不同品种油茶可培养内生真菌多样性研究
2024-01-16何刚丁薇欧阳建萍张红芳孙启彪季晓红陈晔
何刚,丁薇,欧阳建萍,张红芳,孙启彪,季晓红,陈晔*
(1.九江学院 药学与生命科学学院,江西 九江 332005;2.九江市真菌资源保育及应用重点实验室,江西 九江 332005;3.江西省长江流域产业生态模拟与环境健康重点实验室,江西 九江 332005)
植物内生真菌是一类广泛存在于健康植物中的应用前景广阔的资源微生物,具有丰富的物种多样性和生态分布多样性[1]。植物种类不同其植物内生真菌多样性和群落结构也不尽相同[1-2]。此外,同种植物组织器官、生长时期、生长环境不同,其内生真菌的种类和数量存在差异[1-3],而且内生真菌在野生种和栽培种之间的定殖情况亦不同[4]。植物内生真菌与宿主长期协同进化过程中建立了互惠共生关系[5],不仅能有效促进宿主植物生长,提升宿主植物的抗病虫害能力和抗逆能力[6-9],还对维持宿主植物微生态系统动态平衡和增强其生态适应性具有重要作用[10]。
油茶(Camellia oleiferaAbel.)又称普通油茶或茶子树,是山茶科(Theaceae)山茶属(Camelliaspp.)植物,为我国南方特有木本油料树种,至今已有2 300多年栽培和利用历史[11]。普通油茶主要分布于我国湖南、江西等南方省区,其中由中国林业科学研究院亚热带林业研究所主持选育的长林系列是高产、稳产优良油茶品种[12]。油茶籽榨取的茶油是一种优质食用油,因富含油酸、亚油酸等不饱和脂肪酸和丰富的维生素被誉为“东方橄榄油”,同时茶油及其副产品在工业、农业、医药等方面具有多种用途[13]。此外,油茶还是水土保持、水源涵养、环境美化和生物防火的重要树种[11],具有很高的综合利用价值。近年来,油茶相关研究主要集中在油茶品种选育、油茶主要成分与质量分析、油茶深加工、油茶根际微生物群落和有害生物防治等方面[14-15]。虽然有关油茶内生真菌已有少许研究报道[16-18],但是关于普通油茶栽培和野生可培养内生真菌的种类、组成等方面工作鲜见相关报道。因此,有必要弄清不同品种油茶内生真菌群落结构和应用潜能。
本研究以普通油茶野生和长林系栽培品种根系和叶片为材料,在内生真菌分离、鉴定基础上比较分析普通油茶野生和栽培内生真菌的属种组成和多样性,为进一步揭示油茶内生真菌群落生态学意义和挖掘利用有益功能菌株资源提供参考。
1 材料与方法
1.1 实验材料
普通油茶野生植株根系和叶片材料于2021年4月采自江西省九江市庐山国家级自然保护区(29°63′N,116°05′E);长林3号、长林4号、长林53号等3个长林系普通油茶栽培品种植株根部材料于2021年3月采自江西省九江市德安县神州通油茶林泉基地。野外采样按照五点采样法随机选取相隔一定距离的健康植株,采集的样品放入无菌采样袋置于低温冷藏箱,带回实验室后放入4℃冰箱保藏,并在24 h内处理样品进行内生真菌分离。
马铃薯葡萄糖琼脂培养基(Potato dextrose ager, PDA)用于内生真菌分离、纯化和菌株保藏。
1.2 方法
1.2.1 内生真菌定殖观察
先将采集的油茶根清洗干净,再选取直径小于2 mm的细根剪成长度5 mm左右的小段。依次将上述根段置于10% KOH溶液90℃水浴1 h,然后蒸馏水洗涤3次,再将根段置于10% H2O2溶液中室温放置30 min之后使用蒸馏水洗涤,最后将根段置于1%HCl溶液浸泡5 min并清洗干净[19];期间根据油茶根段软硬程度适当调整浸泡试剂时间。经处理的根段采用台盼蓝染液染色。每个品种随机选取20个根段,通过光学显微镜观察并记录内生真菌定殖情况。
1.2.2 内生真菌分离纯化
按照参考文献方法对油茶叶和根进行消毒[20]。分别将叶片剪成边长5 mm左右的正方形小块,将根剪成长度为5 mm左右的小段。采用组织块分离法从油茶根和叶片分离内生真菌,组织块接种于PDA培养基,置于25℃培养箱黑暗培养。每天观察,待有菌丝从组织块边缘长出后,及时挑取单一菌丝转接至PDA培养基。通过菌落的形态特征进行区分,若菌落形态特征不一致,则继续挑取菌落边缘单一菌丝接入PDA培养基进行纯化培养,直至获得单一的菌种。纯化后的内生真菌菌株保存于九江学院九江市真菌资源保育及应用重点实验室。
1.2.4 菌株鉴定
1.2.4.1 形态学鉴定
观察记录菌落形态特征和生长特性,以及菌丝、产孢结构、孢子等微观形态特征,并参考《真菌鉴定手册》等相关文献进行鉴定[21]。
1.2.4.2 分子生物学鉴定
采用氯化苄法提取内生真菌菌丝基因组DNA[22]。采用真菌通用引物ITS4和ITS5进行PCR扩增[23],扩增产物送擎科生物进行测序。所得序列在美国国家生物信息技术中心(NCBI)中进行同源性比对和分析,确定菌株分类地位,再选取代表菌株序列利用MEGA 11.0软件的最大似然法(Maximum likelihood, ML)进行属级水平的分子系统发育分析[20]。
1.3 数据统计分析
采用定殖率(Colonization rates, CR)衡量植物组织中内生真菌的丰富程度;采用分离率(Isolation rate,IR)反映组织块受侵染的发生频率;采用相对频率(Relative frequency, RF)分析内生真菌的优势菌群[20]。
采用Sorenson相似性指数(Sorenson’s similarit coefficiens, Cs)对内生真菌组成的相似程度进行比较和分析[24]。采用Shannon-Weiner多样性指数(H)进行内生真菌种群的多样性水平分析[20]。
式中:j代表2个样本共有真菌数量,a和b分别代表2个样本中各自分离到的真菌数量。
2 结果与分析
2.1 油茶根系真菌侵染定殖形态特征
采用台盼蓝染色后,观察油茶根内生真菌定殖情况,结果表明内生真菌在不同油茶品种中定殖存在一定的差异,长林3号定殖率为95%,长林4号为100%,长林53号为95%,野生小果油茶为100%。从图1中可观察到内生真菌在油茶根内细胞定殖的菌丝、泡囊、微菌核等形态结构特征。
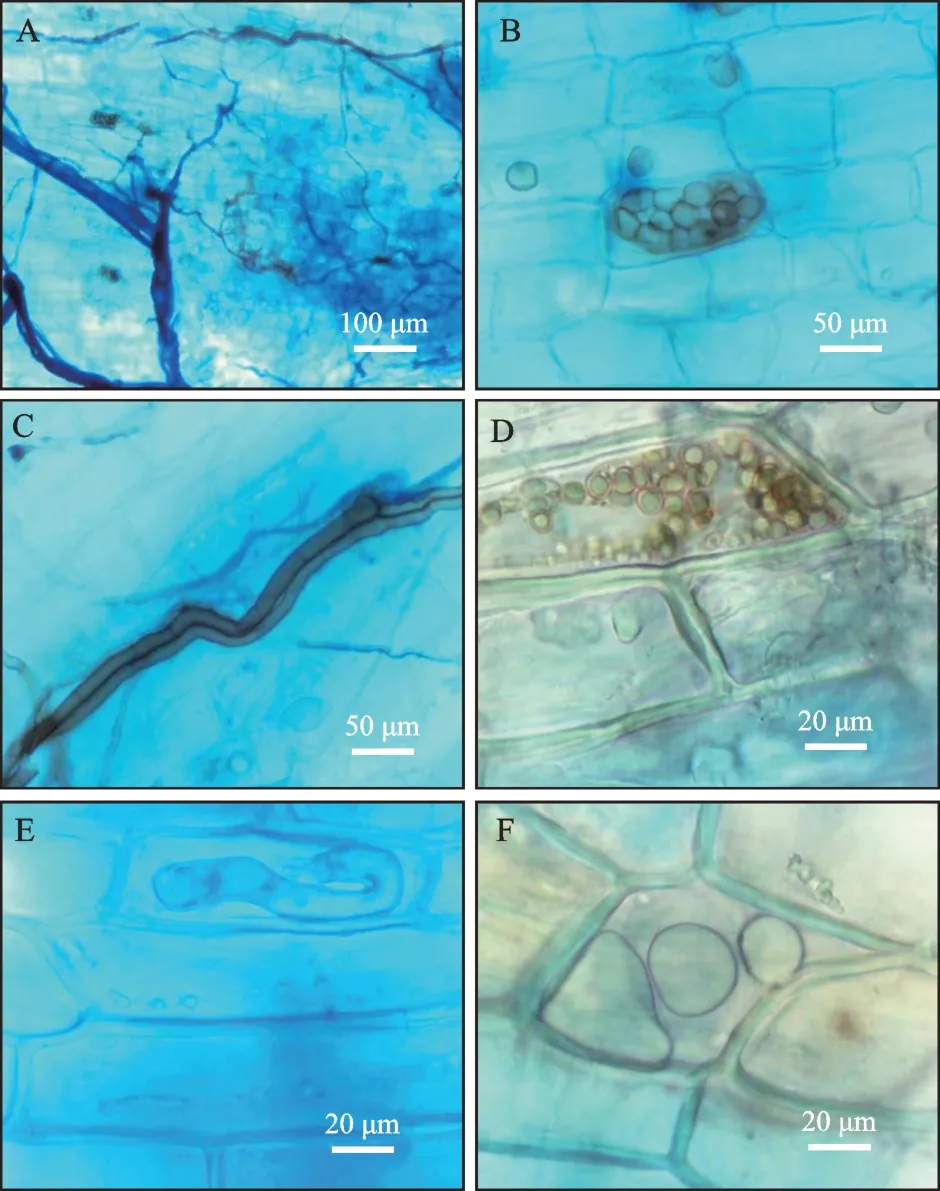
图1 内生真菌在根内定殖的镜检图片Fig.1 Microscopic images of endophytic fungi colonizing the roots

图2 基于ITS基因序列借助最大似然法构建的油茶内生真菌系统发育树Fig.2 Molecular phylogenetic tree of endophytic fungi isolated from C. oleifera based on ITS gene sequences using maximum likelihood method
2.2 内生真菌分离菌株数和分离率
由表1可知,从1 440个油茶组织块中共分离获得菌株数864株,总分离率为60.00%,其中从叶片组织中获得631株(占总菌株数73.03%)、根部获得233株(占总菌株数26.97%),分离率分别为87.64%和32.36%,可见从油茶不同部位分离获得的菌株数量存在较大差异。
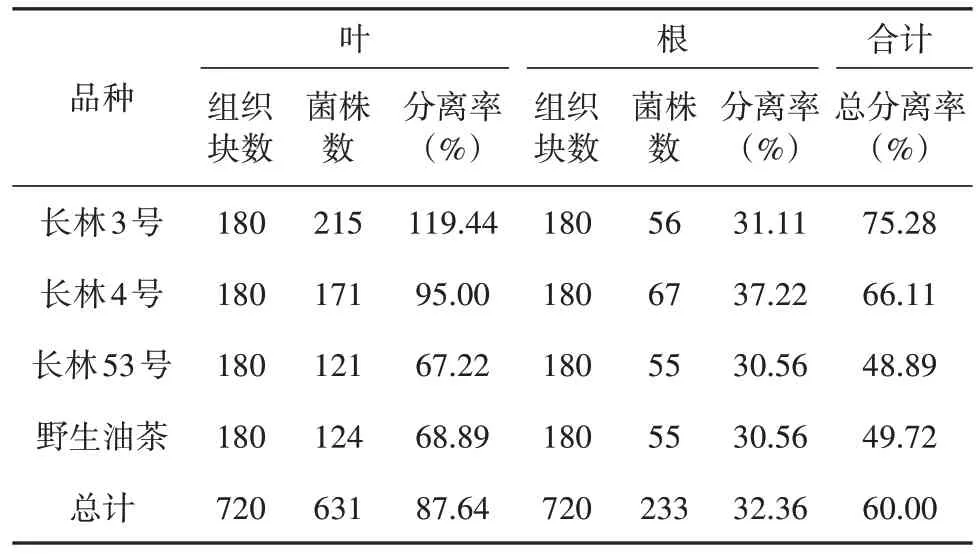
表1 不同品种油茶内生真菌分离结果Tab. 1 Isolation results of endophytic fungi from different varieties of C. oleifera
同时,不同油茶品种之间分离获得内生真菌菌株数量也有差别。四个品种油茶的内生真菌总分离率为长林3号(75.28%) > 长林4号(66.11%) > 野生小果油茶(49.72%) > 长林53号(48.89%),其中长林3号获得菌株数最多为271菌株,长林53号获得菌株数最少为176株。此外,同一器官组织在不同品种间内生真菌分离率也存在差异,如叶在长林3号品种分离率最高为119.44%,长林53号最低为67.22%;而根在长林4号品种中分离率最高为37.22%,长林53号和野生小果油茶最低,均为30.56%。
2.3 内生真菌主要组成群落分析
将从不同油茶品种1 440个组织块分离获得的864株内生真菌菌株合并为118个不同形态型,每个形态型各选择1株代表菌株经分子鉴定后共划分为64个分类单元。根据基于ITS序列构建的系统发育树(图 2)可知,油茶叶和根的内生真菌物种种类丰富,主要形成了四个分支且叶内生真菌与根内生真菌在属级物种组成上存在较明显差别。
所获内生真菌菌株隶属于6纲14目26科39属(表2)。按相对频率,在属级水平上内生真菌优势属为炭疽菌属、间座壳属、链格孢属,相对频率分别为18.63%、16.78%、10.53%;葡萄座腔菌属、青霉属(Penicillium)、曲霉属(Aspergillus)相对频率在5% ~ 10%之间,其余均低于5%(图3)。
由表2可知不同油茶品种内生真菌优势类群存在明显差异。长林3号优势属为炭疽菌属、间座壳属,相对频率分别为9.26%、3.59%,特有种是肉色隔孢伏革菌(Peniophora incarnata)、大孢树粉孢(Oidiodendron maius)等。长林4号优势属为炭疽菌属、葡萄座腔菌属,相对频率分别为5.32%、4.86%,特有种是Neofusicoccum batangarum、蜜柑间座壳菌(Diaporthe unshiuensis)等。长林53号优势属为链格孢属、间座壳属,相对频率均为3.82%,特有种是Periconia igniaria、Hypoxylon commutatum等。野生油茶优势属为间座壳属、链格孢属,相对频率分别为6.25%、2.66%,特有种是扩散炭垫菌(Nemania diffusa)、橘青霉(Penicillium citrinum)等。
同时,油茶不同部位内生真菌优势属也不尽相同。油茶叶内生真菌优势属为炭疽菌属、间座壳属,相对频率分别为14.58%、13.66%;油茶根内生真菌优势属是曲霉属、青霉属,相对频率分别为6.60%、4.86%。由此可见,油茶不同部位内生真菌群落组成有较大差异。此外,尽管不同油茶品种在同一部位内生真菌的优势属近似,但物种组成有一定差异,栽培油茶和野生油茶各有其特有物种。油茶的品种、生存环境、栽培条件、土壤条件及周围植物组成等可能是影响油茶内生真菌群落组成差异的主要因素。
2.4 真菌相似性及多样性分析
2.4.1 真菌相似性分析
从相似性系数看栽培品种油茶与野生油茶分离获得的内生真菌发现(表3),长林3号与长林4号相似性系数最高(Cs = 0.51),长林53号与野生油茶相似性系数较高(Cs = 0.50),长林4号与野生油茶相似性系数最低(Cs = 0.24)。另外长林3号与长林53号的内生真菌相似性系数偏中等(Cs = 0.45);长林3号与野生油茶相似性系数不高(Cs = 0.39);长林4号与长林53号相似性系数较低(Cs = 0.35)。

表3 不同品种油茶内生真菌相似性系数Tab. 3 Similarity Coefficient of endophytic fungi in different varieties of C. oleifera
2.4.2 真菌多样性分析
从Shannon-Wiener 指数看栽培品种油茶与野生油茶分离获得的内生真菌多样性发现(图4),长林4号(H = 1.22)和长林3号(H = 1.18)真菌多样性较高,长林53号(H = 0.91)和野生油茶(H = 0.83)真菌多样性较低。

图4 不同品种油茶真菌多样性指数Fig.4 Diversity index of fungi in different Camellia oleifera varieties
3 结论与讨论
本研究通过采用形态特征和分子系统学证据相结合的方法对分离获得的864株内生真菌进行鉴定,结果表明不同品种油茶可培养内生真菌具有较丰富的物种多样性,且油茶不同部位的内生真菌不仅具有多样性和差异性,其分布也呈现一定组织特异性;然而,油茶不同品种对油茶内生真菌多样性和分布影响有限,这与已有研究报道一致[16-18]。本研究所涉油茶品种中,长林3号内生真菌分离率最高,长林3号与长林4号之间的内生真菌相似性系数最高,长林4号内生真菌多样性指数最高;分离所得内生真菌优势属为炭疽菌属、间座壳属、链格孢属。此外,野生油茶与栽培品种内生真菌物种组成存在一定差异,如野生油茶特有菌株为扩散炭垫菌、橘青霉等,而栽培油茶特有菌株则有肉色隔孢伏革菌、Neofusicoccum batangarum等,这种差异是否由人工栽培条件引起还有待进一步探究。
本研究还发现,对于油茶根和叶而言,不仅内生真菌的分离率差异明显,群落组成也不尽相同。即使油茶品种不同,在相同组织部位分离得到的内生真菌优势属也较相似,但同一油茶品种不同组织部位分离得到的内生真菌属种组成存在较大差异。其中,油茶叶的内生真菌分离率较高,且多为不产孢真菌,少数有气生菌丝;油茶根的内生真菌分离率较低,且多为产孢菌株。
油茶具有较高经济价值,系统研究各品种油茶内生真菌多样性和群落组成可为深入探究内生真菌对油茶生长影响机制及利用油茶资源提供理论参考。
