灵武长枣叶阿拉伯半乳糖蛋白分布及光合特性
2024-01-13陶珊珊章英才
陶珊珊, 章英才, 王 静
(宁夏大学 生命科学学院,宁夏 银川 750021)
阿拉伯半乳糖蛋白(AGPs)为广泛分布于植物各器官细胞中的糖蛋白,由阿拉伯糖和半乳糖构成多糖支链和蛋白质核心链组成结构复杂的大分子[1-3]。作为植物细胞壁的一种结构蛋白,AGPs 与植物适应逆境有关[4]。叶是植物对环境变化比较敏感且可塑性较大的器官,叶的AGPs 分布性状直接影响到叶的功能。研究表明,枣叶中有丰富的黄酮类[5-6]、三萜类[7]、糖类和皂苷等[8],不同发育时期灵武长枣(Ziziphus jujubaMill ‘Lingwuchangzao’)叶营养成分含量与果实营养成分和果实品质间存在明显的相关性[8];AGPs 参与了灵武长枣果实维管束发育过程的形态建成、细胞分裂和体积增大,并为果实发育提供营养支持及保护[2,9],因此叶的AGPs 分布与叶的干旱适应特征相关,并有可能对果实品质的形成发挥重要的影响。采用分子生物学等技术对植物细胞中AGPs 糖蛋白的研究表明,AGPs 在生殖发育过程,包括花粉发育、花粉管生长、自交不亲和、大孢子母细胞形成,果实的成熟和衰老过程[9-11],以及根和茎等营养器官的发生与发育等方面起着重要作用[2,12]。研究表明,宁夏枸杞多糖及果实的细胞壁多糖是阿拉伯半乳聚糖AGPs 糖蛋白,是枸杞子中具有免疫调节功能的药用有效成分[10-11];番茄LeAGP-1基因过量表达的植株株高减少、分枝增多、种子数量减少、果实减小,表明此AGPs 参与了营养生长和生殖发育[13];过量表达黄瓜(Cucumis sativus)CsAGP1 基因的烟草植株株高增加,说明AGPs 促进了茎的生长[14];拟南芥AtAGP30 对种子萌发和根的发育起重要的调控作用[15],AtFLA1 促进根的发育和细胞分化[16],AtFLA11、AtFLA12参与次生细胞壁的发育[17-19],与木质素形成基因同源的AtXYP1、AtXYP2与维管束的发育密切相关[20],AtAGP31 参与维管束的发育与逆境应答[21-22],AtAGP19基因在细胞分裂与生长、叶的发育以及生殖发育过程中起着重要作用[23]。基于AGPs 相关基因的表达模式证实了多种类型的AGPs 基因在不同组织部位的特异性表达,在植物营养器官和繁殖器官发育中起非常重要的作用,因此探明AGPs 在不同组织部位分布的特征尤其重要[2-3]。AGPs 存在于植物各器官中,利用AGPs的特异性抗体进行识别,是研究不同植物中不同部位AGPs 分布与特性的重要手段,已在枸杞果实AGPs 免疫定位[9-11],低温引起雏菊(Bellis perennis)胚珠和花药发育过程中AGPs 分布的变化[4],利用AGPs 单克隆抗体JIM8、JIM13、MAC207、LM2 对拟南芥有性生殖过程中AGPs 进行定位的研究中取得了成果[2-3,24]。在营养器官方面,JIM4 识别的AGPs 与胡萝卜根早期维管组织的发育有关,JIM14 识别的AGPs 与根次生加厚的筛管有关[25],JIM13 识别的AGPs 定位在幼嫩的木质部和根冠[26],LM2、LM14 识别的AGPs 定位在野生型大麦根毛的细胞壁和细胞质中[27]。由此说明,不同的单克隆抗体识别的AGPs 在根组织发育中有着特定的分布,但目前对叶发育中AGPs 分布的研究较少。通过针对性抗体对AGPs 进行定位的免疫学方法具有较高的特异性,因此成为研究不同植物中不同部位AGPs分布的较好的研究途径[2-3]。灵武长枣是鼠李科枣属的鲜食枣品种,原产于宁夏灵武市,耐干旱和盐碱环境,果实具有优良的药用品质和食用品质。近年来,对灵武长枣果实的研究取得了丰富的成果[2-3,9],而有关灵武长枣叶的AGPs 糖蛋白分布及其干旱适应特征以及对果实品质的影响尚不明了。本试验以不同发育时期灵武长枣叶为材料,应用免疫荧光定位技术,较系统地研究不同时期叶的AGPs糖蛋白分布、光合特性及其干旱适应特征,旨在为进一步研究叶AGPs 在灵武长枣生长发育及果实品质方面的功能奠定基础。
1 材料与方法
1.1 试验材料
试验材料采自宁夏灵武市宁夏红枣工程技术研究中心试验基地6年生灵武长枣树。试验采用随机设计,设3 次重复,每次重复选择生长发育良好、树势适中、长势相近、栽培管理水平一致的5~10 株植株[2-3,9],使用3种不同颜色的毛线于6月10日标记同一天开放的花朵,每个重复标记花朵3 000 朵,分别在谢花后30、60、90、110 d,即分别在果实膨大前期(7 月10 日)、快速膨大期(8 月9 日)、着色期(9 月8日)、完熟期(9月28日)取样,共取样4次。每次取样均在09:00—11:00进行。在试验植株树冠的东、西、南、北4个方位及上、中、下、里、外各个方向选择枣吊中部果柄附近的叶[2-3,9]。同时测定光合速率及可溶性糖含量等指标。
AGPs 单克隆抗体JIM8 购自美国乔治亚大学,碱性磷酸酶标记的山羊抗小鼠IgG 二抗购自Sigma-Aldrich公司[3,9]。
1.2 试验方法
参照Bao等[11]、王莹莹[28]的方法,略有改动。
1.2.1石蜡切片的制作 选取各个不同发育时期的叶,用锋利的刀片将叶片分割成含有3 条主脉的约长0.6 cm×宽1 cm 的小片,立即放入含有φ(甲醛)=3.7%的2F4 固定液中,盖紧瓶盖后用注射器抽气至样品下沉瓶底,室温固定4 h。更换固定液3次,每次30 min。用PBS 磷酸缓冲液(10 mmol/L,pH=7.0)清洗3 次,每次15 min。样品在4 ℃下用φ=0.05% Toluidine blue 染色15 min,在4 ℃下依次在φ=30%、50%、70%、90%的乙醇梯度下脱水,每次60 min,φ(乙醇)=100%脱水3~4 次,60 min/次。在37 ℃恒温箱中将样品用φ(乙醇)∶φ(Steedman’s wax)=1∶1渗透过夜,次日更换φ(乙醇)∶φ(Steedman’s wax)=1∶3 在37 ℃下渗透2.5 h,再用纯Steedman’s wax 在37 ℃下渗透3 次,每次2 h。将样品用纯Steedman’s wax 在牛皮纸盒中包埋,凝固即可。后期蜡块的固着、整修、切片、贴片和展片等步骤参考王静等[2-3,9]的方法。
1.2.2糖蛋白免疫荧光定位 切片脱蜡、复水、PBS淋洗浸泡、封闭液处理等参考王静等[2-3,9]的方法。用PBS(φBSA=1%)以1∶100 比例稀释后的AGPs 单克隆抗体JIM8,4 ℃下孵育过夜,次日用PBS淋洗浸泡3 次,每次10 min,以去除多余抗体,再用PBS(φBSA=1%)以1∶300 比例稀释后的碱性磷酸酶标记的山羊抗小鼠IgG 二抗在36 ℃、黑暗条件下孵育1 h。标记后,用PBS 淋洗浸泡除去未标记的二抗,φ=0.01%Toluidine blue染色去除植物本身的自发荧光,经PBS淋洗浸泡后封片[2-3,9]。在Leica STELLARIS 5 激光共聚焦显微镜下观察切片并拍照。
1.2.3叶光合速率等指标的测定 利用CIRAS-1型光合测定仪测定果柄附近叶片的净光合速率(Pn,μmol·m-2·s-1)、蒸腾速率(Tr,mmol·m-2·s-1)、气孔导度(Gs,mmol·m-2·s-1)、胞间CO2浓度(Ci,μmol·mol-1)等光合参数。叶片光合色素叶绿素质量比按常规方法进行测定。
1.2.4可溶性糖的提取和质量比测定 参考郑国琦等[29]的方法,分别称取不同发育时期的多个叶片在45 ℃烘干粉碎后的混合样品,加提取液(V乙醇∶V氯仿∶V水=12∶5∶3),再匀浆3~5 min,5 000 g 离心15 min,取上清液,重复3 次。合并提取液,转入分液漏斗,加水使之分层,5 000 g 离心10 min 去除氯仿层,用c(NaOH)=0.1 mol·L-1调pH 至7.0,在45 ℃真空中干燥,用蒸馏水定容。使用高效液相色谱仪(LC-20AT)测定葡萄糖、果糖和蔗糖的质量比,色谱条件:流动相(V乙腈∶V重蒸水=85∶15),流速1.0 mL·min-1,氨基柱,柱温30 ℃,RID-10A 示差检测器,LC solution数据处理系统。
利用SPSS 22.0 和Excel 2010 软件进行数据统计处理。
2 结果与分析
2.1 花后30 d叶AGPs的分布特征
1)表皮。叶上表皮细胞较大,近长方形,下表皮细胞较小,方形,垂直于叶片方向的细胞壁较薄,抗体所识别的抗原荧光AGPs 分布较少,而表皮细胞外切向壁较厚,抗体所识别的抗原荧光AGPs 分布较多,形成了较厚的角质层;气孔仅分布于下表皮,保卫细胞中分布着少量AGPs(图1A—图1D)。
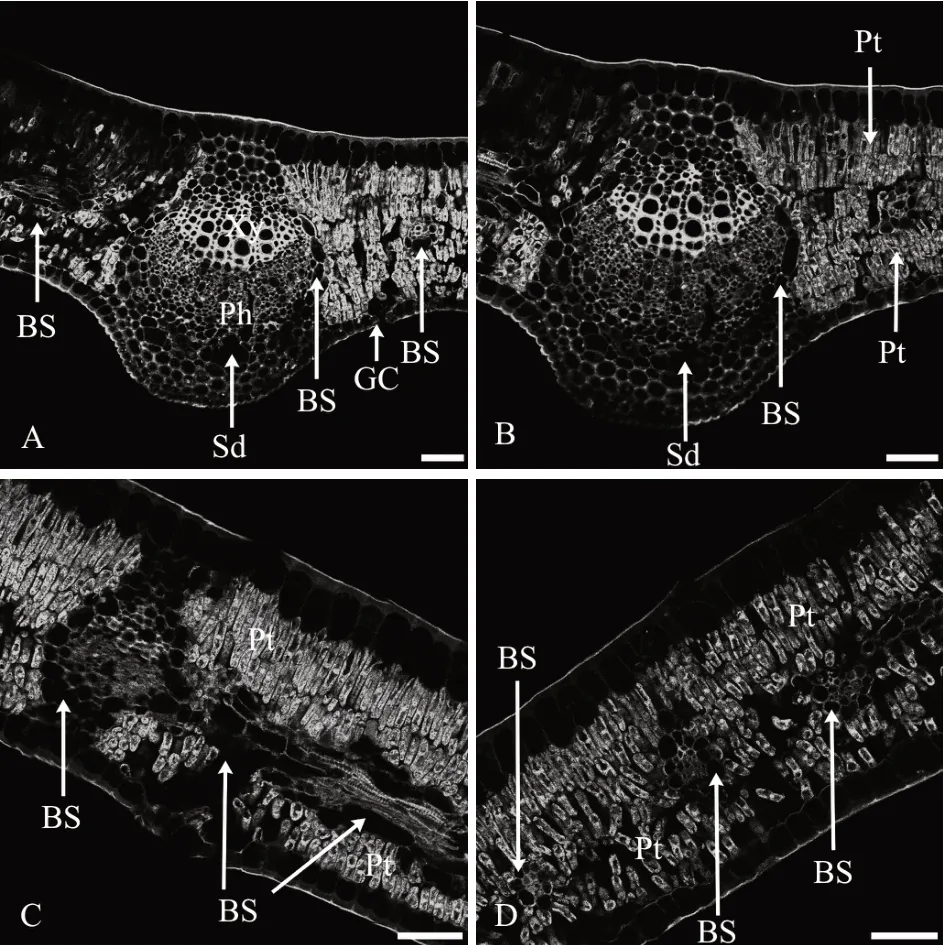
图1 花后30 d叶AGPs免疫荧光分布
2)叶肉。靠近上表皮的叶肉组织由3层垂直于上表皮的长柱状栅栏组织细胞组成,排列很紧密;近下表皮的叶肉组织由3~4层短柱状的栅栏组织细胞组成,排列也较紧密,为典型的等面叶,具有较典型的旱生结构特征,栅栏组织细胞壁和细胞内部均密集分布着抗体所识别的抗原荧光AGPs(图1A—图1D)。
3)叶脉。叶具有3 条发达的主脉,主脉维管束木质部导管和薄壁组织细胞的细胞壁上密集分布着大量抗体所识别的抗原荧光AGPs,在近形成层的木质部中AGPs 尤其丰富(图1A—图1B);形成层和韧皮部排列紧密的细胞壁和细胞内部均密集分布着抗体所识别的抗原荧光AGPs。维管束外由一圈卵圆形的分泌细胞形成维管束鞘,细胞中基本没有抗原荧光AGPs分布(图1A—图1B)。
主脉处的表皮之下为多层厚角组织,细胞壁分布着较多抗体所识别的抗原荧光AGPs,厚角组织内部圆形或卵圆形较大的薄壁细胞壁上也分布着较多抗体所识别的抗原荧光AGPs,其中分布有数个分泌道,内部没有AGPs荧光分布(图1A—图1B)。
叶肉中分布着横切或纵切比较小的结构简单的侧脉和细脉。外面是一圈含分泌物的薄壁细胞组成的维管束鞘,细胞中没有抗原荧光AGPs 分布,内部木质部和韧皮部细胞的细胞壁和细胞内部均密集分布着抗体所识别的抗原荧光AGPs(图1C—图1D)。可见,叶肉是叶AGPs分布的主要部位。
2.2 花后60 d叶AGPs的分布特征
1)表皮。表皮细胞外切向壁较厚,抗体所识别的抗原荧光AGPs 分布较多,形成较厚的角质层;垂直于叶片方向的表皮细胞壁较薄,抗体所识别的抗原荧光AGPs 分布较少,气孔保卫细胞中分布着少量AGPs(图2A—图2D)。
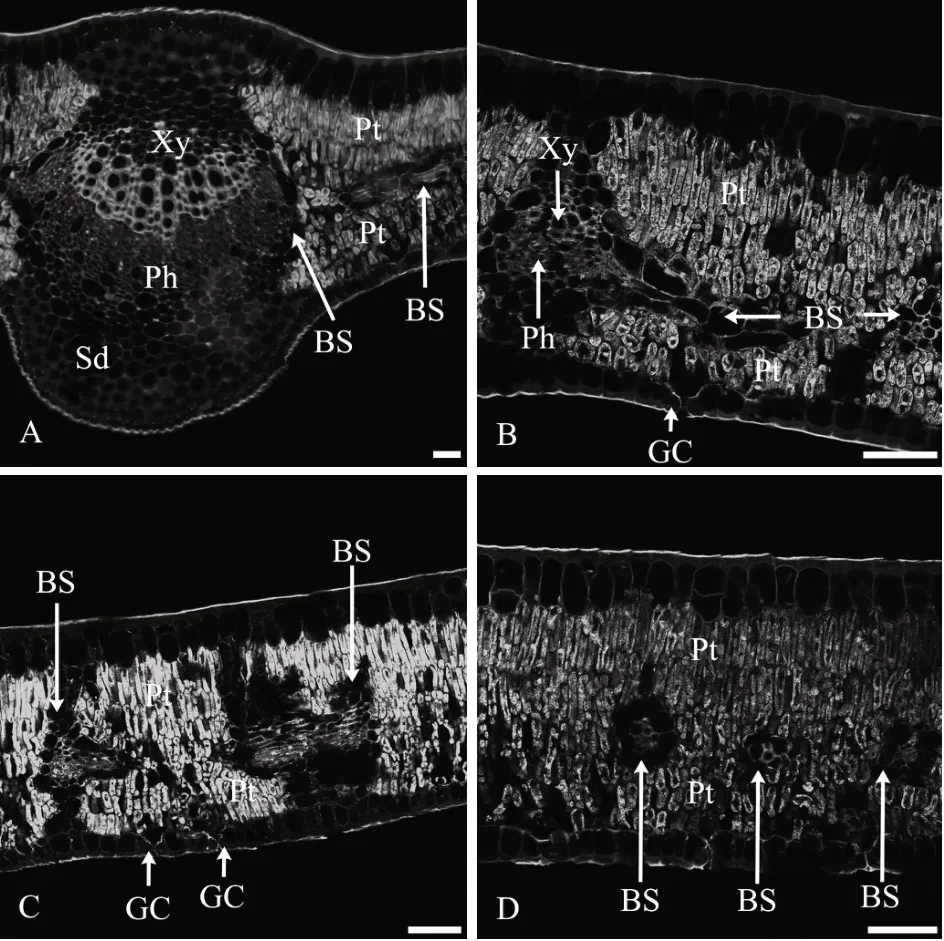
图2 花后60 d叶AGPs免疫荧光分布
2)叶肉。叶肉栅栏组织细胞的细胞壁和细胞内部均密集分布着抗体所识别的抗原荧光AGPs,AGPs分布特征与花后30 d的叶相似,叶肉栅栏组织是叶AGPs分布的主要部位(图2A—图2D)。
3)叶脉。主脉维管束在木质部导管和薄壁组织细胞的细胞壁上分布着大量抗体所识别的抗原荧光AGPs,形成层、韧皮部细胞壁和细胞内部分布着较多抗体所识别的抗原荧光AGPs;与花后30 d的叶相似,维管束外由分泌细胞形成的维管束鞘细胞中基本没有抗原荧光AGPs分布(图2A)。
主脉的上下表皮之内的厚角组织细胞壁和细胞内部,以及紧邻其内的圆形或卵圆形较大薄壁细胞的细胞壁和细胞内部均分布着抗体所识别的抗原荧光AGPs,但相比花后30 d 的叶略有减少(图2A);上下表皮厚角组织内部薄壁细胞中分布的分泌道中均没有AGPs荧光(图2A)。
与花后30 d 的叶相似,叶肉中间部位分布的结构简单的侧脉和细脉,外面含分泌物的维管束鞘薄壁细胞中没有抗原荧光AGPs,内部木质部和韧皮部细胞的细胞壁和细胞内部均密集分布着抗体所识别的抗原荧光AGPs(图2B—图2D)。
2.3 花后90 d叶AGPs的分布特征
1)表皮。花后90 d 叶上、下表皮细胞壁上抗体所识别的抗原荧光AGPs 分布情况与花后60 d 叶相似,表皮细胞外切向壁较厚,抗体所识别的抗原荧光AGPs 分布较多,形成了较厚的角质层(图3A—图3D)。
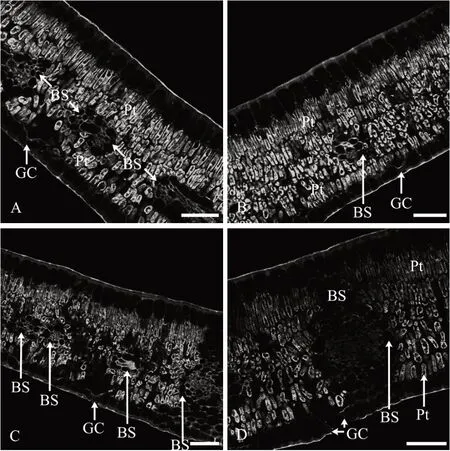
图3 花后90 d叶AGPs免疫荧光分布
2)叶肉。与之前时期的叶略有不同,花后90 d叶的叶肉组织细胞排列更加密集,甚至邻近下表皮处的栅栏组织细胞排列也非常紧密,而且叶肉栅栏组织细胞的细胞壁和细胞内部均密集分布着大量抗体所识别的抗原荧光AGPs(图3A—图3D)。
3)叶脉。主脉AGPs 的分布特征与之前时期的叶相似,侧脉和细脉维管束木质部和韧皮部细胞的细胞壁和细胞内部均密集分布着抗体所识别的抗原荧光AGPs(图3A—图3D);维管束外的维管束鞘细胞中基本没有抗原荧光AGPs 分布(图3A—图3D)。
2.4 花后110 d叶AGPs的分布特征
1)表皮。花后110 d 叶上、下表皮细胞壁上抗体所识别的抗原荧光AGPs 的分布情况与之前时期的叶相似,表皮细胞外切向壁抗体所识别的抗原荧光AGPs 分布较多,形成了较厚的角质层(图4A—图4D)。
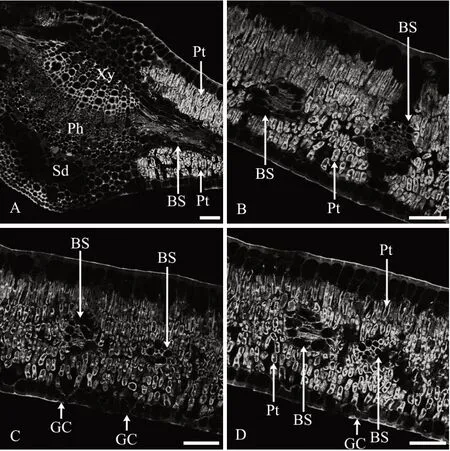
图4 花后110 d叶AGPs免疫荧光分布
2)叶肉。花后110 d 叶肉栅栏组织细胞与花后90 d 的叶相比排列更加紧密,栅栏组织细胞的细胞壁和细胞内部均密集分布着大量抗体所识别的抗原荧光AGPs(图4A—图4D)。
3)叶脉。与花后90 d 的叶相似,主脉维管束木质部导管和薄壁组织细胞的细胞壁上密集分布着大量抗体所识别的抗原荧光AGPs,而韧皮部细胞壁和细胞内部分布着较多抗体所识别的抗原荧光AGPs(图4A)。主脉厚角组织、薄壁细胞及分泌道AGPs 分布特征与花后90 d 的叶相似(图4A)。侧脉和细脉维管束木质部和韧皮部细胞的细胞壁和细胞内部均密集分布着抗体所识别的抗原荧光AGPs(图4B—图4D)。各叶脉维管束鞘细胞中均没有抗原荧光AGPs分布(图4A—图4D)。
2.5 不同发育时期叶的光合特性及可溶性糖质量比的变化
1)叶的净光合速率基本呈高低交替的变化趋势,花后30 d处于较高水平,之后有所下降,花后90 d达到最高水平,再次呈现峰值,至花后110 d 有一定下降,果实发育成熟(图5A)。
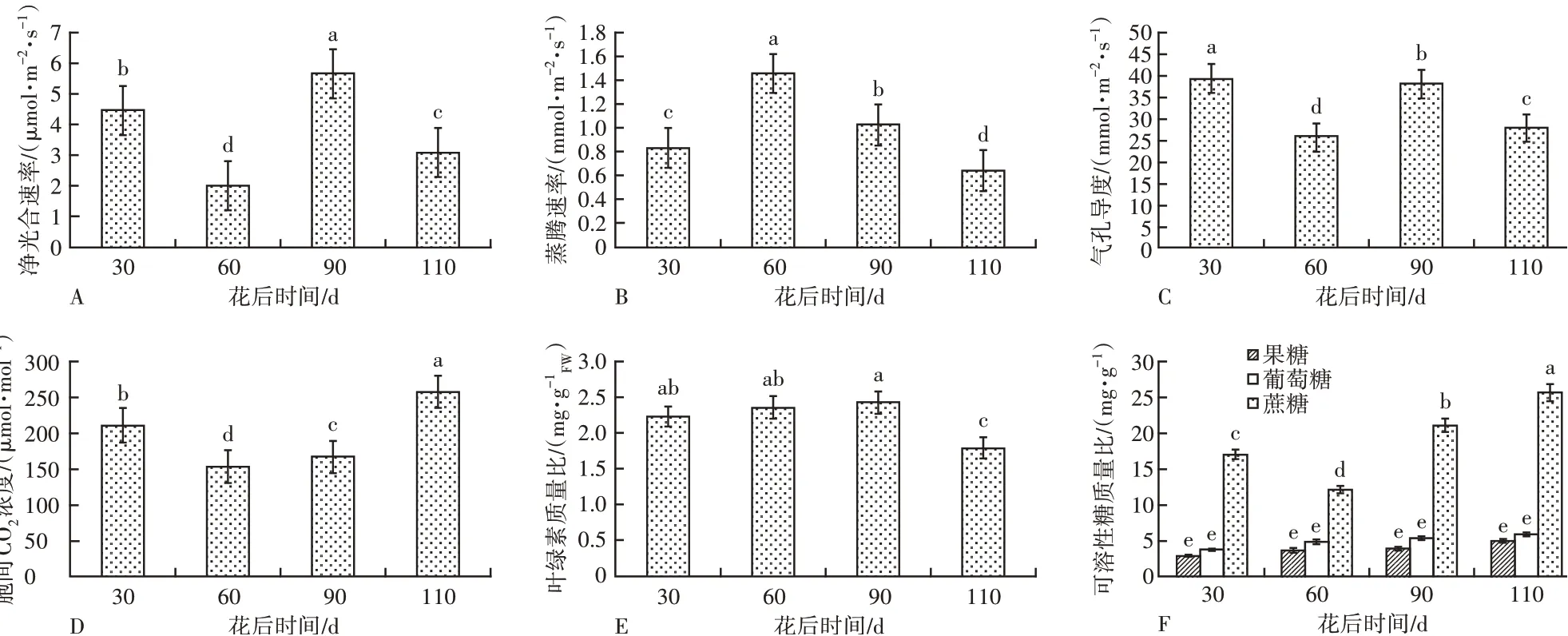
图5 不同发育时期叶光合特性及可溶性糖质量比的变化
2)叶的蒸腾速率在花后30 d 处于相对较低的水平,在花后60 d 达到最高值,花后90~110 d 逐渐降低,这与气温的变化规律相适应(图5B)。
3)叶的气孔导度呈现高低交替的变化特征,与净光合速率的变化趋势相似,但气孔导度在花后30 d处于最高水平,在花后90 d再次出现峰值(图5C)。
4)叶的胞间CO2浓度在花后30 d 相对较高,花后60 d 降至最低,而后升高,至花后110 d 出现峰值,这与蒸腾速率的变化趋势相反(图5D)。
5)叶中叶绿素质量比始终处于较高水平,在花后30~90 d,叶绿素质量比逐渐上升到最大值,在花后110 d叶绿素质量比略有下降(图5E)。
6)叶中葡萄糖、果糖和蔗糖的质量比分别呈现出不同的变化趋势,在生长发育过程中,葡萄糖和果糖的质量比变化趋势相似,均为逐渐上升,且葡萄糖的质量比始终高于果糖,2 种糖的积累总量较为平缓,在花后110 d 质量比出现峰值。蔗糖的质量比随着叶的发育呈先下降后逐渐上升的趋势,在花后30~60 d 逐渐下降,花后60 d 以后蔗糖质量比不断升高,至花后110 d 质量比出现峰值,蔗糖质量比的增长速度和净增长量始终远高于葡萄糖和果糖,可见叶片以积累蔗糖为主(图5F)。
3 讨论与结论
AGPs 是一类高度糖基化的蛋白质[2-3],作为植物细胞壁的一种结构蛋白,AGPs与植物适应逆境有关[4],在植物抗冷害机制中发挥着重要作用,与植物冷害存在重要相互关系,被证明在胁迫下参与信号传导和细胞壁代谢[30]。研究表明,棉花根部组织中的1 个AGPs 基因GhAGP31 在冷胁迫下表达量显著上升,表明AGPs 参与了植物的抗寒过程[31];香蕉中的AGPs 基因MaFLA2-1、MaFLA17-3、MaFLA2-2 的表达量可能与香蕉的抗冷性密切相关,AGPs抗原可能参与了香蕉的抗冷过程,这揭示了AGPs 与植物冷害的关系,为香蕉抗冷育种提供了参考[28]。近年来还发现,AGPs 在植物应对生物[32]及盐胁迫等非生物胁迫过程中也扮演着重要角色[33],Lamport 等[33]利用盐胁迫分析AGPs 功能认为,盐胁迫使烟草BY-2细胞中AGPs 大量上调,AGPs 通过高度多孔的果胶网络参与了细胞扩张,AGPs含量和细胞扩张速率可能存在相关性。不仅如此,低温可能导致“生理性干旱”胁迫的产生[34],并可能影响AGPs 在植物体中的分布[30]。Leszczuk 等[4]在低温对雏菊(Bellis perennis)胚珠和花药发育过程中AGPs 分布变化的影响研究中,利用识别AGPs 的特异性抗体揭示了低温胁迫条件下AGPs 在雏菊雌性和雄性生殖结构中的分布发生了显著变化,AGPs 在胚囊壁中完全消失,AGPs定位于小孢子的公共壁和成熟花粉粒,以及在残余的绒毡层细胞中积累;Yan 等[35]采用免疫荧光标记技术研究了人工低温胁迫下多种AGPs 在香蕉幼苗叶片中的分布及含量变化规律;王莹莹[28]发现,JIM8等抗体所识别的AGPs 抗原在香蕉叶表皮、保卫细胞均有分布,既具有保护作用,又抵御低温胁迫。可见,低温等非生物胁迫影响AGPs 在植物体中的分布,AGPs 在植物应对低温、盐碱胁迫导致的生理干旱等非生物胁迫过程中发挥了重要作用。
灵武长枣是干旱盐碱地和沙荒地种植的先锋树种,叶表皮具有较厚的角质层,分布了大量的AGPs,这减少了植物体内水分的过量蒸腾,有助于叶片的光合作用。叶高度发达的栅栏组织维持了较高的光合作用能力。研究表明,低温胁迫下,香蕉叶片组织细胞内的可溶性总糖、还原糖、甘油及可溶性蛋白质的含量均随着温度的降低而升高,并且淀粉与糖之间的转化与植物的抗冷性存在密切关系[36];烟草AGPs 从引导组织向花粉管转移,为花粉管的生长提供营养支持[37]。本试验表明,灵武长枣4个时期叶栅栏组织细胞的细胞壁和细胞内部都分布着大量JIM8抗体所识别的抗原,基本没有明显的变化,叶肉始终是叶AGPs 分布的主要部位,并且后期随着叶片的发育,栅栏组织排列更加紧密,AGPs分布有所增强。本试验还发现,由于灵武长枣发育早期花后30 d 气温相对较低,蒸腾速率较小,水分条件相对较好,较高的气孔导度和胞间CO2浓度保证了较高的净光合速率,满足了早期果实对营养物质较高的需求;而花后60 d 气温高,蒸腾速率增大,其他光合指标及可溶性糖质量比均下降;花后30~60 d,蔗糖质量比逐渐下降至最低,这可能与叶片分解类酶活性的增长较高有关,促进了叶片中蔗糖的降解;在花后90 d,气孔导度和胞间CO2浓度再次增高,净光合速率明显提高,叶可溶性糖尤其是蔗糖质量比也大幅增长,这更有利于蔗糖从源叶向果实库的运输,以适应干旱环境下果实发育后期对营养的大量需求。叶肉栅栏组织细胞是植物进行光合作用的主要场所,光合作用受外界生态因子和内部因素的综合影响,栅栏组织细胞中大量的AGPs可能既参与了叶营养物质的合成,又作为营养物质来源,为干旱条件下植物的生长发育和后期果实营养物质的积累提供了保障和营养支持。而主脉和侧脉维管束木质部和韧皮部细胞的细胞壁和细胞内部分布的AGPs,为干旱条件下水分和养分的高效运输提供了保障[2,9]。因此,干旱条件下叶结构的适应性与各部位AGPs 的分布有关,AGPs 被认为有助于植物应对生物和非生物胁迫的反应[38]。
