大洋硅藻席沉积与冰期旋回中的碳-氮循环
2024-02-22蔡雯沁黄恩清柳双权田军
蔡雯沁,黄恩清,柳双权,田军
1. 同济大学海洋地质国家重点实验室,上海 200092
2. 中国科学院深海科学与工程研究所,三亚 572000
“中生代海洋浮游生物革命”之后,海洋浮游真核藻类出现三大分支,即颗石藻、沟鞭藻和硅藻,它们共同构成了现代大洋颗粒有机碳泵以及大洋输出生产力的主体[1]。其中硅藻辐射分异的最晚,在新生代始新世、渐新世之交,即东南极冰盖扩张之后,化石记录中的硅藻多样性程度才出现第一次的显著增加[2]。但硅藻演化相当迅速,目前已经分异出数百个属和大约十万个种[3]。硅藻发展出一系列特殊的本领。首先,生活在开放大洋环境的硅藻,发育占了细胞40%体积以上的大液泡,这个液泡可以储存高浓度的硝酸盐和磷酸盐[4]。因此,在营养盐存在频繁脉冲式供应的地区,例如在近岸上升流或者高纬紊流发育的海区,硅藻具有明显的生存竞争优势。一方面,当营养盐供应突然增加时,硅藻可以高效利用、储存并快速勃发;另一方面,当营养盐供应突然消失时,液泡储存的营养盐让硅藻在短时间内可以不依赖于外界的供应[2]。其次,由于通过基因水平交换获得了其他真核生物体的缘故(即二次内共生),硅藻存在类似动物的“尿素循环”[5]。这种功能可以加快细胞内碳和氮的周转和运移,促进新陈代谢过程所需的含氮化合物的合成(例如多氨和谷氨酰胺)。因此,当水体环境出现营养盐供应时,“尿素循环”可以帮助硅藻迅速增殖,从长期氮限制环境中快速恢复和繁盛[5]。
正是硅藻在细胞尺度上的特殊本领,使得硅藻在全球规模的碳、氮循环中扮演重要角色。据估算,现今硅藻贡献了整个大洋约40%的净生产力和高达50%的有机碳输出[6]。每年全球硅藻固定二氧化碳的规模,与整个热带雨林相当[3]。硅藻也是著名的大洋“铁肥实验”(Iron Fertilization)的研究焦点。在南大洋“高营养盐低叶绿素海区”撒入溶解二价铁之后,会出现小型硅藻的勃发,并且大约一半的勃发生物量能够沉降到水深超过1 000 m 的海水中,由此可以推测硅藻勃发在“碳封存”中的重要角色[7]。
在地质时间尺度上,颗石藻和沟鞭藻在白垩纪—古近纪繁盛,而硅藻在渐新世之后崛起。推测在“暖室期”气候条件下,高纬异常温暖,经向能量差异小,全球风场和洋流较弱,相对静水环境有利于颗石藻和沟鞭藻生长。而渐新世之后极地冰盖体积快速增大,高纬显著降温,高纬海区风场、紊流的发育有利于硅藻的繁盛[2,8]。但硅藻并非环境变化的被动产物,硅藻崛起很可能深刻塑造了地球的表层环境。有学者推测硅藻繁盛导致海洋生物泵效率提高,造成大气二氧化碳浓度的持续降低,推动地球气候从“暖室期”进入“冰室期”[3]。这一推测目前还未获得地质记录的佐证,但是硅藻与气候系统的相互作用已经有了明确证据。晚新生代硅藻多样性程度的几次显著增加,与始新世、渐新世之交的C3草原扩张,中中新世C4草类出现以及晚中新世C4草原扩张,在时间上都存在着良好的对应关系[2]。推测草类植被可以加速陆地硅酸盐风化,而入海溶解硅通量的增加促进了硅藻的分异和繁盛。反过来,海洋硅藻勃发加剧了碳埋藏以及气候变冷趋势,而气候干旱化和季节性的增强又有利于草原植被替代森林植被[2,9-10]。因此,气候、硅藻、陆地草原生态系统的协同演化是晚新生代地球表层圈层相互作用的一个精彩例子。
除了在动荡、富营养盐水体中快速勃发的小型硅藻,还有一个重要类群是在层化水体或者洋流锋面交界处勃发的大型硅藻。这种大型硅藻生长周期长达数月,死亡之后可以快速通过水柱沉降在海底,形成限制底栖生物活动的席状或网状沉积物,称为硅藻席沉积(Diatom-mat Deposits)。半个世纪以来这种硅藻席沉积物在全球大洋都有发现。近年来随着我国远洋科考航次的增多,多家海洋研究单位在热带-亚热带西太平洋水深超过5 000 m 的数十个站位,都发现了末次冰期以及冰消期的硅藻席沉积[11]。中文文献中,对该学术主题也做过介绍[12-13]。据估算,大型硅藻勃发对大洋初级生产力以及有机碳、硅输出的贡献并不亚于小型硅藻勃发[14-15]。但无论现代观测还是地质记录,对大型硅藻勃发储碳作用的研究程度都不充分,这导致许多碳循环模型和生物地球化学模型并不包含硅藻席模块。鉴于此,本文从硅藻席沉积时空分布和勃发机制等角度综述该主题的研究进展,并尝试讨论硅藻席沉积对轨道时间尺度上全球碳-氮循环的潜在影响。
1 硅藻席沉积属种、时空分布及其勃发机制
1.1 硅藻席属种
硅藻席主要由大型硅藻组成(图1),包括海毛藻Thalassiothrixspp.、根管藻Rhizosoleniaspp.、半管藻Hemiaulusspp.、角毛藻Chaetocerosspp.、掌状冠盖藻Stephanopyxis palmeriana、圆筛藻Coscinodiscusspp. 和大筛盘藻Ethmodiscus rex等。其中E. rex是热带深海硅藻软泥的常见属种,根据Kolbe 的记载[16],最早由科林伍德在孟加拉湾发现。1952 年英国“挑战者”号科考船在马里亚纳海沟南部水深10 505 m地方再次获得含这种硅藻碎片的沉积物,Wiseman与Hendey 将其正式命名为Ethmodiscus rex(Rattray)Hendey[17]。

图1 常见成席硅藻属种A:海毛藻属Thalassiothrix spp..[14], B:角毛藻属Chaetoceros spp.[18], C:根管藻属Rhizosolenia spp.(拍摄者Lars Edler, http://nordicmicroalgae.org),D:半管藻属Hemiaulus spp.[19], E:圆筛藻属Coscinodiscus spp.[20], F:大筛盘藻Ethmodiscus rex(本次研究,样品来自西北太平洋马里亚纳海沟挑战者深渊)。Fig.1 Images of typical mat-forming diatomsA: Thalassiothrix spp.[14], B: Chaetoceros spp.[18], C: Rhizosolenia spp.(photo by Lars Edler, http://nordicmicroalgae.org), D: Hemiaulus spp.[19],E: Coscinodiscus spp.[20], F: Ethmodiscus rex (this study; samples were taken from the Challenger Deep, the Mariana Trench).
不论小型硅藻勃发,还是大型成席硅藻沉降,都容易形成具有纹层构造的沉积物(图2)。这有两方面的原因:首先,硅藻和陆源输入物通量随时间发生交替变化,同时陆源输入的营养盐也会促使硅藻的优势种属发生更迭,这些会导致层状沉积特征的形成。其次,如前所述,成席硅藻大规模快速沉降在海底形成席状或者网状结构,即便在底层水充氧条件良好状况下也会限制底栖生物的掘穴和扰动活动,因此有利于原始层状沉积构造的保存。例如,在加利福利亚湾全新世沉积剖面里,发现Thalassiothrix longissima硅藻壳体相互铰接,形成了硅藻席沉积的纹层构造[21-22]。

图2 大洋硅藻席沉积形成的纹层构造a:加利福尼亚湾(站位JPC56, 27°30′N、112°6′W, 水深818 m), 主要硅藻属种是Thalassiothrix spp.[22];b:赤道西太平洋(站位TS01-B10, 11°11.7′N、141°48.7′E, 水深8 638 m),主要硅藻属种是E.rex(本次研究)。Fig.2 Laminated structure of diatom-mat depositsSamples are from (a) Guaymas Basin, Gulf of California (JPC56, 27.5°N,112.1°W, 818 m water depth). The dominant diatom species was Thalassiothrix spp.[22]; (b) the equatorial Western Pacific (TS01-B10,11°11.7′N, 141°48.7′E, 8638 m water depth). The main diatom species is E.rex (this study).
1.2 硅藻席沉积的时空分布
现代表层沉积物调查表明,硅藻软泥(Diatom Ooze,硅藻壳体占比超过30%,由小型硅藻、大型硅藻或者二者混合物构成)主要分布在高纬海区,其中3/4 的沉积量分布在南纬50°宽约900~2 000 km的纬向环带中;其次分布在北纬40°以北的太平洋阿拉斯加湾、白令海、鄂霍次克海、日本海等海区[23]。硅藻席沉积除了分布于上述高纬海域外,在中低纬以及赤道海域也大量出现(图3)。不同海域硅藻席的主要构成种属也存在差别。
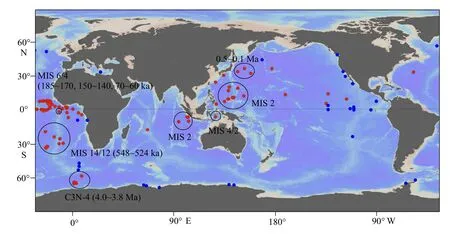
图3 目前发现的全球硅藻席沉积的时空分布其中红色圆点为E. rex 硅藻席, 蓝色圆点为其他种属的硅藻席[14,35-49]。有明确年龄的E. rex 硅藻席沉积用圆圈标注。热带-亚热带西太平洋和热带印度洋E. rex 硅藻席的年龄主要集中在末次冰期[13,17,24-27,50-61], 东太平洋的E. rex 硅藻席沉积大致在上新世和更新世[53,62-64], 大西洋E. rex 硅藻席沉积发生在中—晚更新世冰期[28-31,65-68], 南极洲附近的E. rex 硅藻席沉积, 根据古地磁年龄判断约为4.0~3.8 Ma[32,69-71]。Fig.3 Spatiotemporal distribution of global diatom-mat deposits discovered so farRed dots denote diatom-mat deposits comprised of E. rex, and blue dots represent those comprised of other diatom species[14,35-48]. The E. rex diatom-mat deposits with reliable chronology constraints are circled. The E. rex blooming events in the tropical-subtropical western Pacific and tropical Indian Ocean occurred during the last glacial period[13,17,24-27,50-61], and the blooming in the eastern Pacific occurred in both Pliocene and Pleistocene[53,62-64], while those in the Atlantic occurred across the Mid-Late Pleistocene ice ages[28-31,65-68]. The E. rex blooming event close to Antarctica occurred during 4.0 ~ 3.8 Ma, constrained by paleomagnetic dating[32,69-71].
由于钻孔调查资料的限制,目前发现的硅藻席沉积主要集中在晚第四纪。其中更新世冰期时全球热带、亚热带海区出现大规模的E. rex硅藻席沉积。目前具有可靠定年数据的沉积区域,包括热带-亚热带西太平洋(氧同位素期次4 和2,MIS 4 和2)[11,24-26]、赤道东印度洋(MIS 2)[27]和热带东大西洋(MIS 6 和4)[28-29]以及30°S 附近的亚热带南大西洋(MIS 14 和12)[30](图3)。地质历史时期寡营养热带海区出现大范围E. rex硅藻席沉积的现象,被称为“E. rex难题”[31],它的成因迄今仍在争论。此外,E.rex硅藻席沉积在南极威德尔海区域也有发现,古地磁方法给出的约束年龄约为4 Ma[32]。
在更老的地层中,硅藻席沉积也有零星报道。例如在更新世北非季风强盛期,地中海出现硅藻席勃发和腐泥层沉积,主要由rhizosolenid diatom(广义根管藻,主要包括Rhizosoleniaspp.、Probosciaspp.和Pseudosoleniaspp.等多个属种)和Hemiaulusspp.组成[33]。15~4.4 Ma 期间,赤道东太平洋区域深海钻探钻孔里也出现数次成规模的硅藻席沉积,主要由Thalassiothrixspp.组成[34]。
1.3 大型硅藻勃发机制
目前通过遥感技术、沉积物捕获器以及现场观测等技术手段,发现大型成席硅藻勃发与沉降常常发生在两种水文环境中,即层化水体或者水团锋面交汇处(图4)。热带-亚热带大洋环流圈(tropicalsubtropical oceanic gyres)占据了全球大洋约60%的面积,区域内水体温度较高,层化较强,次表层营养盐很难通过垂向运移输送到表层水体中。再加上远离大陆,缺乏陆源营养盐供应,绝大部分热带-亚热带环流圈的营养盐含量和生物量较低[72-73]。但是成席硅藻的一系列特殊本领却让它们在这种层化水体中获得了生存竞争优势。首先,部分成席硅藻可以在弱光环境下生长,如Rhizosoleniaspp. 和Stephanopyxisspp.[74],因此可以利用深层叶绿素高值区(Deep Chlorophyll Maxima, DCM,一般在100 m以下水深)中的营养盐。由于它们生活在表层水团的底部,因此也可以利用风暴等强扰动事件打破温跃层障碍上涌的营养盐[14,20,75]。其次,成席硅藻的细胞尺寸较大,可以主动调节自身浮力,使其在深层营养源和透光层之间垂向移动,因此既可以利用次表层海水中营养盐,又可以利用表层海水中的光照条件[76-77]。例如Rhizosoleniaspp.个体可达175 μm,通过相互连接,长度可达30 cm,通过调控浮力下沉到营养跃层摄入营养盐后,再上浮到表层进行光合作用[78]。E. rex也可以下潜至水深200 m 左右次表层海水中[31]。可以在弱光条件下生存或者具有浮力机制的成席硅藻也被称为“树荫种”硅藻(Shade Flora Diatoms)[20]。第三,成席硅藻都具有巨大的液泡,可以储存所摄取的营养盐,能够应付长期营养不足的环境[79]。最后,部分成席硅藻可以与固氮蓝细菌共生,获得氮供应(下文详述)[80]。因此,在相对寡营养、层化的海洋环境中,大型成席硅藻可以占据优势的生态位[14,20,75,81]。

图4 “树荫种”硅藻的生长和沉降机制A:生长于深层叶绿素高值区的典型“树荫种”硅藻, B:锋面汇聚带水团相互作用导致的成席硅藻生长和沉降, C:季节性海洋层化结构破坏造成的硅藻席沉积(改自Kemp and Villareal[15])。Fig.4 Mechanisms for the growth and the deposition of shade flora diatomA: Schematic showing the growth of typical shade flora diatom in the “Deep Chlorophyll Maxima” zone, B: the growth and the deposition of mat-forming diatom in the oceanic frontal convergence zone, C: seasonal breakdown of stratification results in a massive deposition of giant diatom (after Kemp and Villareal)[15].
Kemp 等[14]对季节尺度海洋表层环境变化与硅藻勃发的关系做过详细描述。在加利福尼亚湾和地中海,春季时海水混合增强,水体上涌带来了丰富的营养物,导致小型硅藻迅速生长。春末夏初海洋层化加强,表层营养成分逐渐耗尽,小型硅藻勃发结束并快速沉降,以上过程称为“春季勃发”(Spring Bloom)[14]。从早夏到晚秋,海洋表层水由于温度变暖而密度减小,形成较稳定的层化结构,进入成席硅藻占据优势的季节[75]。秋末冬初时随着上层水体的扰动,成席硅藻大量死亡并造成大规模堆积的“秋季倾泻”现象(Fall Dump)[14-15,75]。
此外,Yoder 等[82]在赤道太平洋水团锋面交汇处也观测到成席硅藻的勃发和沉积事件。赤道太平洋叶绿素浓度最高的水体并不在赤道上升流区域,而是位于与热带不稳定波(tropical instability waves, TIWs)传播路径相吻合的水团锋面交汇处。锋面南侧为水温较低的南赤道流,北侧为水温较高的北赤道逆流。根管藻(Rhizosoleniaspp.)在暖水一侧大量富集,导致水体呈暗绿色,而冷水一侧硅藻数量相对较少,水体为浅色。这种水体颜色分界线可以延伸数百千米。推测大型硅藻可能在锋面冷水侧吸收和存储上升流带来的营养盐物质,然后主动迁移到暖水一侧进行生长和勃发(图4B),导致颗粒有机碳输出在热带不稳定波的活跃期间显著增强[14,82-84]。
如前所述,晚第四纪冰期热带-亚热带大洋出现规模巨大的E. rex勃发(图2),但每个海区的勃发机制存在差异。西太平洋和赤道东印度洋E. rex勃发都与大洋层化水体相关。冰期时亚洲冬季风可以向西太平洋供应风尘硅[11],但印度洋的营养盐来源存在争议。Broecker 等认为通过印度尼西亚贯穿流,太平洋温跃层水团可以向东印度洋输送硅藻席勃发所需的营养物质[27]。De Deckker 等则认为由于冰期季风降雨减少有助于东印度洋表层形成高盐度和高营养盐的环境,从而促进了硅藻席勃发[61]。
热带-亚热带大西洋E. rex席勃发与洋流锋面位置的关系更加密切。在热带大西洋,北大西洋中央水团(North Atlantic Central Water, NACW)与南大西洋中央水团(South Atlantic Central Water, SACW)在16°~25°N 位置、水深100~500 m 的地方相遇,形成了佛得角锋面(Cape Verde Frontal Zone)。该锋面在冰期时向赤道方向移动了10~15 个纬度[85]。由于西非岸外大量硅藻席沉积站位与该锋面位置重合,推测硅藻席很可能在锋面两侧通过位置迁移发生勃发。此外,非洲河流可以向近岸地区输入硅藻勃发所需的硅酸盐[29]。在亚热带南大西洋,E.rex硅藻席勃发一般被认为与阿古拉斯洋流(Agulhas Current)输送的营养盐相关[86]。
2 现代大洋成席硅藻与碳-氮耦合循环
据推测,大型硅藻勃发可能贡献了整个大洋约20%的初级生产力和25%的颗粒有机碳输出[14-15]。虽然有硅藻外壳的保护,硅藻勃发产生的绝大部分有机碳还是发生了降解,并不能进入地质储库中[87]。在全球大洋沉积物调查中也常常发现,高生物硅含量的深海沉积物往往只有均值水平的有机质含量[88]。成席硅藻勃发可能是通过影响大洋“呼吸碳库”的方式来调控碳循环(详见第3 部分)。由于成席硅藻碳循环和硅利用方式已经有文献的详细介绍,本节主要聚焦前人较少关注的氮循环部分。
2.1 现代大洋氮循环与同位素分馏
海洋氮循环可以调控大洋硝酸盐和铵盐的浓度水平,因此是决定海洋初级生产力和生物泵效率的关键要素之一,并与碳循环过程紧密相连。海洋氮循环主要包括固氮作用和反硝化作用,此外还包括生物同化作用、再矿化作用和硝化作用等。氮在海洋中的滞留时间约为3 000 年,以硝酸盐为主,储量约为6 000 亿t[89]。生物固氮作用是大洋氮输入的最重要途径,主要发生在寡营养开阔大洋的表层海水中,依靠自养型的蓝细菌完成,每年固氮总量达到约1.63 亿t[90]。相比于大气氮气的同位素值(定义为0),生物固氮产物的氮同位素相较于大气氮气偏负(其值约-2‰~0)[91],低于大洋硝酸盐的平均氮同位素值(5‰)[92]。
海洋反硝化作用主要发生在赤道东太平洋、热带东南太平洋和阿拉伯海等缺氧海区,将硝酸盐还原为氮气[89]。水柱反硝化过程中氮同位素分馏效应比较显著,分馏幅度达到约25‰,但沉积物中反硝化作用几乎不产生同位素分馏效应[91]。每年从水柱和沉积物中通过反硝化途径离开海洋的氮分别约为0.68 和1.32 亿t[89-90]。
2.2 成席硅藻与固氮蓝细菌共生体
如前所述,部分成席硅藻可以利用次表层海水中的营养盐,另一部分成席硅藻可以通过与蓝细菌共生获得氮营养[81]。目前已经发现可以形成硅藻-固氮生物共生体(Diatom-diazotroph Associations,DDAs)的成席硅藻包括半管藻、根管藻、角毛藻以及掌状冠盖藻等属[93-94]。固氮蓝细菌将大气中的氮气转化为可以被寄主硅藻利用的含氮化合物,帮助硅藻突破寡营养海域氮营养的限制。培养实验发现共生体中蓝细菌约22%的碳元素由宿主硅藻提供,宿主硅藻78%~91%的氮元素来自蓝细菌。硅藻细胞越大,固氮和固碳的速率越高[95]。此外,藻类-蓝细菌共生体的固氮/固碳效率还受到多种因素的影响,在一定范围内,光照、温度与固氮效率呈正相关关系[95]。加入铁元素和风尘也可以加强蓝细菌UCYN-A 对氮气的固定[96]。硅藻与蓝细菌共生体的固氮量十分可观,在热带北大西洋海域,硅藻-蓝细菌共生体(Hemiaules-Richelia)与蓝细菌中的束毛藻Trichodesmiumspp.一起为表层水提供了近25%的氮营养来源[97]。
2.3 硅藻沉积物氮同位素指标与海洋生产力
氮同位素是追踪地质历史时期氮循环过程的最重要工具。但是直接测量海洋沉积物全岩样品获得的氮同位素(δ15Nbulk)是一个混合信号,很多时候并不能有效地追踪氮循环过程,这是因为从海洋上层掉落的颗粒氮,在水柱中发生了降解和再矿化作用。在氧化条件下,富含14N 的氨基酸更易被分解,产生的NH4+被黏土吸附,或替代层状硅酸盐矿物中的钾离子[91,98]。这些过程会导致氮同位素发生分馏。但是在还原条件下,有机质降解的同位素分馏效应很小,可以较好地保存海洋有机质的初始δ15N 组成[98]。此外,沉积物样品中有机氮有多个来源,包括陆源输入和多种海洋浮游藻类的生产力贡献。
在富含硅藻的沉积物,尤其是硅藻席样品中,有机质主要由勃发的硅藻产生,来源相对单一,并且硅藻产生的蛋白石可以作为保护有机质的储层,使得硅藻壳体内嵌合的有机质在沉降和埋藏过程中相对不容易受到成岩作用的影响。因此,富含硅藻沉积物常常是氮同位素测试的良好载体。利用硅藻沉积物样品,可以测试有机氮同位素(δ15Norg)、叶绿素(Chlorophyll)氮同位素(δ15Nchl)、硅藻内嵌(Diatom-bound)有机质氮同位素(δ15Ndb)等指标。根据室内培养实验结果,发现δ15Norg与δ15Ndb变化趋势一致,但二者同位素值存在偏移,δ15Ndb总是小于δ15Norg,偏差幅度在1.0‰~14.0‰之间。综合来看,利用同一种属硅藻测试的δ15Ndb指标,是反演表层水体硝酸盐利用程度以及古生产力变化最为可靠的指标[99]。
利用硅藻沉积物氮同位素已经成功回答了地质历史时期海洋生产力变化的一些重要问题。例如,东地中海海底存在著名的腐泥层沉积,有机碳含量可以达到5%。一般认为这些腐泥层形成于北半球夏季太阳辐射量的高值期和北非季风的强盛期(例如早全新世和末次间冰期),一方面非洲尼罗河泛滥和营养盐输入导致地中海生产力勃发,另一方面淡水注入后打断了地中海表层海水的潜沉,底层海水通风减弱逐渐失去氧气,导致有机质在缺氧条件下得到较好的保存[33]。进一步分析发现,腐泥层沉积中事实上存在大量的成席硅藻Hemiaulusspp.和rhizosolenid 硅藻[100],硅藻勃发是当时海洋生产力的主力。通过测试氮同位素,发现在上层海水层化条件加强的情况下,成席硅藻是通过利用次表层营养盐实现大规模勃发的,并与固氮蓝细菌共生形成广泛的固氮作用[15,101]。这纠正了之前的认识,即非洲季风强盛并没有造成地中海表层水的富营养化[101]。值得一提的是,白垩纪时特提斯洋洋底也沉积了大规模富含有机碳的黑色页岩,同样也是成席硅藻Hemiaulusspp.和rhizosolenid 硅藻勃发造成的[102]。
3 大规模硅藻席沉积与大洋碳-氮循环
3.1 冰期热带硅藻席沉积与全球碳循环
第四纪大气CO2浓度在冰期-间冰期旋回过程中有80~100 p.p.m.v (Parts Per Million by Volume,百万分比浓度)波动,其变化机制是学术界长期探索的课题。一般认为,大气CO2浓度变化主要是通过调节深海碳储量来实现的[103]。由于冰期大洋内部通风速率下降,再矿物作用产生的“呼吸碳”(Respired Carbon),可以更长久地滞留在大洋内部,引起了大洋无机碳含量上升和溶解氧浓度下降[104-105]。冰消期时,在南大洋附近随着深部海水上翻,大洋内部“老碳”释放并再次进入大气圈中[106]。
过去认为冰期深海内部发生降解的颗粒有机碳主要来自表层生产力输出,但晚第四纪冰期热带大洋广泛存在的E. rex成席硅藻勃发事件(图3),说明在大洋寡营养海区的次表层海水中,也存在规模可观的生产力输出事件。据估算,在热带西太平洋,E. rex勃发事件期间的海洋初级生产力与近岸上升流地区水平一致[87]。不过硅藻席沉积物的有机碳含量仅为0.2%~0.3%[87],说明硅藻勃发产生的绝大部分有机质在水柱以及水-沉积物界面处发生降解,成为冰期大洋内部的“呼吸碳”,因此冰期E.rex勃发充当了从大气向深海内部运输碳的重要角色。但成席硅藻勃发在碳循环中的角色长久被忽略,未来需要进一步调查,厘定冰期E. rex勃发事件规模及其对大洋碳存储的影响。
此外,在冰期-间冰期时间尺度上,海洋固氮作用与反硝化作用的比例波动调控了海洋氮储库总量和大洋初级生产力,它们也可以影响到大气CO2浓度[2,89,107]。一般认为,冰期时大陆架暴露、赤道上升流增强等因素,可以增加磷元素供应,刺激固氮作用[107-109]。同时冰期水体混合增强,溶解氧含量增加,总体上会削弱反硝化作用[110]。因此,冰期大洋氮储库总量水平上升。模拟实验也表明,相比于全新世,末次冰盛期时全球海洋氮储库最多可能增加了一倍,即从6 000 亿t 提高到1.2 万亿t[111]。同样地,这些研究忽略了冰期E. rex勃发事件对大洋固氮作用的影响。目前对冰期E. rex勃发机制和营养盐来源还存在争论。在热带西太平洋,硅同位素示踪研究结果表明,E. rex勃发利用的是亚洲内陆的风尘硅输入[11]。根据现代生态调查,E. rex并不能与固氮蓝细菌共生,很可能是通过浮力机制利用了次表层水体中的营养盐,包括硝酸盐和磷酸盐等,因此获得了生存竞争优势并触发了勃发事件。冰期热带大洋E. rex大规模勃发提高了上层海洋营养盐的周转速率和利用水平,因此可能刺激广大寡营养海区固氮微生物的生产力,从而提升大洋的固氮作用。
3.2 硅藻席沉积与大洋碳同位素重值现象
新生代以来,大洋无机碳碳同位素(δ13C)最显著的轨道周期就是40 万年长偏心率周期[112]。最近5 Ma 以来,全球大洋δ13C 波动一共出现了约13 次长偏心率周期,在每个偏心率低值期,都会出现δ13C 偏重现象,被命名为碳同位素重值期(δ13Cmax)。其中,近1.6 Ma 以来,大洋δ13C 波动不再严格跟随天文长偏心率周期,而是拉长成为延续时间达到50 万年的事件[113-114]。最近两次碳同位素重值事件分别发生在氧同位素13 期(δ13Cmax-II, 0.53~0.47 Ma)和3-1 期(δ13Cmax-I, 0.05~0 Ma)(图5)。先前研究已经注意到,这两次碳同位素重值事件都伴随着大洋硅藻席勃发事件。首先,氧同位素14 和12 期时亚热带南大西洋发生E. rex勃发事件[30],对应于δ13Cmax-II事件。其次,末次冰期热带西太平洋-印度洋以及赤道东大西洋E. rex勃发事件[11,24-29],对应于δ13Cmax-I事件(图5)。目前还未查清E. rex勃发与大洋碳同位素重值信号是否存在机制上的关联。
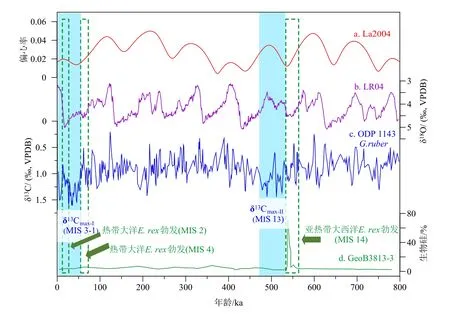
图5 80 万年以来两次大洋碳同位素重值事件与成席硅藻勃发事件的可能关联a:地球偏心率变化来自La2004[115], b:全球底栖有孔虫氧同位素合成曲线来自LR04[116], c:南海ODP 1143 站位浮游有孔虫δ13C 记录(9°22′N、113°17′E, 水深2 772 m)[117], d:亚热带南大西洋GeoB3813-3 站位生物硅含量变化(32°16′S、 21°58′W, 水深4 331 m)[118]。蓝色阴影标示碳同位素重值事件,绿色虚线框标示大洋硅藻席勃发事件。Fig.5 Potential linkage between oceanic δ13Cmax events and diatom blooming events over the past 800 thousand yearsa: The Earth’s orbital eccentricity changes (from La2004) [115]; b: global stack of benthic foraminiferal δ18O, LR04[116]; c: the δ13Cmax events represented by planktonic foraminifera δ13C of ODP Site 1143, southern South China Sea (9°22′N、113°17′E, water depth 2 772 m)[117]; d: biogenic silica concentration(SiO2%) of core GeoB3813-3, South Atlantic (32°16′S、 21°58′W, water depth 4 331 m)[118]. Blue rectangles indicate the duration of the δ13Cmax events and green dotted line boxes show the mat-forming diatom blooming events.
为了解释大洋δ13C 的长偏心率周期,先前研究提出“溶解有机碳假说”[114]。现在全球海洋溶解有机碳库总储量为6 620 亿t,与大气碳储库7 500 亿t接近[119]。并且海洋溶解有机碳库中的95%为惰性溶解有机碳[120],在海洋中滞留长达6 000 年[121]。因此,海洋溶解有机碳库变化可以大幅度影响到大气CO2浓度以及海洋无机碳δ13C 值。根据假说,在偏心率低值期,由于热带季节性和季风减弱,入海营养盐输入和海洋真核藻类生产力减弱,导致大洋生产力以原核类生物为主,海洋溶解有机碳库储量扩大,扣押了大量12C,导致海洋无机碳δ13C 值上升[114]。
由于硅藻席勃发时,大量有机质在水柱中发生了降解,只有极少部分有机质进入埋藏的海洋沉积物中[87]。推测硅藻席有机质降解产物中,除了一部分形成无机形式的“呼吸碳”,还有一部分被微生物转化,形成溶解有机碳。这两种碳的赋存形式对大洋无机碳δ13C 值的作用刚好相反。“呼吸碳”的同位素值较轻,会降低大洋无机碳的δ13C 值;但如果形成溶解有机碳,相当于从其他储库中带走12C,最终使得大洋无机碳δ13C 值升高。因此,硅藻席勃发如何影响偏心率低值期的大洋无机碳δ13C 重值信号,还需要大量记录的约束以及模拟的验证。上的关联。未来更深入的研究需要对地质历史时期的硅藻席勃发和沉积事件进行更广泛的调查,再结合模型来定量约束硅藻席勃发事件在轨道尺度的大洋碳-氮循环中的作用。
4 结论与展望
(1)成席硅藻在层化或锋面环境中勃发可能与其独特的生理结构密切相关。硅藻在海洋浮游藻类演化序列上出现和分异的时间较晚,导致硅藻具备了与蓝细菌共生和尿素循环等特殊功能。成席硅藻更是进一步发展出大液泡,可以通过浮力调控机制利用次表层水体中的营养盐,从而在面积广阔的热带-亚热带层化和寡营养海洋中获得了生存竞争优势,形成大规模勃发事件。
(2)硅藻席勃发和沉积过程可能促进了冰期大气二氧化碳浓度的降低。一方面,成席硅藻席勃发过程中,由于绝大部分有机质在水柱中发生了降解,推测可以贡献冰期深海内部的“呼吸碳库”。另一方面,硅藻席勃发还可以提高海洋上层营养盐的周转速率,很可能导致冰期大洋氮储库容量和表层生产力的扩增。这两个过程都可以促进冰期大气二氧化碳浓度的降低。
(3)成席硅藻勃发事件的气候意义并没有得到充分的重视和理解。在晚第四纪冰期阶段,成席硅藻E. rex在热带-亚热带大洋发生大规模的勃发和埋藏,向大洋深部和海底沉积物输送了大量有机碳和生物硅,但现在的碳循环模型以及海洋生物地球化学模型都缺少硅藻席模块。
(4)低纬大洋E. rex的勃发事件可能与大洋无机碳碳同位素重值事件相关。50 万年以来海洋无机碳碳同位素出现两次重值事件,期间都伴随低纬大洋E. rex的成规模勃发,推测两者之间存在成因
