小黄花茶群落优势木本植物生态位及种间联结性
2024-01-25李锦婷申开平白小节臧丽鹏何跃军
李锦婷,穆 君,申开平,郭 云,3,白小节,臧丽鹏,李 青,韩 勖,赵 颖,何跃军,*
1 贵州大学林学院,森林生态研究中心,贵阳 550025 2 贵州省赤水桫椤国家级自然保护区管理局,赤水 564700 3 贵州民族大学生态环境工程学院,贵阳 550025
濒危物种是生物多样性的重要组成部分,对生态系统结构及功能维持具有不可替代作用[1]。物种致濒原因包括物种自身特性[2]、生物间的相互作用[3],以及物种与环境因子之间的相互作用等[4]。濒危物种利用资源及适应环境的能力较弱[5],在群落中常受到其它物种的竞争排斥[6],限制其种群更新和维持[7]。然而,濒危物种与其伴生种之间也可能存在互利互补关系,对其生长与存活有促进影响,Soliveres等[8]在研究中发现,伴生植物不仅提高了特定区域中濒危植物的数量,还可能通过防止濒危植物的局部灭绝来促进多物种共存;此外,Holmes等[9]、Van Der Putten等[10]、McIntire和Fajardo[11]在研究中也指出,伴生植物通过缓解濒危植物与其它物种间的竞争强度、降低土壤中的病原体数量以及促进濒危植物的繁殖等方式对濒危植物的生长存在积极作用。因此,研究濒危物种与其伴生种之间的相互作用,探索物种致濒原因及物种共存机制,对珍稀濒危物种的保护具有现实意义[12]。
生态位与种间联结性可从时空尺度上了解物种利用资源和适应环境的能力以及物种间的协作与竞争[13],并能够客观反映现阶段群落结构稳定性[14]。生态位指种群在生态系统中的时空位置及其与相关种群间的功能关系[15],种间联结是不同物种之间的相互作用和空间关系[16]。刘益鹏等[17]认为研究群落中濒危物种的生态位和种间联结性,有助于探索濒危物种在生境中的适应性及与其它物种之间的功能关系。黄祥童等[18]在研究中提到适度限制与濒危物种生态位较高重叠的物种规模,提高与濒危物种有正相互作用的物种比例,有利于促进濒危物种适宜生境营造并达到实效保护的目的。因此,通过对物种生态位和种间联结性特征的研究,能够为珍稀物种或重要模式物种制定种群复壮的保护对策。
小黄花茶(Camellialuteoflora)是山茶科(Theaceae)山茶属(Camellia)小黄花茶组(Sect.Luteotlora)的多年生常绿灌木或小乔木[19],主要模式产地在赤水桫椤国家级自然保护区,于1988年被列为贵州省珍稀濒危保护植物[20]。该物种花型较小、花色金黄、具蜡质光泽,在形态、结构上的特殊性使其具有一定的科研价值和经济价值[21]。最新调查研究发现,赤水桫椤国家级自然保护区境内8个小生境现存小黄花茶种群数量仅为545株,小黄花茶种群数量日渐减少,呈急速衰退的趋势[22]。因此,开展小黄花茶的保护拯救工作刻不容缓。汪豪[23]认为小黄花茶虽然存在一定的生殖障碍,但其遗传多样性保持在非较低水平,说明物种自身特性并非是其致濒的主因。小黄花茶自然更新与其伴生种以及环境之间的相互作用密切相关,探索保护物种种群生态位和种间联结性,有助于清楚认识物种间相互关系并确定群落稳定性,为濒危物种保护提供科学依据[17]。目前,对小黄花茶的研究主要集中在生物学特性[24]、物种起源[25]、种群结构[22]、空间分布格局[26]、繁殖技术等方面[27],而未见对小黄花茶群落物种生态位与种间联结性的研究报道。这些研究难以清楚解释小黄花茶种群衰退的濒危机制。本研究拟通过小黄花茶种群生态位及其与伴生种之间的联结性探索群落稳定性,为小黄花茶物种保护和管理提供理论参考。
1 研究区与研究方法
1.1 研究区概况
小黄花茶集中分布地位于赤水桫椤国家级自然保护区内葫市镇金沙村闷头溪2.14km2范围内,地理位置为:28°28′01″-28°28′56″N,105°57′35″-105°58′47″E,海拔352-756m。该区域属于中亚热带湿润季风气候,河谷具有类似南亚热带的气候特征[26]。年均温17.7℃,年均降水量1200-1300mm,相对湿度大于84%[20]。土壤类型为紫色土,土层深厚肥沃。区域内生物资源丰富,植被类型多样,主要分布中亚热带常绿阔叶林、亚热带常绿落叶阔叶混交林、南亚热带雨林层片的常绿阔叶林等植被类型[28]。本研究调查样地位于赤水桫椤国家级自然保护区葫市镇金沙村闷头溪下河沟、观音岩谷、石缸岩、大湾子、黄泥沟、陈田、血台子、半坡头8个集中分布点,调查样地位置分布如图1。
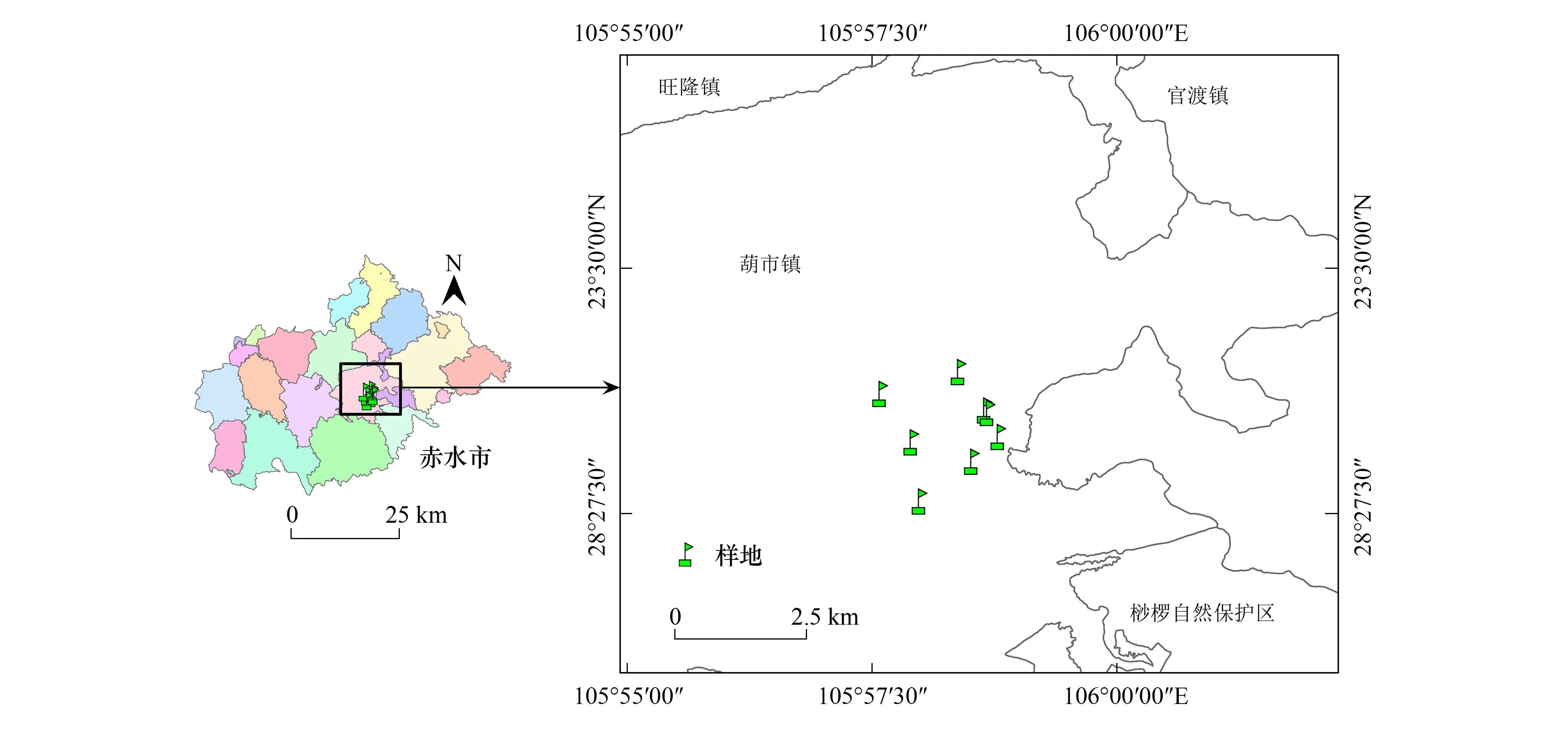
图1 研究区小黄花茶群落调查样地位置Fig.1 Plot location of C. luteoflora community for survey
1.2 样地设置及调查方法
本研究采用典型样地法对以上8个小黄花茶种群集中分布点进行群落学调查。每个调查样地大小20m×20m,在每个样地设置4个10m×10m的相邻格子样方,共计32个样方。调查时记录了所有样方中的小黄花茶种群个体数量及地径、高度和冠幅,以及物种名称、高度、冠幅、株数、乔木胸径及灌木地径等,并记录生境信息如表1。本次调查发现小黄花茶群落乔木层物种主要为毛竹(Phyllostachysedulis)、慈竹(Bambusaemeiensis)、樟(Cinnamomumcamphora)、枫香树(Liquidambarformosana)、赤杨叶(Alniphyllumfortunei)等;灌木层主要物种为小黄花茶、桫椤(Alsophilaspinulosa)、毛桐(Mallotusbarbatus)、鹅掌柴(Heptapleurumheptaphyllum)、盐麸木(Rhuschinensis)等;草本层物种为竹叶草(Oplismenuscompositus)、卷柏(Selaginellatamariscina)、荩草(Arthraxonhispidus)、菝葜(Smilaxchina)和黄精(Polygonatumzanlanscianense)等。
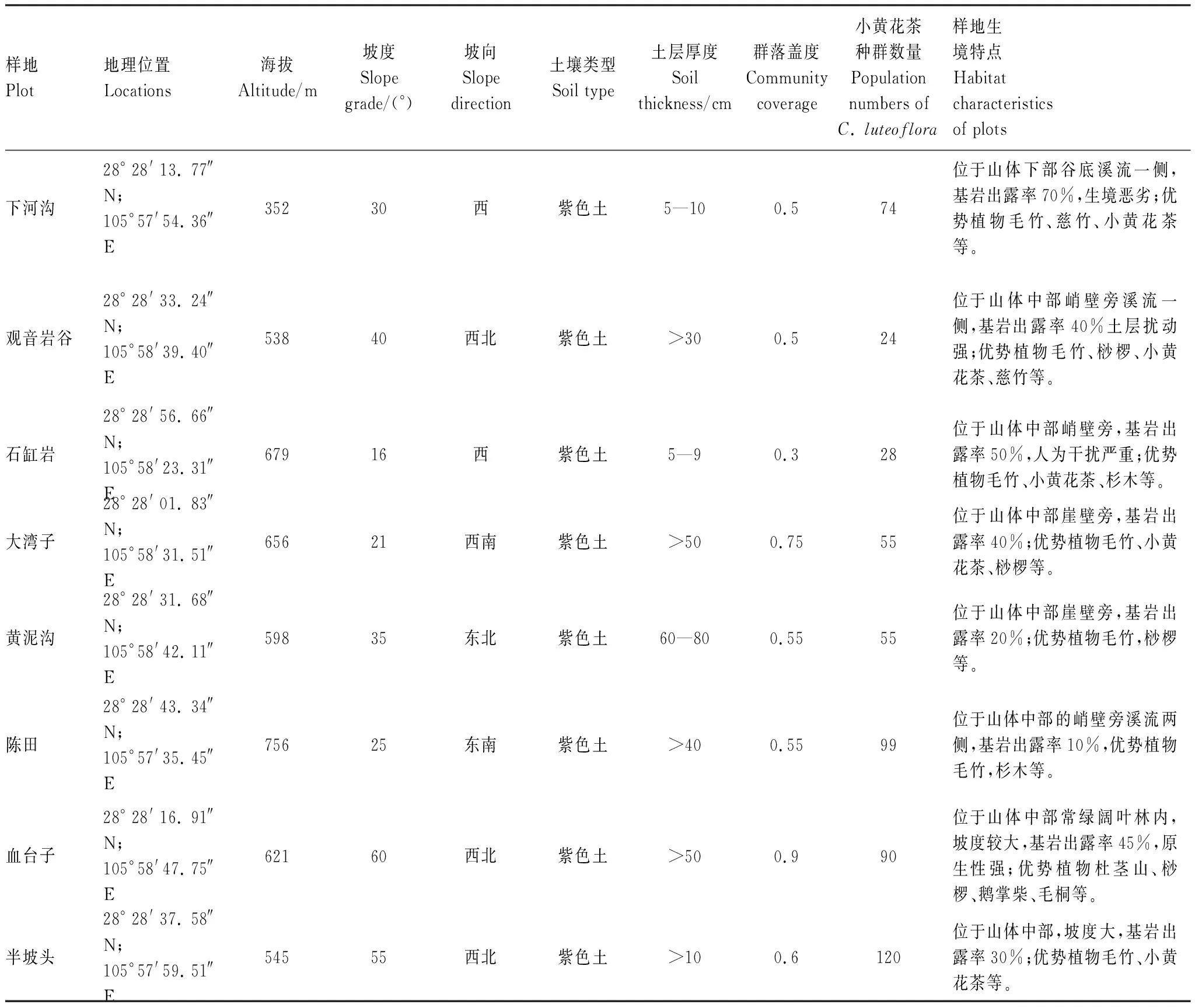
表1 小黄花茶群落调查样地生境特征Table 1 Habitat charateristics of surveied plots on the C. luteoflora community
1.3 数据处理
计算木本层所有物种重要值[29],选择重要值排序前15个优势木本物种进行生态位和种间联结性分析。采用Shannon指数(BS)和Levins指数(BL)计算优势种生态位宽度[30-31],Schoener指数(Cik)和Pianka指数(Oik)计算种间生态位相似性和生态位重叠度[32-33],利用方差比率(VR)和检验统计量(W)分析群落总体联结性[34],选择卡方检验(χ2)、联结系数(AC)和共同出现百分率(PC)联合分析种间联结性[35]。利用R 4.1.3中‘spaa’包进行重要值、生态位与种间联结特征的相关计算及半矩阵图的绘制,Origin 2021绘制AC值、PC值与Cik、Oik之间的线性回归图。各计算公式如下:
(1)Shannon生态位宽度(BS):
式中,BS∈[0,lnr],BS值越大生态位越宽。
(2)Levins生态位宽度(BL):
式中,BL∈[1,r],BL值越大生态位越宽。
(3)生态位相似比(Cik):
式中,Cik∈[0,1],Cik值越大,生态位相似性越高。
(4)生态位重叠值(Oik):
式中,Oik∈[0,1],Oik值越大,生态位重叠程度越高。
(5)方差比率(VR):
式中,VR>1:正关联,VR<1:负关联,VR=1:无关联。
(6)检验统计量(W):
W=VR×r
式中,χ2(0.95,r)
(7)卡方检验(χ2):
式中,ad>bc:正联结,ad
(8)种间联结系数(AC):
式中,AC值越趋近1,正联结性越强,越趋近-1,负联结性越强,AC= 0,种间无联结。
(9)共同出现百分率(PC):
式中,PC值越趋近1,正联结性越强,PC= 0,种间无联结。
公式(1)-(9)中:r为样方总数;Pij=nij/Ni,Pkj=nkj/Nk,nij(nkj)为物种i(k)在样方j的重要值;Ni(Nk)为物种i(k)在全部样方的重要值;Tj为样方j中出现的物种数;t为样方中物种平均数;S为总物种数;ri为物种i出现的样方数;a为物种i和k同时出现的样方数;b为只有物种k出现的样方数;c为只有物种i出现的样方数;d为物种i和k都不出现的样方数。
2 结果与分析
2.1 优势植物重要值与生态位宽度
由表2可知,在群落15个优势物种重要值大小排序中,小黄花茶重要值最大为0.2086,其次为毛竹和慈竹,分别为0.1242和0.1118,这三个物种为群落优势物种,伴生种盐麸木重要值最小为0.0155。15个种生态位宽度Shannon指数BS和Levins指数BL平均值分别为1.36和5.58,群落中小黄花茶生态位宽度BS和BL均最大,分别为2.89和15.76,其次为桫椤,BS和BL分别为2.6728和12.7743。毛竹、毛桐和慈竹生态位宽度值BS和BL均相对较大。枫香树、香椿(Toonasinensis)、杨梅(Morellarubra)和黄葛树(Ficusvirens)的生态位宽度值BS和BL均最小,分别为0和1。结果表明,小黄花茶在群落中具有最大的重要值和生态位宽度值。
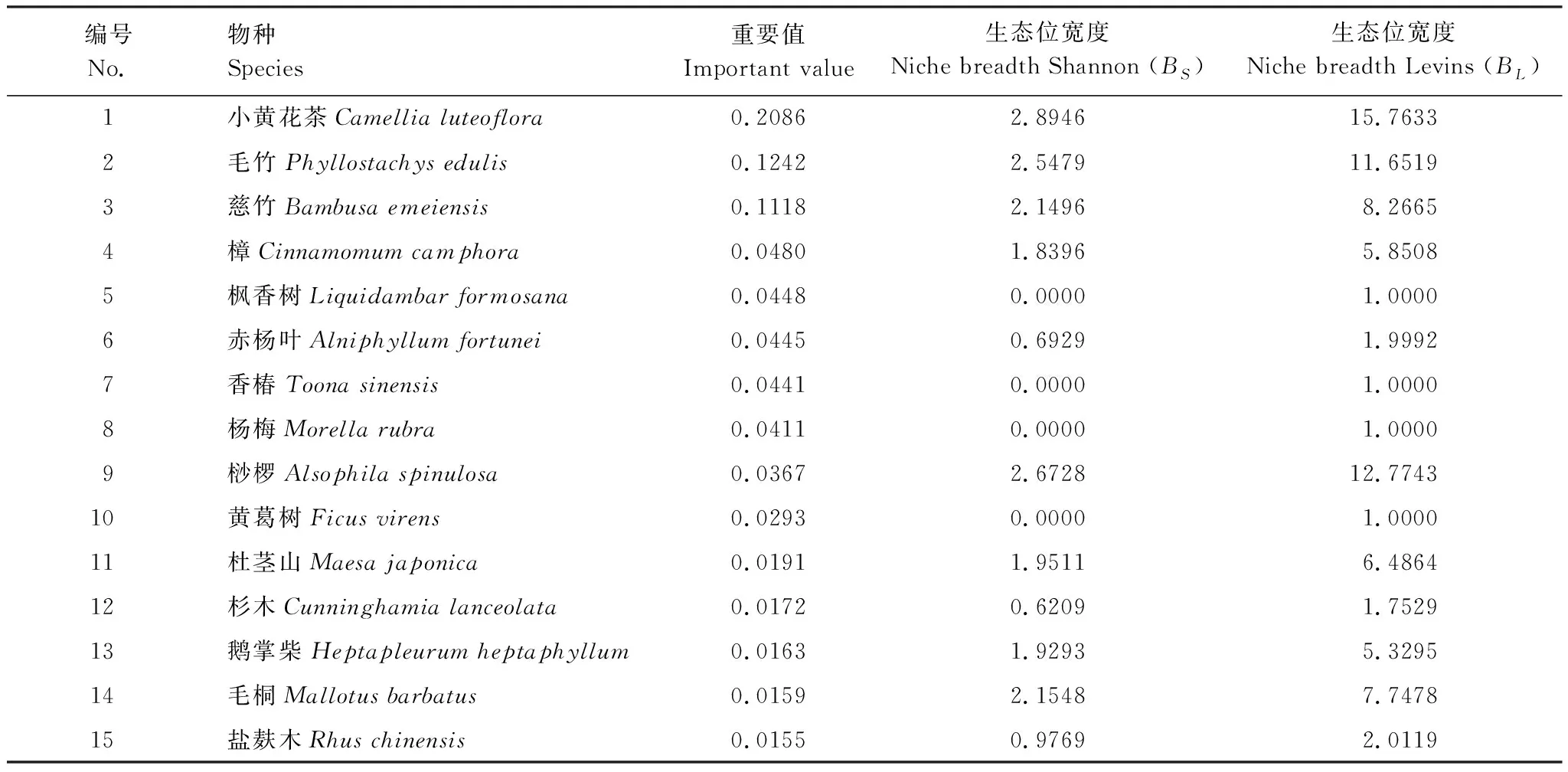
表2 优势植物重要值与生态位宽度Table 2 The important value and niche breadth of dominant plants
2.2 优势植物生态位相似比与生态位重叠值
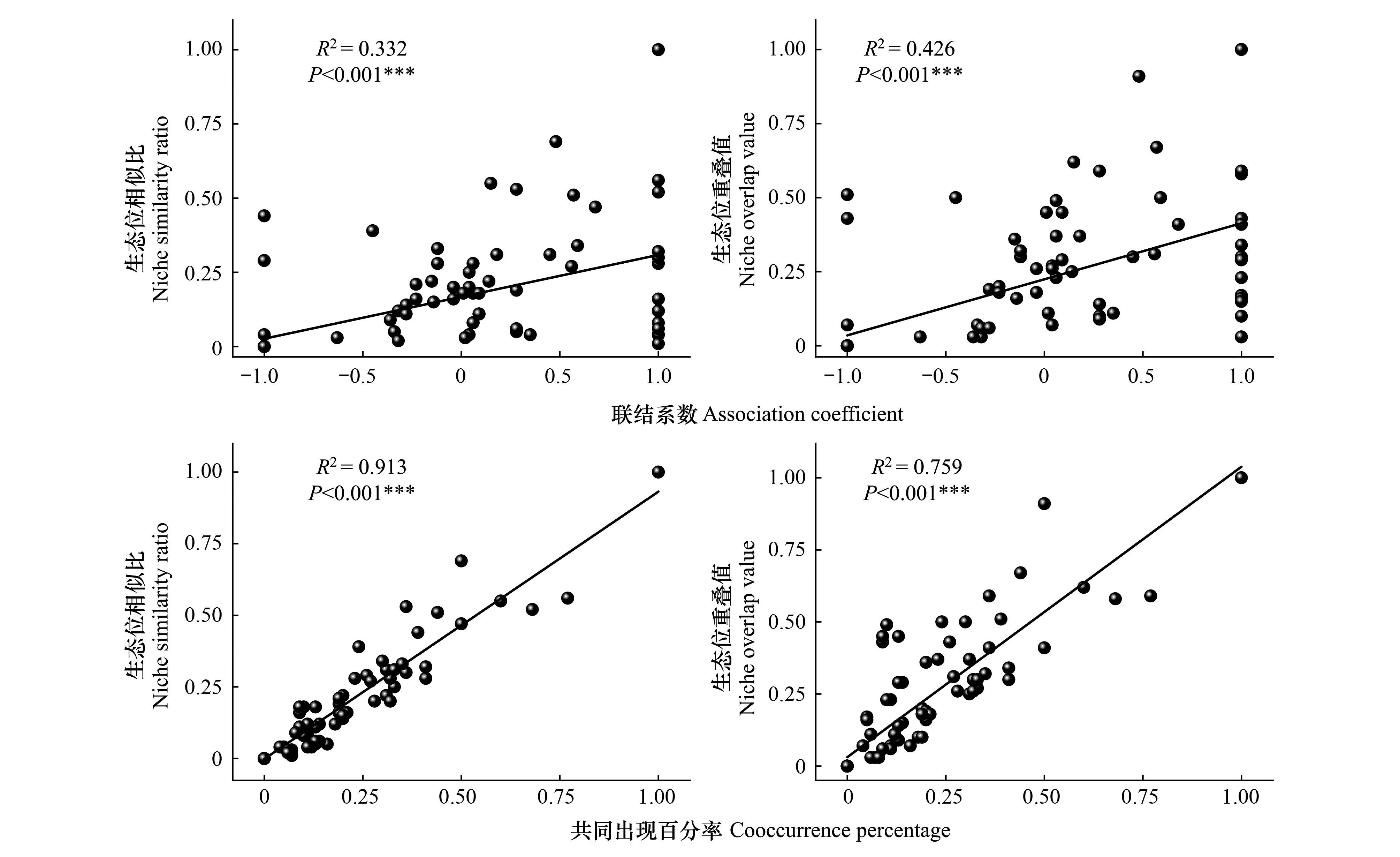
由表3可知,在群落中15种优势植物共组成105个种对,优势植物生态位相似比和重叠值均值分别为0.13和0.17。其中,Cik≥0.50有7个种对,占比6.67%。0 表3 优势植物生态位相似比与重叠值Table 3 The niche similarity ratio and overlap value of dominant plants 对优势植物总体关联性进行分析可知,方差比率(VR)= 0.63,小于1,表明群落优势物种总体关联性为负关联(表4)。卡方临界值χ2(0.95,32)= 20.07,χ2(0.05,32)= 46.19,利用检验统计量W检测VR值偏离的显著性,W= 20.16,介于χ2(0.95,32)与χ2(0.05,32)之间,表明小黄花茶群落15个优势物种总体联结性呈现为不显著负联结。 2.4.1卡方检验 由表5可知,15种优势木本植物组成的105个种对中,有42对呈正联结,占比40%,负联结有63对,占比60%,正负关联比为0.67,显著与不显著之比为0.11。其中,呈显著正联结的有4对,占比3.81%。呈不显著正联结的有38对,占比36.19%。有6对呈显著负联结,占比5.71%。不显著负联结的种对数有57对,占比54.29%。小黄花茶与其它物种组成的14个种对中,有3个种对呈显著正联结(小黄花茶-香椿、小黄花茶-杨梅和小黄花茶-黄葛树),1个种对呈显著负联结(小黄花茶-枫香树),其余10个种对之间联结性均不显著。卡方检验结果表明,优势种之间大多呈不显著负联结;小黄花茶与多数物种联结性较弱。 表5 优势植物种间联结性χ2检验表Table 5 The inspection table of interspecific associations χ2 of dominant plants 2.4.2联结系数AC 由图2可知,105个种对中,正联结种对数为42,负联结种对数为63,占比分别为40%和60%,正负关联比为0.67。其中联结系数AC≥0.67(极显著正联结)的种对数为16,占比15.24%。0.33≤AC<0.67(显著正联结)有5对,占比4.76%。0≤AC<0.33(不显著正联结)有21对,占比20%。-0.33≤AC<0(不显著负联结)有12对,占比11.43%。-0.67≤AC<-0.33(显著负联结)有4对,占比3.81%。AC<-0.67(极显著负联结)有47对,占比44.76%。联结系数AC结果显示,优势物种之间负联结性显著大于正联结性;小黄花茶与其它物种组成的14个种对中,有4个种对呈极显著负关联(小黄花茶-樟,小黄花茶-枫香树,小黄花茶-杉木,小黄花茶-毛桐),其余10个种对呈极显著正关联。 图2 优势植物种间联结性AC值半矩阵图Fig.2 Semi-matrix diagram of AC value of interspecific associations of dominant plantsAC:联结系数Association coefficient;1:小黄花茶;2:毛竹;3:慈竹;4:樟;5:枫香树;6:赤杨叶;7:香椿;8:杨梅;9:桫椤;10:黄葛树;11:杜茎山;12:杉木;13:鹅掌柴;14:毛桐;15:盐麸木;○为极显著正联结;□为显著正联结;△为不显著正联结;●为不显著负联结;■为显著负联结;▲为极显著负联结 2.4.3共同出现百分率PC 为了避免AC值受到d值(两种植物都不出现的样方数)影响造成的偶然误差,选用PC值进一步分析种间联结性。由图3可知,PC≥0.83(极显著正联结)的种对仅有香椿与杨梅,占比0.95%。0.50≤PC<0.83(显著正联结)有5对,占比4.76%,分别为小黄花茶-毛竹、小黄花茶-桫椤、毛竹-桫椤、毛竹-鹅掌柴、枫香树-杉木。0.17≤PC<0.5(不显著正联结)有27对,占比25.71%。PC<0.17(无联结性)有72对,占比68.57%。小黄花茶与其它物种组成的14个种对中,有2个种对呈显著正联结(小黄花茶-毛竹和小黄花茶-桫椤),其余12个种对之间正联结性均较弱。PC结果表明,优势种之间大多呈无关联性;小黄花茶与其它物种间关联性较弱。 图3 优势植物种间联结性PC值半矩阵图Fig.3 Semi-matrix diagram of PC value of interspecific associations of dominant plantsPC:共同出现百分率Cooccurrence percentage;1-15为物种编号同图2;○为极显著正联结;□△为显著正联结;●■为不显著正联结;▲为无联结性 将15个优势物种的AC值(种间联结系数)和PC值(共同出现百分率)分别与相应的生态位相似比和生态位重叠值进行回归分析。由图4可知,线性回归图表明优势物种的AC值、PC值与生态位相似比、生态位重叠值之间存在极显著正相关关系(P<0.001),总体表现为15个优势植物的种间正(负)联结性越强,生态位相似性与重叠程度越高(低)。 图4 优势植物种间联结与生态位相关性回归分析Fig.4 Regression analysis between interspecific associations and niches among dominant plant species Schellenberger Costa等[36]认为重要值和生态位较大的物种具备更强的资源利用能力和环境胁迫适应能力。本研究中,小黄花茶较其他伴生优势植物具有最大的重要值和生态位宽度值(表2),表明小黄花茶在该生境下的群落内部具有较强的竞争适应能力,对维持群落内部环境以及稳定性起重要作用。然而,Serrano等[37]却认为濒危物种通常具有独特的生境需求,生态位较狭窄,这与本研究中小黄花茶具有较宽的生态位不一致,其原因可能由于本研究是基于8个局域生境对小黄花茶进行典型样地调查,从而扩大了该物种在区域尺度上的生态位宽度所致[38]。代玉烜等[39]采用最大熵生态位模型对小黄花茶潜在适应性进行预测研究,发现小黄花茶适应于长江流域黔、川、渝及滇部分区域,意味着该物种具有较宽生态位的潜在适应性。据文献记载,赤水桫椤国家级自然保护区境内(主要分布于闷头溪)小黄花种群数量从2005年1708株下降到2016年800余株[40-42],本此调查发现该生境集中分布的8个局域点留存的小黄花茶种群数量仅为545株,表明小黄花茶种群数量呈急速衰退的趋势。因此,即便小黄花茶具有较宽生态位的潜在适应性[39],但种间竞争挤压,也可能影响小黄花种群数量的维持。此外,曾洪和陈小红[43]认为特定环境下濒危物种能在群落中占据优势地位,但种群结构并不健康,仍然面临着自然更新困难的窘境。本研究前期通过小黄花茶种群数量结构和存活曲线分析发现该种群趋近于Deevey-Ⅰ型,属衰退型种群[22],该区域小黄花茶种群现存幼苗数量严重不足,种群受干扰压力大。由此可见,具有较宽生态位并不代表小黄花茶是生态位泛化种,其可能仍然面临较大的生存压力,种群保育急需开展。再有,生态位较宽的物种之间往往更容易占据相同的资源维度,从而产生较高的生态位相似及生态位重叠现象[44];Costa-Pereira等[45]认为生态位相似和生态位重叠度越高的两个物种,对同等级共享资源的利用方式越相似,在空间配置分布上的竞争潜力越大。本研究中,小黄花茶与毛竹和桫椤之间有较高的生态位相似性和生态位重叠度(表3),表明小黄花茶与毛竹和桫椤之间可能存在较大的资源竞争现象。毛竹生长快、易成林、竞争力强[42],桫椤是赤水桫椤自然保护区主要的旗舰保护物种,而毛竹等竹类在该区域快速扩张,极大程度上抑制了小黄花茶和桫椤在光照和养分的生态位空间,这可能是导致小黄花茶种群数量衰退的重要因素。因此,对毛竹合理经营和管理是保护该生境小黄花茶的重要措施。 本研究联结系数AC结果发现,小黄花茶与其它14个物种形成的种对呈显著正联结的数量多于χ2检验和PC分析数量(表5; 图2,3)。引起这一现象的原因可能是小黄花茶在8个集中分布的局域小生境高频度出现增加了小黄花茶与其它物种出现在同一样方中的概率,从而扩大了小黄花茶与其它物种间的联结显著度,这种现象与叶权平等[46]所提到联结系数AC会扩大种对间的联结显著度一致。此外,本研究还发现小黄花茶在其局域群落与伴生优势物种间关联性较弱,种间联结程度松散(表5; 图2,3)。群落的种间关联受到种间竞争、环境变化、研究尺度等因素的制约[47-48]。赤水桫椤自然保护区地势起伏大,地表形态复杂多样[49],物种长期适应不同微环境,对不同生境资源利用的差异促使物种生态位分化[50]。因此,种间关系趋于相对独立状态,从而导致小黄花茶在该保护区局域生境群落中与其它物种关联性松散。通常情况下,群落的成熟度越高,植物种类组成越趋于完善和稳定,种间关系趋于正联结,从而促进更多物种共存[14]。回归分析发现,小黄花茶群落种间联结性与生态位相似性及重叠度呈极显著正相关性(图4),而小黄花茶群落各优势物种的种间联结分析呈不显著负联结(表5; 图2,3),优势物种间的生态位相似性和重叠程度均较低(表3),表明小黄花茶群落各优势种种间关系松散且群落不稳定。该结果与刘益鹏等[17]研究观光木和李丘霖等[51]研究桫椤等珍稀濒危植物群落生态位和种间联结性的结果一致。 本研究的8个局域生境小黄花茶种群在空间上构成了该保护区小黄花茶复合种群,局域种群分散导致基因交流受阻,增加了种群灭绝风险。此外,因该区域属保护区竹海公园内,毛竹种群快速扩张增强了小黄花茶生态位资源空间压力,竹林经营干扰也进一步加剧其种群衰退趋势。基于本研究结果,小黄花茶种群与其优势伴生种的种间关联性弱,优势物种间呈相对独立分布,群落稳定性低。采取有效措施对小黄花茶进行种质资源保护已势不可缓。可通过排除干扰,加强生境保护,促进小黄花茶种群自然更新。一方面,针对就地保护,可采取适度疏伐同小黄花茶生态位相似性和重叠度高(如毛竹)以及负关联较强的物种(如枫香树),减缓种间竞争压力,使之形成相对稳定的种间关系,提高小黄花茶群落的稳定性;另一方面,针对迁地保护,可选择与小黄花茶生态位重叠度较小且种间正关联较强的物种进行群落配置以促进群落稳定性提升。再有,濒危植物种群衰退的因素是多方面的。本研究仅采用典型样方法对8个局域群落进行调查研究,所获得的结果解释了局域群落内物种之间的相互作用,但一定程度上限制了小黄花茶非集中分布生境群落与小黄花茶的功能关系。因此,下一步的研究应从基础生物学和生态学角度研究小黄花茶生殖生理、种群更新与繁殖障碍,并结合岛屿地理生物学更大尺度探索小黄花茶种群濒危衰退机制。 在本研究中,小黄花茶在群落中具有最大的重要值及生态位宽度值,与其它伴生物种的生态位相似性及重叠程度均较低且种间联结性较弱。优势种生态位相似性及重叠程度均较低;群落总体联结(VR、W)与种间联结(χ2、AC、PC)分析结果均表明各优势物种间呈不显著负联结。线性回归分析显示优势物种的AC值、PC值与生态位相似比、生态位重叠值之间呈极显著正相关关系,表现为优势物种的种间正(负)联结性越强,生态位相似性与重叠程度越高(低)。这些结果表明,赤水桫椤国家级自然保护区分布的小黄花茶在其局域群落与伴生的优势物种间关联性松散,群落各优势种群间相对独立,群落稳定性较弱。
2.3 优势植物总体联结性分析
2.4 优势植物种间联结性分析



2.5 优势植物种间联结与生态位回归分析
3 讨论
3.1 小黄花茶种群生态位
3.2 小黄花茶群落的种间联结性
3.3 小黄花茶种群保育与研究展望
4 结论
