地菍总黄酮抗2 型糖尿病作用机制的网络药理学研究及实验验证
2024-01-29莫烨云朱盼唐雨菲李笑笑杨秋莉李金燕韦丽秋李丽
莫烨云,朱盼,唐雨菲,李笑笑,杨秋莉,李金燕,韦丽秋,李丽
广西中医药大学 药学院 广西高校中药神经-代谢及免疫药理重点实验室,广西 南宁 530200
糖尿病是目前全球最突出的慢性疾病之一,其病因与遗传、环境、年龄等多种因素有关[1]。相关研究表明,中药能显著改善糖尿病患者的血糖和临床指标,有效延缓糖尿病的进展[2]。地菍为野牡丹科植物地菍Melastoma dodecandrumLour.的全草,广西民族民间主要用来治疗“消渴”“虚劳”“肠炎”等病证,是瑶医药的常用药材之一[3]。在本课题组的前期研究中,地菍不同部位提取物对胰岛素抵抗、链脲佐菌素(STZ)所致高血糖大鼠和小鼠模型均有不同程度的降血糖和调脂作用,其中地菍总黄酮不仅能降低2 型糖尿病大鼠的血糖,改善血脂及氧化应激紊乱,缓解糖尿病病程发展引起的肝肾功能损伤及胰腺病理损伤,且其代谢组学研究结果表明地菍总黄酮发挥抗糖尿病药理活性的主要作用途径可能是苯丙氨酸、酪氨酸和色氨酸的生物合成通路,甘油磷脂代谢通路,花生四烯酸代谢通路,牛磺酸和牛磺酸的代谢通路,胆汁酸的生物合成通路及烟酸和烟酰胺代谢通路[4-17]。但地菍总黄酮治疗2 型糖尿病的物质基础及作用靶点仍不清楚。因此,本研究采用网络药理学的研究方法,对地菍总黄酮的化学成分及其治疗2 型糖尿病的相关靶基因和代谢通路进行较全面系统地分析,探讨地菍总黄酮治疗2 型糖尿病的作用机制;再结合动物实验以高脂饮食诱导和ip STZ 构建2 型糖尿病模型大鼠,记录大鼠体质量、饮水量和摄食量,检测血糖及血脂的相关指标,qPCR 法检测地菍总黄酮对2 型糖尿病大鼠肝组织中磷脂酰肌醇3-激酶(PI3K)、蛋白激酶B(Akt)、腺苷酸活化蛋白激酶(AMPK)作用靶点的调控作用,蛋白质印迹(Western blotting)法检测大鼠肝脏中Akt、AMPK、胆固醇调节元件结合蛋白-1C(SREBP-1C)的蛋白表达量,为后续的机制研究提供一定的理论和实验基础。
1 材料与方法
1.1 动物
SPF 级雄性SD 大鼠,180~220 g,购于湖南斯莱克景达实验动物有限公司,许可证号SCXK(湘)2019-0004;室内环境温度:21~25 ℃,相对湿度:50%~70%,在光照周期12 h/12 h 环境中分笼适应性饲养。动物实验经广西中医药大学实验动物福利伦理委员会批准,批号DW20230528-103。
1.2 材料与试剂
地菍购于南宁小瑶王药店,由广西中医药大学药学院药用植物教研室梁子宁教授鉴定为地菍Melastoma dodecandrumLour.的干燥全草。
链脲佐菌素(质量分数≥75%,美国Sigma 公司,批号20200320);盐酸二甲双胍片(中美上海施贵宝制药有限公司,规格 0.85 g/片,批号ABQ2906);总RNA 提取试剂盒(上海Promega Corporation 公司;批号LS1040);反转录试剂盒(赛默飞世尔科技公司,批号K1622);无水乙醇(广州化学试剂厂);引物(上海捷瑞股份有限公司);MaximaTMSYBRGreen/ROXqPCRMasterMix(赛默飞世尔科技公司,K0221);Akt 兔抗(Cell Signaling Technology,货号#4685);AMPK兔抗(Cell Signaling Technology,货号#2603);SREBP-1C 兔抗[亚飞(上海)生物医药科技有限公司,货号AF6283];GAPDH鼠抗(上海康成科技发展有限公司,货号KC-5G4);PVDF 膜(美国密理博,货号IPVH00010);Marker(赛默飞世尔科技公司,货号PM2510);蛋白定量分析试剂盒(上海博彩医疗器械有限公司,货号K3000);HRP 羊抗鼠(武汉博士德生物工程有限公司,货号BA1050);HRP 羊抗兔(武汉博士德生物工程有限公司,货号BA1054)[2]。
1.3 主要仪器
匀浆机(Fastprep-24 MP);LX-100 离心机(江苏海门其林贝尔仪器制造有限公司);YXQ-LS-30S立式压力蒸汽灭菌器(上海博迅实业有限公司医疗设备厂);SKG-01 电热恒温干燥箱(湖北省黄石市医疗器械厂);LightCycler96 PCR 仪(Roche 公司);TS-1 摇床(金坛市富华仪器厂);JS-680A 自动凝胶图像分析仪(上海培清科技有限公司);TGL-16 台式高速冷冻离心机(Cence);VE-186 电泳槽(上海天能科技有限公司)。
2 实验方法
2.1 网络药理学预测
2.1.1 地菍总黄酮化学成分的收集与筛选 利用文献报道[3,18-19]检索收集地菍黄酮类的化学成分,在Swiss Target Prediction(http://www.swisstargetpre diction.ch/)平台,根据化合物的吸收、分布、代谢、排泄(ADME)特性进行筛选[3]。
2.1.2 活性成分作用靶点的筛选 利用 Swiss Target Prediction 平台,预测活性成分的潜在靶点(Probability>0),即得药物作用靶点基因。
2.1.3 疾病靶点的收集 在GeneCards 数据库(https://www.genecards.org/)、DrugBank 数据库(https://www.drugbank.ca/)、TTD 数据库(https://TTD.org/)中输入关键词“type 2 diabetes mellitus”进行检索,得到2 型糖尿病相关靶基因。对从GeneCards 数据库获得的靶点按中位数进行筛选,3个数据库去除重复基因,即可得到与2 型糖尿病相关的靶基因。
2.1.4 “成分-疾病-靶点”网络的构建 在Venn 2.1 在线软件作图工具平台(https://bioinfogp.cnb.csic.es/tools/venny/)上分别录入地菍黄酮类成分靶点与2 型糖尿病相关的作用靶点,得到药物与疾病的共同靶点。由Cytoscape 3.7.1 构建活性“成分-疾病-靶点”相互作用网络图。
2.1.5 靶蛋白相互作用网络(PPI)的构建 将作用靶点导入String 数据库(https://string-db.org/),选择multiple proteins,限定物种为homo sapines,置信度>0.4,构建PPI 关系图,并根据蛋白之间的关联度进行排序。
2.1.6 基因功能和通路富集分析 在R 3.6.1 软件安装Bioconductor 软件包“org.Hs.eg.db”并运行,将药物-疾病共同靶点转换成entrezID。然后在R软件安装“clusterProfiler”包,根据已转换的entrezID,以P<0.05,Q<0.05 进行关键靶基因基因本体(GO)与京都基因与基因组百科全书(KEGG)功能富集分析,将结果以条形图形式输出。
2.2 动物实验验证
2.2.1 地菍总黄酮的提取与制备 地菍全草粉碎后称取适量粉末,按照药材质量与溶剂体积为1∶30 加入50%乙醇溶液浸泡过夜,使用真空抽滤泵对药液进行抽滤并收集抽滤液。使用真空旋转蒸发仪对抽滤液进行减压浓缩并收集浓缩液。使用AB-8大孔树脂对浓缩液进行纯化,上样后依次用纯化水和70%乙醇进行洗脱,并收集70%乙醇洗脱液。将洗脱液置于55 ℃恒温水浴锅上干燥,得到棕黄色地菍总黄酮粉末。经测定地菍总黄酮的质量分数为73.9%,RSD 为0.83%。对地菍总黄酮的最大给药量进行安全性测定,结果显示地菍总黄酮ig 的日最大给药量为11.2 mg/g。动物给药前将地菍总黄酮粉末放入研钵中仔细研磨,称取适量药粉加入0.5%羧甲基纤维素钠(CMC-Na)溶液,配制不同浓度的地菍总黄酮混悬液,置于4 ℃冰箱保存备用。
2.2.2 2 型糖尿病大鼠模型建立、分组及给药 将60 只SPF 级SD 雄性大鼠适应性饲养1 周后,根据体质量随机抽取8 只大鼠给予普通维持饲料作为对照组,其余大鼠均以高脂高糖饲料(普通饲料73.5%、蔗糖10%、猪油10%、蛋黄粉5%、胆固醇1%、胆酸钠0.5%)连续喂养6 周,禁食不禁水12 h后,按照40 mg/kg 的剂量ip STZ(临用前使用pH 4.2 柠檬酸-柠檬酸钠缓冲液配成1%的溶液),1 周后尾静脉采血,测定空腹血糖(FBG)≥11.1 mmol/L 即为造模成功[10-11]。
将2 型糖尿病模型造模成功的40 只大鼠随机分成模型组、二甲双胍组(0.2 g/kg)组、地菍总黄酮高剂量(0.34、0.45、0.60 g/kg)组,每组8 只,继续提供高脂饲料喂养。各组每天ig 相应药物,1次/d,对照组和模型组ig 0.5% CMC-Na 溶液,连续给药5 周。
2.2.3 样本收集与处理 给药5 周后,将大鼠用水合氯醛麻醉后腹主动脉取血,处死大鼠,在冰面上分离大鼠肝脏,经冰生理盐水洗净后擦干称质量,-80 ℃冷冻保存[13]。
2.2.4 大鼠体质量、饮水量及摄食量 记录各组大鼠给药前后的饮水量、摄食量变化。每周将各组大鼠禁食不禁水10 h 后检测和记录其空腹体质量值。
2.2.5 糖代谢指标FBG 检测 在给药前、给药第7、14、21、28、35 天,将所有大鼠禁食不禁水12 h 后经尾静脉采血,使用罗式血糖仪测定大鼠FBG。
2.2.6 脂代谢指标检测 按照试剂盒说明书检测各组大鼠血清中低密度脂蛋白胆固醇(LDL-C)、高密度脂蛋白胆固醇(HDL-C)、三酰甘油(TG)、总胆固醇(TC)。
2.2.7 qPCR 法检测大鼠肝脏组织PI3K、Akt、AMPKmRNA 水平 用总RNA 提取试剂盒提取大鼠肝脏总RNA,并测定浓度。按照逆转录试剂盒进行逆转录合成cDNA,用于PCR 反应。根据GenBank 中大鼠PI3K、Akt1、AMPK、β-actin基因的mRNA 序列,用Primer Premier 5.0 软件设计引物,引物序列见表1,由上海捷瑞股份有限公司合成。PCR 反应体系10 μL:SYBR Green Master Mix 5 μL,上、下游引物各0.5 μL,cDNA 模板1 μL,ddH2O 补至10 μL。PCR 反应条件:95 ℃预变性10 min;95 ℃变性15 s,60 ℃延伸1 min,共40 个循环。每个样本重复3 次。

表1 引物序列Table 1 Primer sequence
2.2.8 Western blotting 法检测大鼠肝脏组织Akt、AMPK、SREBP-1C 蛋白表达水平 制备肝组织匀浆,裂解后提取肝组织总蛋白,采用BCA 试剂盒对蛋白进行定量。取适量的蛋白样本变性后进行电泳分离,并转移至PVDF 膜;将PVDF 膜封于5%脱脂奶粉孵育2 h,封闭完成后加入按倍数稀释好的Akt、AMPK、SREBP-1C 一抗4 ℃孵育过夜;孵育过夜后的PVDF 膜洗涤后,加入山羊抗兔二抗孵育继续90 min;孵育完成的PVDF 膜洗涤后,按照ECL 发光试剂盒说明书操作,使用凝胶系统进行观察并采集图像,并用JS-680A 自动凝胶成像分析仪器中SensiAnsys 软件进行目的条带的灰度值分析。
2.2.9 数据处理 应用SPSS 20.0 软件进行统计学分析,结果以表示,采用ANOVA 分析多组间比较,采用t检验分析组间比较,荧光定量PCR结果用2-ΔΔCt法进行计算。
3 网络药理学分析结果
3.1 黄酮类活性成分及其作用靶点
从文献检索出地菍黄酮类化学成分并通过Swiss Target Prediction 数据库进行ADME 筛选,最终筛选出活性成分 12 个,利用 Swiss Target Prediction 数据库进行靶点预测得到776 个基因靶点,删除重复项后得到213 个药物潜在靶点。其中4 个成分未预测到靶点,其余8 个活性成分见表2。

表2 地菍黄酮类活性成分基本信息Table 2 Basic information on the active ingredients of flavonoids in M. dodecandrum
3.2 疾病靶点的收集
基于GeneCards、DrugBank、TTD 平台筛选、去重后,分别获得1 151、116、81 个靶点,合并3个数据库的靶点,删除重复项后共得到1 274 个疾病潜在靶点。
3.3 “成分-疾病-靶点”网络分析
使用Cytoscape 软件构建“成分-疾病-靶点”相互作用网络图,如图1 所示,其中长方形表示药物,三角形表示8 个黄酮类成分,椭圆形表示这些活性成分对应的抗2 型糖尿病作用相关靶点,菱形表示疾病,边代表活性成分与抗2 型糖尿病靶点的相互作用。

图1 “成分-疾病-靶点”网络Fig.1 “Active ingredient -target -disease” network
3.4 靶蛋白PPI 网络分析
在String 数据库中录入上述98 个共同靶点,分析得到PPI 网络,如图2 所示,节点表示靶点,每条边代表蛋白之间的相互作用关系,线条越多代表关联度越大。

图2 地菍黄酮类靶点PPI 网络图Fig.2 PPI network diagram of target of M. dodecandrum flavonoids
PPI 网络中,共有98 个节点,981 条边,平均节点度为20,平均局部聚类系数为0.54。排名前30位的靶点对应的边数如图3 所示,可见蛋白激酶B1(Akt1)、血管内皮生长因子A(VEGFA)、表皮生长因子受体(EGFR)、丝裂原活化蛋白激酶 1(MAPK1)、络氨酸激酶(SRC)、环状素受体1(ESR1)、磷酸肌醇3 激酶调节亚基1(PIK3R1)等靶点位于网络的核心位置,在PPI 网络中发挥关键调控作用。

图3 靶点对应的边数Fig.3 Number of edges corresponding to the target
3.5 GO 注释分析和KEGG 富集分析
将98 个共同靶点经R 语言运行后,GO 分析选取生物学过程、细胞组分及分子功能3 部分,得到交集基因集合共富集至1 712 条生物学过程、53 条细胞组分、121 个与分子功能相关的过程。分别选取前20 条结果形成GO 功能富集的条形图,如图4所示,P代表富集的显著性,颜色越深则显著性越高。交集基因主要参与抗氧化反应(response to oxidative stress)、蛋白激酶B 信号传导(protein kinase B signaling)、脂酰肌醇 3-激酶信号(phosphatidylinositol 3-kinase signaling)、对氧化应激的反应(cellular response to oxidative stress)等生物学过程;主要涉及磷脂酰肌醇3-kinase 复合物(phosphatidylinositol 3-kinase complex)、转移酶复合物,转移含磷基团(transferase complex,transferring phosphorus-containing groups)、膜筏(membrane raft)、膜微结构域(membrane microdomain)、膜区域(membrane region 等细胞组分表达过程;主要富集蛋白质酪氨酸激酶活性(protein tyrosine kinase activity)、蛋白磷酸酶结合(protein phosphatase binding)、磷酸酶结合(phosphatase binding)、跨膜受体蛋白酪氨酸激酶活性(transmembrane receptor protein tyrosine kinase activity)等分子功能。
KEGG 分析发现,交集基因富集在126 条通路上,主要涉及磷脂酰肌醇3 激酶/蛋白激酶B 信号通路(PI3K/Akt signaling pathway)、缺氧诱导因子信号通路(HIF-1 signaling pathway)、晚期糖基化终末产物-糖基化终末产物受体信号通路(AGERAGE signaling pathway in diabetic complications)、酪氨酸激酶-信号转导及转录激活子信号通路(JAK-STAT signaling pathway)、腺苷酸活化蛋白激酶信号通路(AMPK signaling pathway)、肿瘤坏死因子信号通路(TNF signaling pathway)等。选取前20 条目形成KEGG 功能富集的条形图,见图4。
4 实验验证结果
4.1 地菍总黄酮对2 型糖尿病大鼠体质量、饮水量及摄食量的影响
如表3、图5 所示,在给药35 d 后,与对照组相比,模型组大鼠的饮水量及摄食量显著上升,体质量显著下降(P<0.01);与模型组相比,地菍总黄酮0.34、0.60 g/kg 组和二甲双胍组大鼠空腹体质量显著上升(P<0.05)。与模型组相比,地菍总黄酮各剂量组和二甲双胍组大鼠的饮水量、摄食量均呈减少趋势但不显著。

图5 地菍总黄酮对2 型糖尿病大鼠体质量的影响(,n =8)Fig.5 Effect of M. dodecandrum flavonoids on body weight of type 2 diabetic rats (,n =8)
表3 地菍总黄酮对2 型糖尿病大鼠体质量、饮水量、摄食量的影响(,n =8)Table 3 Effects of M. dodecandrum flavonoids on body weight,water intake,and food intake in type 2 diabetic rats (,n =8)
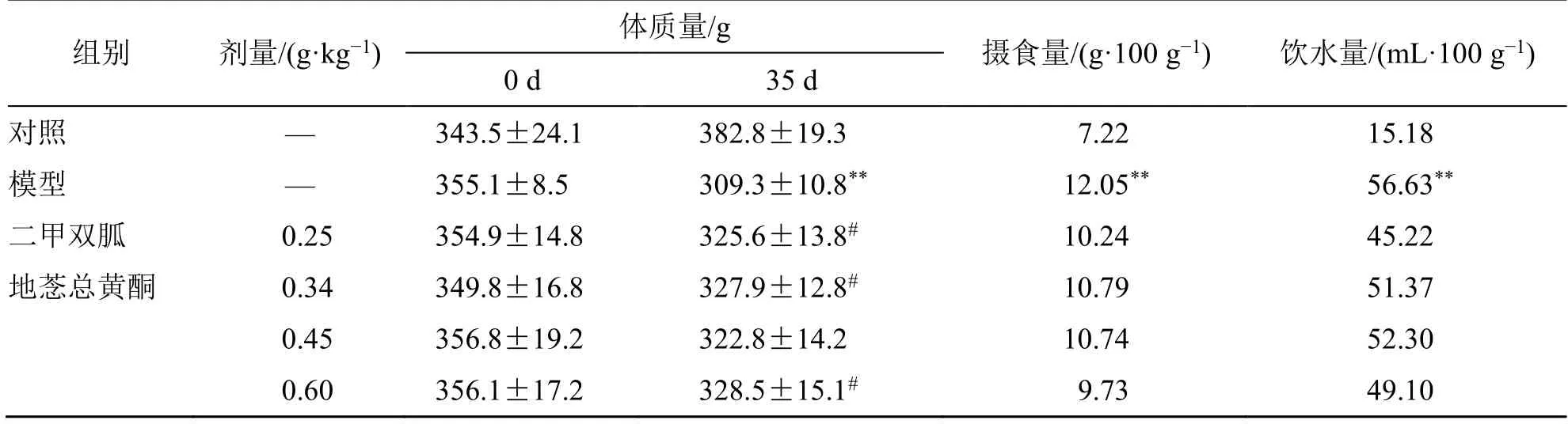
表3 地菍总黄酮对2 型糖尿病大鼠体质量、饮水量、摄食量的影响(,n =8)Table 3 Effects of M. dodecandrum flavonoids on body weight,water intake,and food intake in type 2 diabetic rats (,n =8)
与对照组比较:**P<0.01;与模型组比较:#P<0.05。**P < 0.01 vs control group;#P < 0.05 vs model group.
4.2 地菍总黄酮对2 型糖尿病大鼠空腹血糖的影响
如表4 所示,在给药期间,与对照组相比,模型组大鼠FBG 显著升高(P<0.01),并持续处在高血糖状态;与模型组相比,各给药组大鼠也一直处于高血糖状态,但FBG 呈现逐渐下降趋势;在给药第28、35 天时,除地菍总黄酮0.45 g/kg 组,各给药组大鼠FBG 均显著下降(P<0.05、0.01)。
表4 地菍总黄酮对2 型糖尿病大鼠空腹血糖的影响(,n =8)Table 4 Effect of M. dodecandrum flavonoids on fasting blood glucose in type 2 diabetes rats (,n =8)
与对照组比较:**P<0.01;与模型组比较:#P<0.05 ##P<0.01。**P < 0.01 vs control group;#P < 0.05 ##P < 0.01 vs model group.
4.3 地菍总黄酮对2 型糖尿病大鼠血脂的影响
如表5 所示,与模型组相比,各给药组大鼠血清中TG、TC、LDL-C 水平均下降(P<0.05、0.01),地菍总黄酮0.45 g/kg 组及二甲双胍组HDL-C 水平显著上升(P<0.05)。
表5 地菍总黄酮对2 型糖尿病大鼠血脂的影响(,n =8)Table 5 Effect of M. dodecandrum flavonoids on blood lipids in type 2 diabetes rats (,n =8)
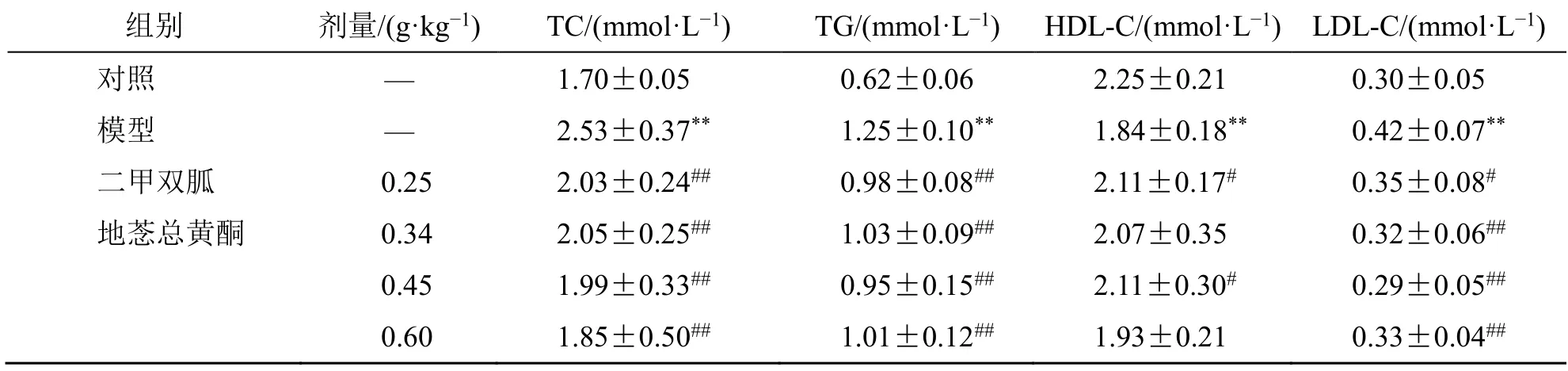
表5 地菍总黄酮对2 型糖尿病大鼠血脂的影响(,n =8)Table 5 Effect of M. dodecandrum flavonoids on blood lipids in type 2 diabetes rats (,n =8)
与对照组比较:**P<0.01;与模型组比较:#P<0.05 ##P<0.01。**P < 0.01 vs control group;#P < 0.05 ##P < 0.01 vs model group.
4.4 地菍总黄酮对2 型糖尿病大鼠肝脏PI3K、Akt、AMPK mRNA 表达的影响
如图6 所示,与模型组相比,各给药组大鼠肝脏中Akt、AMPKmRNA 相对表达量均显著升高(P<0.05、0.01),地菍总黄酮0.60 g/kg 组和二甲双胍组大鼠肝脏PI3KmRNA 相对表达量显著升高(P<0.05)。
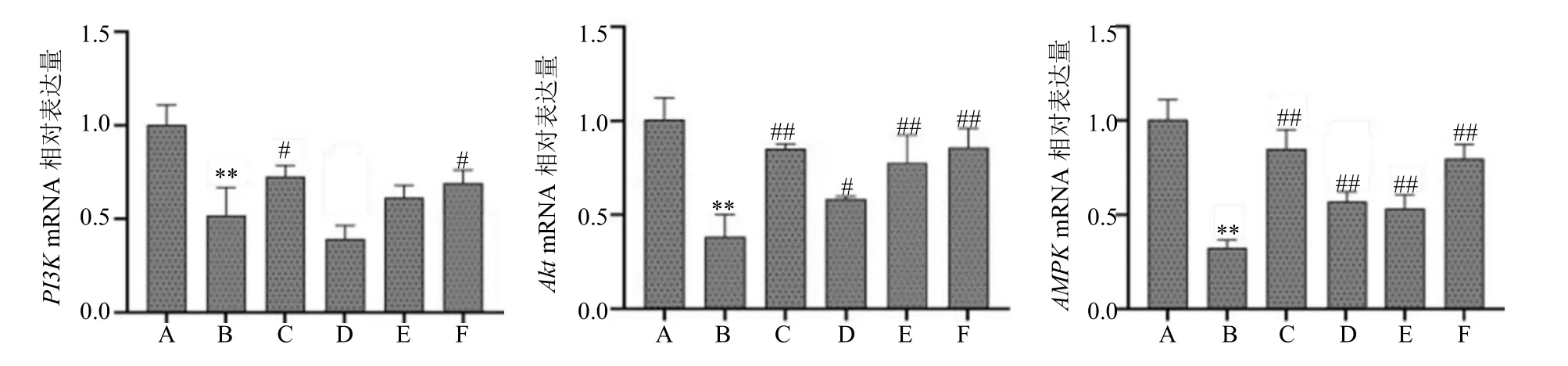
图6 地菍总黄酮对2 型糖尿病大鼠肝脏基因PI3K、Akt、AMPK mRNA 相对表达量的影响(,n =3)Fig.6 Effects of M. dodecandrum flavonoids on the relative mRNA expression of PI3K,Akt,and AMPK in liver of type 2 diabetes rats (,n =3)
4.5 地菍总黄酮对2 型糖尿病大鼠肝脏Akt、AMPK、SREBP-1C 蛋白表达的影响
如图7、表6 所示,与模型组大鼠相比,地菍总黄酮各剂量组大鼠Akt 蛋白表达水平显著升高(P<0.05、0.01);地菍总黄酮0.45、0.60 g/kg组和二甲双胍组大鼠的肝脏AMPK 蛋白表达水平均显著增高(P<0.01),而地菍总黄酮0.34、0.45 g/kg 组大鼠肝脏中SREBP-1C 蛋白表达水平均显著降低(P<0.01)。
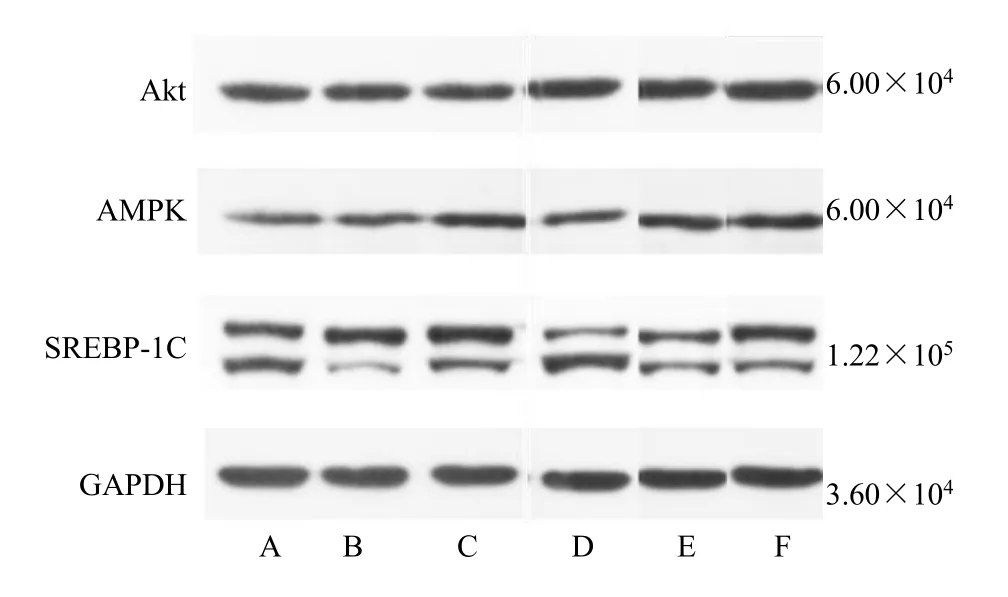
图7 地菍总黄酮对2 型糖尿病大鼠肝脏Akt、AMPK、SREBP-1C 蛋白表达量的影响(,n =3)Fig.7 Effect of M. dodecandrum flavonoids on protein expression of Akt,AMPK,SREBP-1C in liver of type 2 diabetes rats (,n =3)
表6 地菍总黄酮对2 型糖尿病大鼠肝脏Akt、AMPK、SREBP-1C 蛋白表达的影响(,n =3)Table 6 Effect of M. dodecandrum flavonoids on protein expression of Akt,AMPK,and SREBP-1C in liver of type 2 diabetes rats (,n =3)
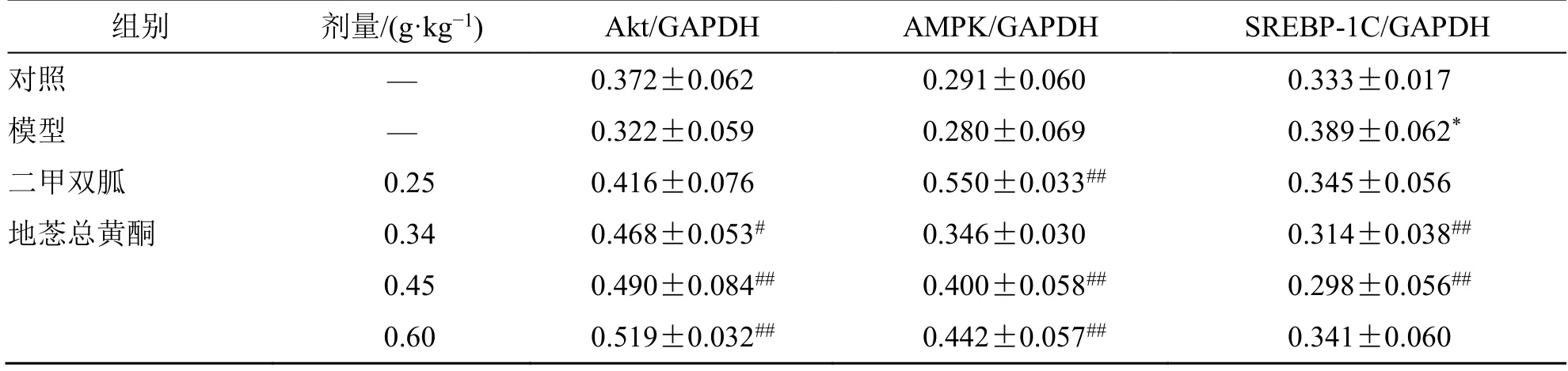
表6 地菍总黄酮对2 型糖尿病大鼠肝脏Akt、AMPK、SREBP-1C 蛋白表达的影响(,n =3)Table 6 Effect of M. dodecandrum flavonoids on protein expression of Akt,AMPK,and SREBP-1C in liver of type 2 diabetes rats (,n =3)
与对照组比较:*P<0.05;与模型组比较:#P<0.05 ##P<0.01。*P < 0.05 vs control group;#P < 0.05 ##P < 0.01 vs model group.
3 讨论
糖尿病是一种多基因疾病,随着时间的推移,单靶向药物的有效性下降,大多数患者需要增加剂量以维持治疗效果,通常与多种不良反应有关[20],因此“一个靶点,一种药物”可能不适用于2 型糖尿病等多基因疾病。多项研究对植物性药物治疗糖尿病进行探索,发现了传统民间药物含有多种具有抗糖尿病活性的活性成分,如黄酮类、三萜类、生物碱和皂甙[21-22]。基于药用植物的天然成分可以针对多种蛋白质,调节多种途径,从而产生协同效应。中草药具有多成分、多靶点、多途径的鲜明特点,可能对糖尿病患者长期血糖的调控发挥重要作用。
本研究通过网络药理学构建“药物-成分-靶点-疾病”的互作网络,筛选出了地菍黄酮类成分抗2 型糖尿病的物质基础可能是槲皮素、芦丁、山柰酚、芹菜素、柚皮素、香叶木素等,相关研究表明槲皮素能有效降低糖尿病大鼠血糖和胆固醇的水平以及调节血脂紊乱[23]。芹菜素能够降低STZ诱导的糖尿病大鼠血糖、血脂水平,增强机体抗氧化能力、降低自由基损伤、改善肝功能[24]。柚皮素具有抑制α-葡萄糖苷酶活性和晚期糖基化终末产物(AGEs)生成的能力,前者能催化水解低聚糖生成葡萄糖,上升餐后血糖,后者过度积累会导致糖尿病并发证的发生发展[25]。因此推测地菍总黄酮中抗2 型糖尿病的主要活性成分可能为槲皮素、芹菜素、柚皮素等。
PPI 网络分析发现地菍总黄酮通过多靶点发挥抗2 型糖尿病的作用,关键靶点可能是Akt1、VEGFA、EGFR、MAPK1、SRC、ESR1、PIK3R1,Akt 属于丝/苏氨酸蛋白激酶家族,是一种调节细胞生长、存活和增殖的多功能蛋白,表达于多种组织中,在2 型糖尿病的发生中起关键作用[26]。Akt 有3 个亚型,即Akt1、Akt2、Akt3,而Akt1 在体内组织中广泛表达[27],许多研究证实Akt1 多态性可能与2 型糖尿病的发生有关[26]。PI3K 是Akt 信号通路中的关键蛋白,可调节细胞存活、生长、分化、葡萄糖运输和利用[28]。PIK3R1 基因编码PI3K 蛋白的调节亚基,可与p110 催化亚基形成PI3K 蛋白[29]。PIK3R1 在胰岛素信号转导中发挥着至关重要的作用[28],其多态性与2 型糖尿病易感性有关[30],沉默PIK3R1 可减轻小鼠的胰岛素抵抗[31]。因此Akt1、PIK3R1 极有可能成为有价值的2 型糖尿病治疗靶点,为2 型糖尿病的治疗提供新的方向。
对靶点进行KEGG 分析表明影响的相关通路可能为PI3K/Akt、AMPK 信号通路,胰岛素信号通路是涉及蛋白质、糖类、脂代谢和生长调控的多条信号通路,其中PI3K/Akt 信号通路是胰岛素信号转导的重要节点,可调节血糖摄取、细胞代谢、细胞存活、增殖、迁移和糖原合成[32-33]。AMPK 通路与PI3K/Akt 通路作为一种中枢能量调节器,在体内调节葡萄糖稳态和脂质代谢中起着重要作用[34]。有研究表明AMPK 激活能调节PI3K/Akt 信号通路,改善肝脏、骨骼肌、脂肪组织糖脂代谢[34]。PI3K/Akt途径也介导脂质合成,SREBP-1C 在肝脏中的过度表达选择性地诱导脂质合成基因的表达。据报道,Akt 可调节mTORC1-S6K1 途径,以调节肝脏中SREBP-1C 的表达[35]。AMPK 也可以通过调节SREBP-1C 的表达来调节脂质代谢,激活脂肪酸、三酰甘油和胆固醇合成相关的基因[36]。本研究动物实验中地菍总黄酮能显著上调2 型糖尿病大鼠肝脏PI3K、Akt、AMPKmRNA 表达,表明地菍总黄酮可能缓解PI3K/Akt、AMPK 胰岛素信号通路的损伤;Western blotting 实验结果显示,Akt、AMPK 蛋白表达量有降低但差异不具显著性,原因可能是mRNA翻译成蛋白的生物学过程具备太多可能性,且它们是通过调控下游产生联级效应,放大通路末游的蛋白SREBP-1C 进而产生调控作用,因此SREBP-1C的蛋白表达水平显著降低,说明地菍总黄酮在一定程度上可能通过影响PI3K/Akt/SREBP-1C、AMPK/SREBP-1C 通路抑制2 型糖尿病大鼠的脂质积累。
综上所述,本研究应用网络药理学方法并结合动物实验,对地菍黄酮类成分与2 型糖尿病之间复杂网络关系进行研究,筛选出了地菍黄酮类成分抗2 型糖尿病的可能物质基础和作用靶点及其抗糖尿病的作用机制。
利益冲突所有作者均声明不存在利益冲突
