼苗造成的生物膜损伤,对棉花幼苗细胞膜起到保护作用。图3
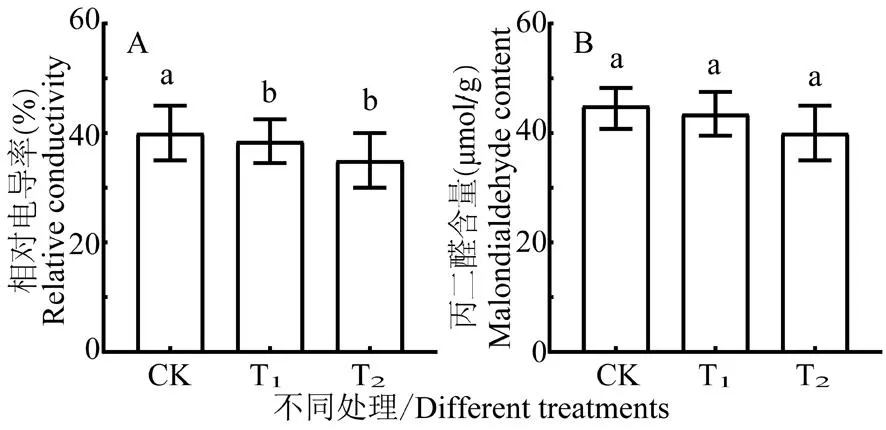
图3 冷胁迫下各包衣处理棉花幼苗 细胞膜系统指标的比较
2.4 各处理对冷胁迫下棉花幼苗渗透调节系统的影响
研究表明,与对照比较,拌种灵和萎锈灵包衣处理的SP含量分别增加6.00%和11.00%,且差异显著;SS含量分别增加3.33%和4.33%,且差异不显著。两药剂处理的SP含量均有所提高,但萎锈灵包衣处理提高幅度较大,萎锈灵更能诱导可溶性蛋白的积累,增强其细胞冷胁迫下的渗透调节能力。图4
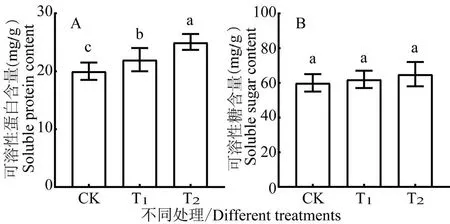
图4 冷胁迫下各包衣处理棉花幼苗 渗透调节物质含量的比较
2.5 各处理对冷胁迫下棉花幼苗抗氧化酶系统的影响
研究表明,与对照比较,拌种灵和萎锈灵包衣处理的SOD活性分别增加6.67%和10.67%,且差异显著;POD和CAT活性分别增加1.50%、1.60%、-3.33%和3.33%,且差异不显著。两药剂处理均能不同程度提高SOD酶活性,萎锈灵包衣处理提高幅度较大,更能提高棉花幼苗叶片抗氧化酶活性,以清除植物体内活性氧的积累,消除活性氧对细胞膜系统的伤害,增强对细胞的保护作用。图5(A-C)
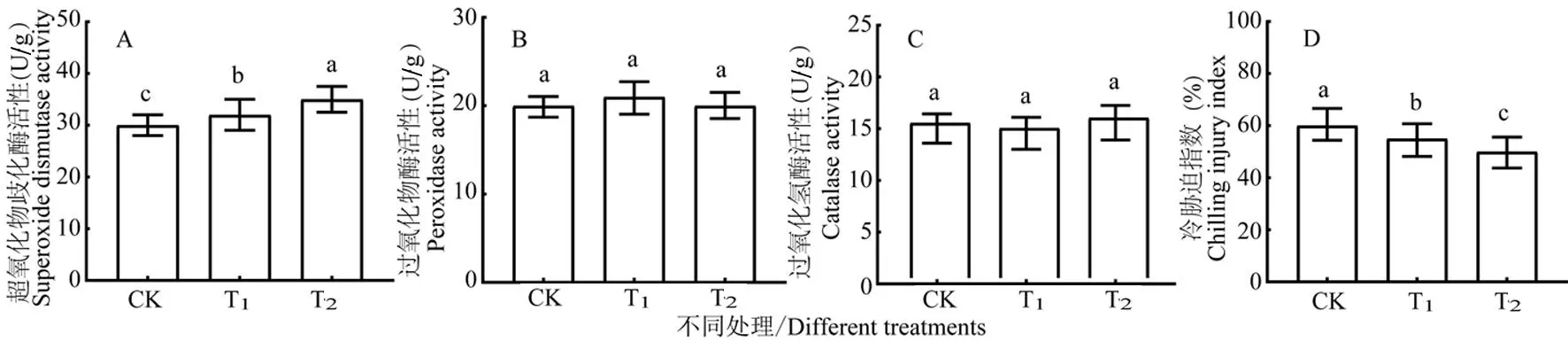
图5 冷胁迫下各包衣处理棉花幼苗抗氧化酶活性和CI的比较
2.6 各处理对冷胁迫下棉苗CI的影响
研究表明,与对照比较,拌种灵种衣剂和萎锈灵种衣剂包衣处理CI分别降低-5.33%和-10.67%,且差异显著。萎锈灵种衣剂包衣处理的CI降幅较大,该处理增强棉花幼苗适应冷胁迫的能力,受冷胁迫程度较轻。图5(D)
2.7 各处理的冷胁迫下棉花幼苗CI与其生理生化指标的相关性
研究表明,拌种灵和萎锈灵包衣处理的棉花幼苗CI与其REC、SP和SOD均显著正相关(P<0.05),相关系数分别为0.91、0.75、0.91和0.89、0.71、0.90;基础种衣剂(对照)包衣处理的棉花幼苗CI与其指标均无显著相关。两药剂包衣处理的棉苗CI与REC、SP、SOD等指标有一定相关性,两药剂处理包衣处理能增强棉花幼苗适应冷胁迫的能力。表1
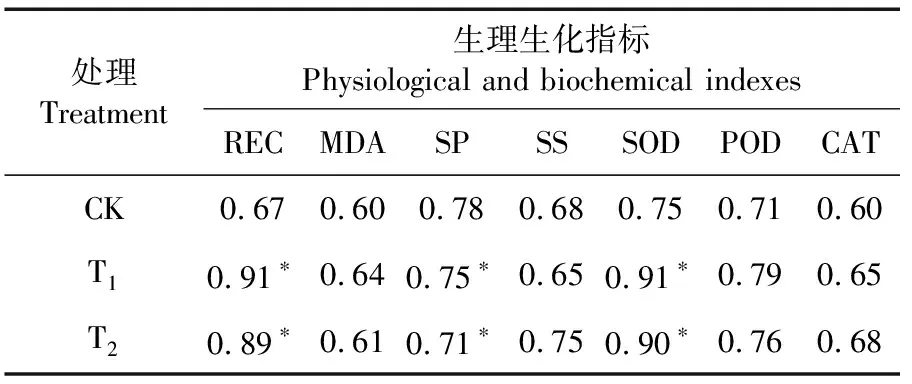
表1 各处理冷胁迫下棉花幼苗CI与 其生理生化指标的相关性
2.8 各处理的冷胁迫下棉花幼苗相关指标的主成分
研究表明,系统默认方差大于1的为主成分,得到第一、第二和第三主成分,其综合评值为F=4.18F1+3.45F2+1.03F3。根据第一、第二和第三主成分与客观权重之积,得到各处理的冷胁迫下棉花幼苗生长的综合影响评判结果。CK、T1和T2处理对棉花幼苗生长影响的综合评价分别为-1.69、2.12和3.69,T1和T2得分均大于1,并且其综合评分分别排在第2位和第1位。萎锈灵包衣处理提高棉花幼苗生长能力的综合影响最佳,拌种灵次之。表2
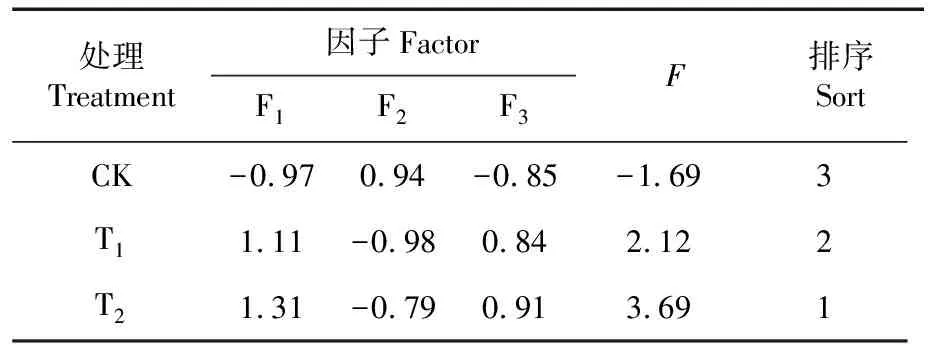
表2 各处理冷胁迫下棉花生长的综合评价
3 讨 论
3.1棉花苗期冷害是制约棉花生产及诱导病害发生的主要因素之一,在萌发过程中种子淀粉酶等酶活性被抑制,细胞内分解转化为能量供细胞生长的物质减少,种子活力减弱,种子萌发延迟,从而抑制种子幼芽和幼根的伸长和生长,造成出苗缓慢,苗瘦苗弱,棉苗无法自然出土;棉花幼苗活性氧代谢失调,自由基积累,细胞膜脂过氧化作用加强,细胞膜结构的完整性遭到破坏、功能丧失、透性增加,而致使细胞液外渗,细胞膜结构的破坏导致叶片坏死,叶绿素含量降低和光合作用效率降低,抗逆性减弱,缺苗断垄甚至毁种重播,最后表现为棉苗植株矮小,生长发育缓慢,对棉花产量品质形成影响非常大。植物耐冷性与植株形态、细胞结构、生理生化以及代谢等方面的抗逆指标相关[16]。大多通过外源植物生长调节物质诱导其耐冷性,如用油菜素内脂处理或脱落酸来提高植物对低温环境下的耐受能力[17,18],也有通过种子包衣技术提高棉花、玉米等在冷胁迫下种子发芽及幼苗的生长能力[19-21]。但是采用杀菌剂提高作物抗冷能力研究报道很少。研究通过琥珀酸脱氢酶抑制剂类杀菌剂制成种衣剂包衣处理,主成分分析表明,萎锈灵处理对增强棉苗耐冷能力的效果优于拌种灵处理。尚需在此理论基础上,进一步通过田间试验考察萎锈灵和拌种灵包衣提高棉苗实际适应冷胁迫的能力。
3.2在逆境环境中,植物体通过增加保护酶活性及时清除活性氧以及调控相关代谢渗透调节剂物质来降低或缓解逆境胁迫带来的伤害[22]。研究表明,2种药剂包衣处理棉苗在处理时间内的保护酶活性存在显著差异,与对照比较,两药剂包衣处理棉花幼苗在冷胁迫下的过氧化物歧化酶活性呈现增高的趋势,丙二醛及可溶性蛋白含量呈现上升的趋势,棉苗通过调节过氧化物歧化酶活性和丙二醛含量、可溶性蛋白含量来降低活性氧伤害,维持细胞内渗透平衡,以避免或降低逆境胁迫对棉苗细胞和组织的伤害,使棉苗耐冷性能力增强,提高适应冷胁迫的调节能力。该结果与前人的抗冷指标变化结论一致[23]。另外,研究结果还表明,棉苗冷胁迫指数与相对电导率、可溶性蛋白和过氧化物歧化酶活性等相关生理生化指标有一定相关性,两药剂包衣处理能增强棉花幼苗适应冷胁迫的能力。有关外源物质包衣处理增强棉花耐冷的能力,是一个复杂的的调控过程,还有待进一步在细胞、分子水平上进行深入研究。
4 结 论
7.2%萎锈灵和40%拌种灵包衣处理均对棉花种子萌发和幼苗的生理特征有一定影响,能促进萌发期种子内营养物质的转化与合成,提高种子活性,从而提高种子的发芽率,通过诱导棉花幼苗细胞内部酶的活性增强,清除幼苗体内活性氧自由基,加快MDA含量的积累,保护幼苗细胞膜结构稳定性,从而提高棉苗适应冷胁迫的能力。两种药剂中,萎锈灵种衣剂诱导棉苗耐冷效果优于拌种灵。
