小球藻多糖的结构与生物活性研究进展
2024-01-03何启煜安子哲陈安进王寒梅李方圆卢海燕
何启煜,安子哲,陈安进,王寒梅,李方圆,卢海燕,赵 雪,
(1.中国海洋大学食品科学与工程学院,海洋食品加工与安全控制全国重点实验室,山东 青岛 266003;2.青岛市市立医院,山东 青岛 266011)
小球藻(Chlorella)属于绿藻门(Chlorophyta)绿球藻目(Chlorococcales)小球藻科(Chlorellaceae)小球藻属(Chlorella),生长在淡水或海水中,按养殖方式可分为自养型、异养型和复合培养型[1]。小球藻是第一个被人成功分离和培育的单细胞绿藻,其体积微小、繁殖速度快[2],目前主要的养殖品种有普通小球藻(C.vulgaris)、蛋白核小球藻(C.pyrenoidosa)、椭圆形小绿藻(C.ellipsoidea)、柱头小球藻(C.stigmatophora)、索氏小球藻(C.sorokiniana)、佐氏小球藻(C.zofingiensis)和单细胞油脂高脂突变株小球藻(C.kessleri),养殖种类繁多。小球藻中富含蛋白质、脂类、多糖、小球藻生长因子、维生素、天然色素和矿物质等物质,已被联合国粮农组织(Food and Agriculture Organization of the United Nations,FAO)认证为一种绿色健康的食品[3],并广泛应用于水产养殖、化妆品、医药与环境保护等领域[4],具有广阔的开发前景。
小球藻中的多糖质量分数为10%~25%[5],是小球藻中一种重要的活性物质。研究发现,小球藻多糖具有抗氧化[6]、降血脂[7]、抗哮喘[8]、抗肿瘤[9]、调节免疫[10]和保护神经[11]等生物活性。小球藻多糖分为胞外多糖、胞壁多糖和细胞基质多糖。按单糖组成将小球藻多糖分为六大类:以葡萄糖(glucose,Glc)为主的杂多糖、以半乳糖(galactose,Gal)为主的杂多糖、以鼠李糖(rhamnose,Rha)为主的杂多糖、以阿拉伯糖(arabinose,Ara)为主的杂多糖、以葡萄糖醛酸(glucuronic acid,GlcUA)为主的杂多糖和以甘露糖(mannose,Man)为主的杂多糖。这些多糖中同明含Glc、Gal、Rha、Ara、Man、木糖(xylose,Xyl)和GlcUA等多种单糖,而且糖链上还具有Gal、Rha或GlcUA等多种支链,部分糖残基上还存在甲基化、硫酸根和乙酰化修饰,因此小球藻多糖的结构非常复杂,这给小球藻多糖相关产品的分析检测和质量控制带来了巨大的挑战。
目前,微藻多糖的分离纯化方法主要有水提取、分级醇沉、阴离子交换色谱法[10,12-13]和凝胶排阻色谱法[14]。多糖结构的分析方法主要有甲基化结合气相色谱-质谱联用(gas chormatography-mass spectrometry,GC-MS)法[8,15]和二维核磁共振(two-dimensional nuclear magnetic resonance,2D-NMR)法[7,16]。
1 小球藻多糖的组成与结构
1.1 葡聚糖和以Glc为主的杂多糖
小球藻中含有较高质量分数的葡聚糖或以Glc为主的杂多糖,其中Glc质量分数可达到50%~70%,Gal 质量分数1 5%~2 5%,同明还含有少量Rha、Xyl、Ara和Man。2008年,Suárez等[17]从蛋白核小球藻(C.pyrenoidosa)中分离纯化出一种β-1,2-葡聚糖,其线性和环状结构的物质的量比为64∶36。环状葡聚糖含有18~35 个Glc残基,其中70%由21~22 个Glc组成。而50%线性的葡聚糖是由20~21 个Glc组成,结构如图1 A 所示。2015 年,Tabarsa 等[10]利用水提取和弱阴离子交换色谱法从普通小球藻(C.vulgaris)中分离纯化出一种分子质量为1.97×103Da的α-1,6葡聚糖。经NMR分析表明有一部分Glc的C-3位上连接有Glc支链。其结构如图1B所示。
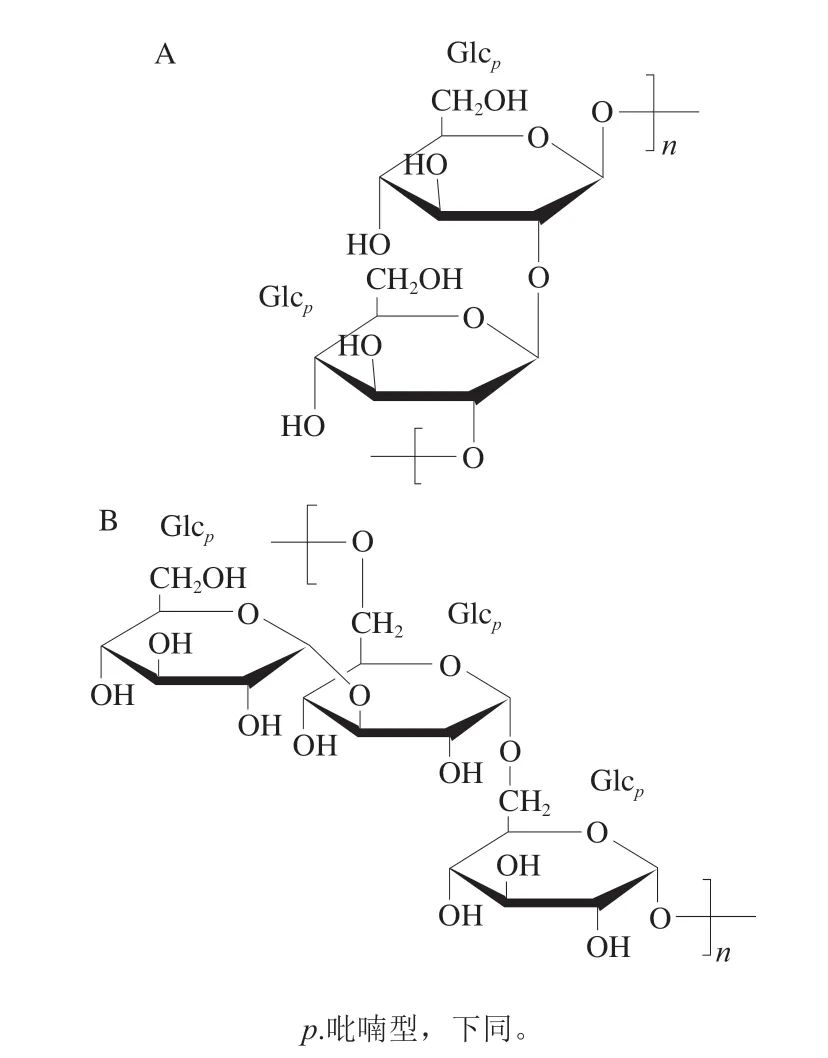
图1 蛋白核小球藻中的β-1,2-葡聚糖(A)和普通小球藻中的α-1,6-葡聚糖(B)的结构[10]Fig.1 Structures of β-1,2-glucan from C. pyrenoidosa (A) and α-1,6-glucan from C. vulgaris (B)[10]
2017年,Qi Jia等[12]利用热水提取、弱阴离子交换色谱法从椭圆形小球藻(C.ellipsoidea)中分离纯化获得一种分子质量为2.37×105Da的葡聚杂多糖,其单糖组成为Glc(58.80%,物质的量百分比,下同)、Gal(27.00%)、Rha(11.60%)和Man(2.60%),同明含有6.00% GlcUA和6.00%硫酸根。甲基化与GC-MS结果表明,其主链由1,4连接的Glc与1,6连接的Gal组成,并在Glc的C-3、Gal的C-4位上存在支链。
1.2 半乳聚糖和以Gal为主的杂多糖
2006年,Suárez等[18]用水提取、强阴离子交换法和凝胶色谱法从蛋白核小球藻(C.pyrenoidosa)中获得一种分子质量为1.50×104Da的呋喃型半乳聚糖。如图2A所示,2D-NMR结果表明,其主要由→2-α-DGalf-1→构成。2010年,Suárez等[19]从蛋白核小球藻(C.pyrenoidosa)中获得一种磷酸化的以Gal为主的杂多糖,同明含有少量Glc和Man。如图2B所示,其糖链是由→3-α-D-Gal-1→组成,在Gal的C-6位上存在支链,其中50%的支链为β-D-Glc,50%为α-D-Man-1-磷酸和3-O-Meα-Man-1-磷酸。同明发现,大约有35%的Gal的C-2与C-4处连有乙酰基修饰。
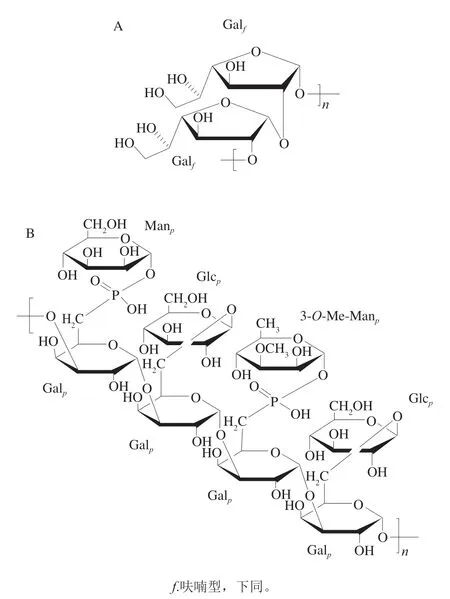
图2 蛋白核小球藻中的α-1,2-半乳聚糖(A)和以Gal为主的杂多糖(B)的结构[19]Fig.2 Structures of α-1,2-galactan (A) and a heteropolysaccharide composed mainly of galactose (B) from C. pyrenoidosa[19]
2007年,Řezanka等[20]从单细胞油脂高脂突变株小球藻(C.kessleri)中提取水溶性多糖,经葡萄糖酶法除去葡聚糖后,用甲基化与GC-MS分析确定,其主链由→4-L-Rhap-1,4-D-Galp-1,4-D-Galp-1,4-D-Galp-1,3-L-Arap-1→构成,并在主链中的部分Gal残基C-3位上存在支链,支链有Galf和Xylp两种类型。结构如图3A所示。
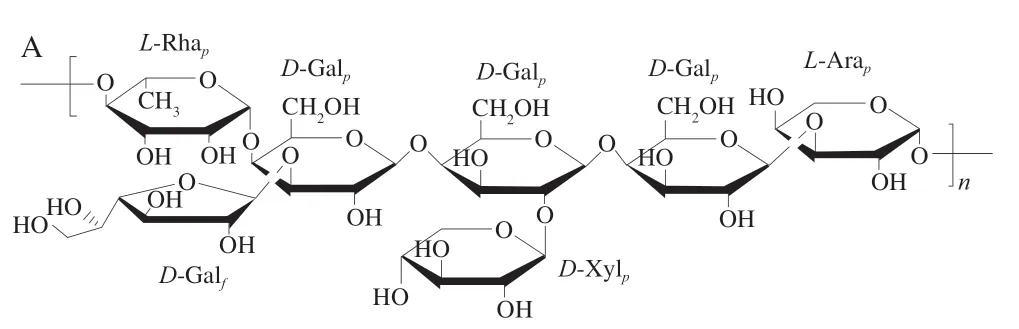
图3 单细胞油脂高脂突变株小球藻中以Gal为主的杂多糖(A、B)和鼠李聚糖(C)的结构[20-21]Fig.3 Structures of a heteropolysaccharide composed mainly of galactose (A and B) and rhamnoglycan (C) from C. kessleri[20-21]
2020年,经热水提取、弱阴离子交换色谱和凝胶色谱分离,Sushytskyi等[21]从单细胞油脂高脂突变株小球藻(C.kessleri)中获得3 个多糖组分,均主要由Gal、Rha、Xyl和4-O-Me-Xyl组成。经甲基化和2D-NMR分析表明,其中一个组分由1,3-连接的Galp构成主链,并在其C-2位上连有Xyl支链,且部分Xyl具有甲基化修饰,结构如图3B所示;还有一个组分为多样连接方式的鼠李聚糖结构,其由1,3-连接的Rha构成主链,并在C-2位上连有1,2-连接的Rha支链,结构如图3C所示。
2019 年,Barboríková 等[8]从普通小球藻(C.vulgaris)的培养液中获得一种分子质量为8.40×104Da胞外多糖,单糖组成为Gal(35.62%)、Ara(32.31%)、Rha(15.75%)、Man(7.24%)、Glc(5.55%)、Xyl(3.53%)。经甲基化和GC-MS分析确定,该多糖中存在1,2,3-连接的Rha、1,3-连接的Rha、1,2-连接的Ara、1,4,6-连接的Gal以及末端的Galf。此外,还发现了部分甲基化的己糖(如3-O-Me-Glc、3-O-Me-Gal)、2-O-Me-戊糖和3-O-Me-戊糖的存在。
2020年,Ferreira等[15]从普通小球藻(C.vulgaris)的培养基中通过水提和醇沉获得一种胞外多糖,其单糖组成为Gal(52.80%)、GlcUA(18.00%)、Rha(8.00%)、Man(7.70%)、Ara(6.40%)、Xyl(5.50%)和Glc(1.60%)。经甲基化和GC-MS分析发现其1,6-连接的Gal的含量最高,且还含有末端连接的呋喃型Gal,此外还存在1,3-连接与1,3,6-连接的Gal,以及少量的1,3-连接的Rha、1,2-连接的Rha和1,2-连接的Man。
2020年,Capek等[22]从普通小球藻(C.vulgaris)的培养液中分离纯化出一种胞外多糖,其单糖组成为Gal(36.00%)、Rha(29.20%)、Ara(19.00%)、Glc(8.40%)、Man(4.40%)、Xyl(3.00%)。经甲基化与2D-NMR分析,该胞外多糖存在1 个由重复单元→2-α-L-Rha-1→3-α-L-Rha-1→序列形成的主链结构,推测部分Rha的C-2、C-3或C-4位连接有聚合度不均一的支链结构,其支链主要由α-1,6连接的Gal或α-1,2连接的Ara构成,同明这些支链本身也连接有小的支链,结构如图4所示。分析结果还表明,多糖中存在大量的末端Galf连接方式,说明这些Gal可能是复杂支链结构的末端单位。在2D-NMR分析中,还鉴定出部分Gal与Ara的C-3位具有甲基化修饰。综合上述分析,该多糖具有高度分支的α-L-Ara-α-L-Rha-α,β-D-Gal结构。
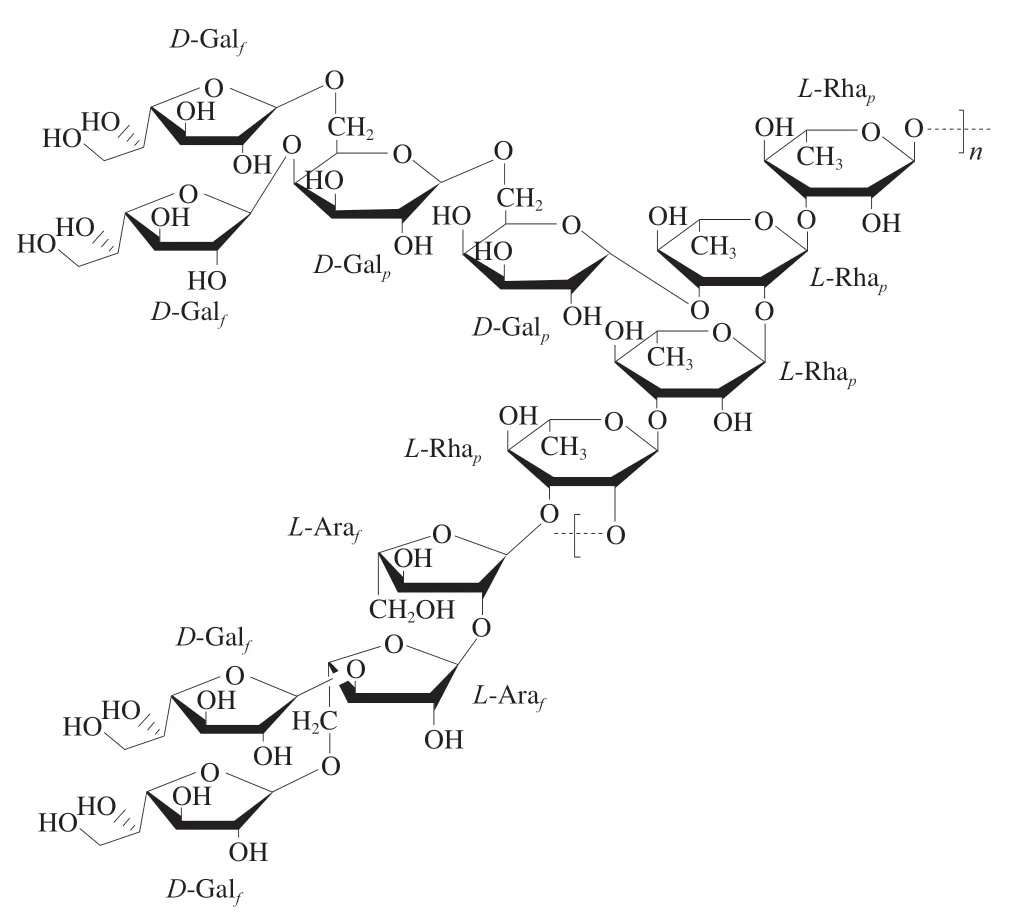
图4 普通小球藻中以Gal为主的杂多糖的结构[22]Fig.4 Structure of a heteropolysaccharide composed mainly of galactose from C. vulgaris[22]
通过比较Barboríková[8]、Ferreira[15]和Capek[22]等通过甲基化和GC-MS研究发现,小球藻多糖中均具有较高比例的1,4,6-连接的Gal、1,3-连接的Rha、1,2,3-连接的Rha、1,2-连接的Ara和末端的呋喃型Gal。推测以上连接方式是从普通小球藻(C.vulgaris)的培养液中获得胞外多糖的重要连接单元。
1.3 以Rha为主的杂多糖
小球藻中还具有独特的鼠李聚糖。1999年,Ogawa等[23]利用阴离子交换色谱从普通小球藻(C.vulgaris)中纯化出一种葡萄糖醛酸-鼠李聚糖。其主要由Rha(57.80%)和GlcUA(14.20%)组成,还含有Ara、Xyl、Man、Gal、2-O-Me-Rha、3-O-Me-Rha。酸降解产物经分离纯化和2D-NMR分析,确定了其-GlcUAp-α-1→3-L-Rhap-α-1→2-L-Rhap三糖结构,结构如图5A所示。2021年,Wan Xuzhi等[16]利用弱阴离子交换色谱和凝胶排阻色谱,从蛋白核小球藻(C.pyrenoidosa)中分离后获得1 个分子质量为1.58×104Da的酸性多糖组分,主要由Rha(34.7%)、Gal(26.0%)、Ara(15.5%)、Man(11.9%)、Glc(8.8%)、GlcUA(2.3%)、半乳糖醛酸(galacturonic acid,GalUA)(0.8%)组成。经2D-NMR分析确定,其具有1,2-连接的Rha、1,4-连接的GalUA、1,3-连接的Man、1,6-连接的Glc、1,3,6-连接的Gal和末端连接的Ara与GlcUA,结构如图5B所示。

图5 普通小球藻和蛋白核小球藻中三糖(A)和以Rha为主的杂多糖(B)的结构[16,23]Fig.5 Structures of trisaccharide (A) and a heteropolysaccharide composed mainly of rhamnose (B) from C. vulgaris and C. pyrenoidosa[16,23]
1.4 以Ara为主的杂多糖
2001年,Pugh等[24]从蛋白核小球藻(C.pyrenoidosa)中提取分离纯化出一种以Ara为主的杂多糖,由Ara(40.00%)、Gal(33.75%)和Rha(15.00%)和其他微量单糖组分组成。2005年,Suárez等[14]从蛋白核小球藻(C.pyrenoidosa)中获得分子质量为4.7×105Da的半乳-阿拉伯聚糖,Ara和Gal的物质的量比为2∶1。经甲基化、GC-MS与2D-NMR分析发现其主链由2-α-L-Ara-1,3-β-DGal-1重复单元构成,并在Gal的C-4位连有L-Ara分支,如图6A所示。
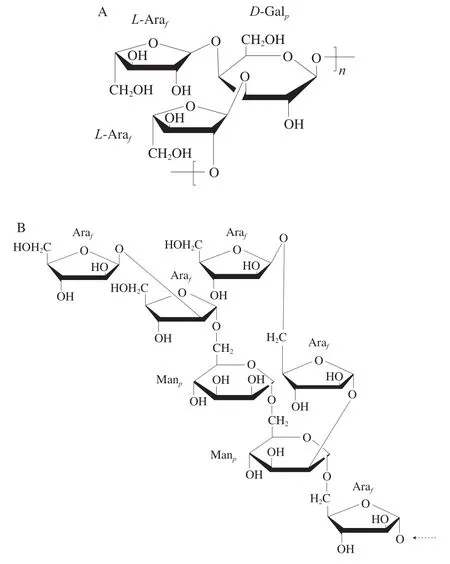
图6 蛋白核小球藻中半乳-阿拉伯聚糖(A)和普通小球藻中甘露-阿拉伯聚糖(B)的结构[14,25]Fig.6 Structures of arabinogalactan (A) and mannan-arabinose (B)from C. pyrenoidosa and C. vulgaris[14,25]
2012年,Pieper等[25]采用碱提和凝胶排阻色谱法从普通小球藻(C.vulgaris)的细胞壁中获得1 种分子质量为8×103Da的甘露-阿拉伯聚糖,Ara与Man物质的量比为39∶16。经电喷雾质谱法与2D-NMR分析确定,其主链的平均单元为2-D-Manp-α-1,6-D-Manp-α-1,5-D-Araf-α-1。在Manp的C-2和C-6位上存在D-Araf-β-1,2-D-Araf-1-和D-Araf-β-1,5-D-Araf-1-的二糖支链,平均结构如图6B所示。
1.5 以Man为主的杂多糖
2019 年,ZhangJianzhi 等[9]从佐氏小球藻(C.zofingiensis)获得一种分子质量为2.66×104Da的胞内多糖,主要由Man(44%)、Gal(31%)和Glc(17%)组成。2020年,Wan Xuzhi等[7]从蛋白核小球藻(C.pyrenoidosa)中获得一种分子质量为5.63×106Da的复杂多糖,其各单糖组成为Man(25.30%)、Rha(23.30%)、Glc(19.30%)、岩藻糖(fucose,Fuc)(17.50%)、Xyl(8.30%)、Ara(6.30%),同明含有5.5%的GlcUA。2D-NMR分析确定复杂糖链的主要骨架为4-α-L-Rhap-1,2-α-L-Fucp-1,3-β-D-Manp-1,4-β-D-Xylp-1,4-α-L-Rhap-1,并在Manp的C-6位连有α-D-Glcp-1,3-β-DGlcUAp-1,4-β-D-Arap-1-支链。结构如图7所示。

图7 蛋白核小球藻(C. pyrenoidosa)中杂多糖的结构[7]Fig.7 Structure of heteropolysaccharide from C. pyrenoidosa[7]
1.6 以GlcUA为主的杂多糖
1998年,Ogawa等[13]采用阴离子交换色谱从普通小球藻(C.vulgaris)中分离纯化得到1 个酸性多糖,其主要含有GlcUA,单糖组成为GlcUA(42.00%)、Rha(17.40%)、Xyl(16.80%)、Man(3.00%)、Glc(1.90%)、Gal(18.90%)。经三氟乙酸降解和2D-NMR分析,获得其二糖的结构为α-D-GlcUAp-1,3-α-L-Rhap,如图8所示。不同种类小球藻多糖的组成和结构比较如表1所示。
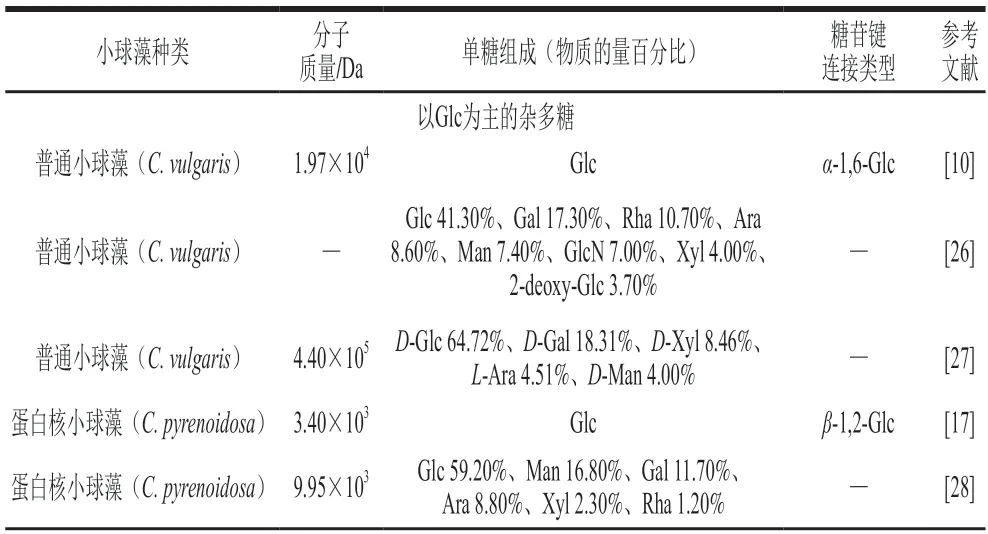
表1 不同种类小球藻多糖的化学组成与结构的比较Table 1 Comparison of chemical composition and structure among polysaccharides from different species of Chlorella
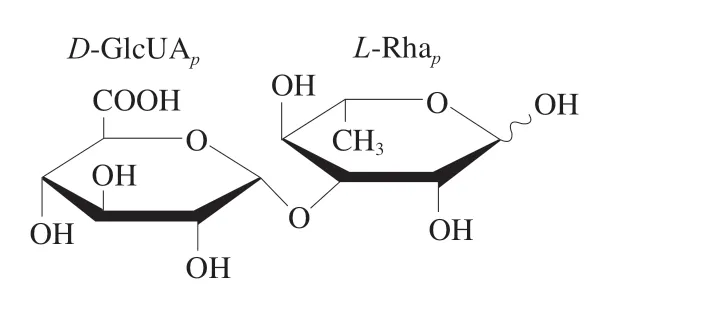
图8 普通小球藻(C. vulgaris)中糖醛酸-鼠李二糖的结构[13]Fig.8 Structure of glucuronic acid-rhanose disaccharide from C. vulgaris[13]
2 小球藻多糖的生物活性及其构效关系
2.1 小球藻多糖的免疫调节活性及其构效关系
目前多项研究表明,小球藻多糖具有良好的免疫调节活性。Yang Fangmei等[39]发现小鼠口服1~2 g/kgmb从蛋白核小球藻(C.pyrenoidosa)中提取的甘露杂多糖后,其巨噬细胞的吞噬率和吞噬指数分别提高了53%和73%。王凌等[40]通过体外细胞实验发现50 μg/mL小球藻多糖可显著促进小鼠腹腔巨噬细胞的增殖,使巨噬细胞的吞噬率提高了1.86 倍,是阳性对照药物脂多糖的1.2 倍。100 μg/mL的小球藻多糖使小鼠腹腔巨噬细胞产生NO的能力提高了1.96 倍,但效果稍弱于脂多糖。Suárez等[17]从蛋白核小球藻(C.pyrenoidosa)中提取分离获得了β-1,2连接的葡聚糖。在体外细胞实验中,15 μg/mL的葡聚糖诱导巨噬细胞产生NO的能力比脂多糖更强,这表明小球藻中的葡聚糖和以Man为主的杂多糖都有很好的提高细胞免疫的活性。
小球藻多糖同明对免疫因子的表达有促进作用。Tabarsa等[10]发现普通小球藻(C.vulgaris)中的α-1,6-连接的葡聚糖能够促进巨噬细胞中一氧化氮合成酶的表达,从而促进NO的产生。Hsu等[41]发现蛋白核小球藻(C.pyrenoidosa)中的鼠李杂多糖通过与I型跨膜蛋白-Toll样受体4的结合,激活了丝裂原活化蛋白激酶通路中的细胞外调节蛋白激酶、应激活化蛋白激酶和P-38蛋白的磷酸化,从而促进了白细胞介素-1β(interleukin-1β,IL-1β)前体mRNA的表达,最终促进IL-1β的表达,反应通路如图9所示。Qi Jia等[42]发现椭球型小球藻(C.ellipsoidea)粗多糖除了能促进炎症因子IL-1β和IL-6的表达外,还能促进抗炎因子IL-10和IL-12的表达,因此能够抑制促炎因子的过度活化。
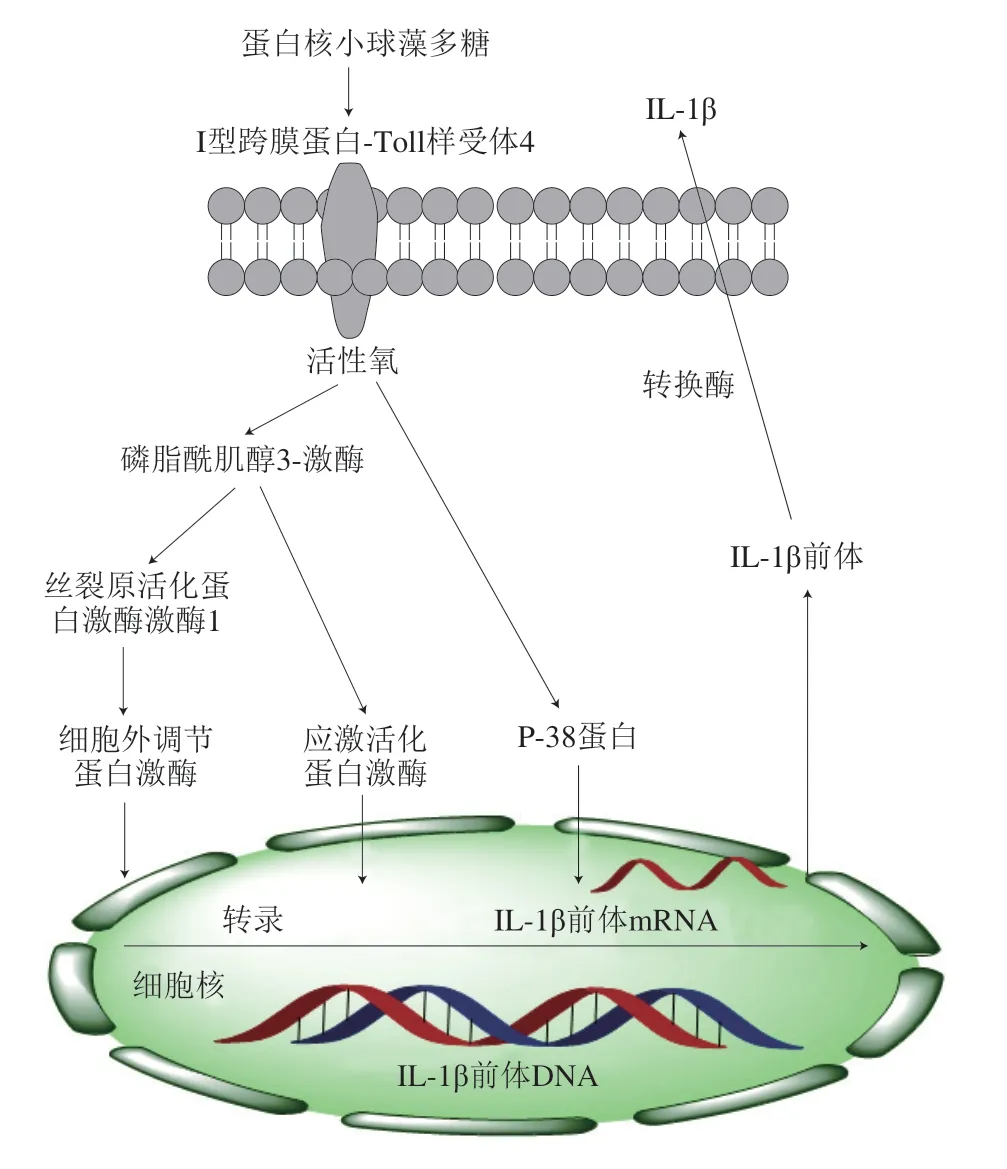
图9 蛋白核小球藻多糖(C. pyrenoidosa)参与的调控IL-1β表达的信号通路[41]Fig.9 C. pyrenoidosa-mediated signal transduction pathways in the regulation of interleukin-1β expression[41]
研究表明,小球藻多糖的免疫调节活性差异可能与其结构有关,单糖种类、分子质量和硫酸根含量均会影响其活性。Qi Jia等[42]研究了椭球型小球藻(C.ellipsoidea)粗多糖,发现分子质量是调节免疫功能的关键因素,在51~237 kDa范围内,椭球型小球藻(C.ellipsoidea)多糖的分子质量越大,其促进巨噬细胞产生NO的能力就越强。这一特性与Suárez[18]和Kralovec[43]等的研究结果相同。Kralovec等[43]研究发现,多糖与蛋白质组成的糖蛋白复合物表现出更强的促进B细胞增殖的作用。但是Qi Jia等[42]的研究结果表明,脱蛋白处理不会影响椭球型小球藻(C.ellipsoidea)粗多糖的免疫调节活性。因此小球藻多糖复合物的构效关系还有待于进一步研究。
2.2 小球藻多糖的抗氧化活性及其构效关系
研究报道小球藻多糖具有很好的抗氧化活性,Zhang Jianzhi[9]和吴思伟[44]等利用体外自由基模型,发现索氏小球藻(C.sorokiniana)中的粗多糖、佐氏小球藻(C.zofingiensis)中以Man为主的杂多糖和普通小球藻(C.vulgaris)中以Gal为主的杂多糖均具有清除羟自由基和1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-trinitrophenylhydrazine,DPPH)自由基的活性,且活性呈剂量依赖性。比较发现,以Gal为主的杂多糖清除羟自由基活性高于以Man为主的杂多糖和粗多糖,但对DPPH自由基的清除活性却相反。Song Hong等[45]发现5 mg/mL北极小球藻(Arctic Chlorellasp.)中的半乳杂多糖对DPPH自由基和羟自由基的清除率大于60%,但是对超氧自由基清除率只有42.2%。
研究表明化学结构对小球藻多糖的抗氧化性有很大的影响。Mohamed等[46]发现,普通小球藻(C.vulgaris)粗多糖的硫酸根含量越高,其抗氧化活性越强。而杨海燕等[47]发现,普通小球藻(C.vulgaris)粗多糖的糖醛酸含量越高,其清除自由基能力越强。此外,不同提取方式获得的小球藻多糖的结构不同,其抗氧化活性也不同。Chen Yixuan等[6]采用醇沉法从蛋白核小球藻(C.pyrenoidosa)多糖中获得了以Glc为主的杂多糖,1.2 mg/mL杂多糖对羟自由基的清除率高达92.71%。张喜峰等[48]采用醇沉法从小球藻中获得了以Gal为主的杂多糖,其清除自由基的活性较其他组分更高,分析原因是其分子质量较低,糖醛酸含量更高。Yu Mengen等[49]利用冻融、微波、超声波、碱、热水和纤维素酶法等不同的提取方法获得了不同结构的小球藻多糖,比较其抗氧化活性的差异发现,超声波法提取的多糖具有更高的硫酸根含量和糖醛酸含量,因此具有最高的清除DPPH自由基、超氧化物、羟自由基活性和金属螯合能力。刘芬等[50]的研究也发现,超声铺助热水浸提法获得的小球藻粗多糖的抗氧化活性高于酶铺助提取法,获得的多糖对羟自由基的最大清除率高达95%。刘凤路等[51]发现螺旋藻多糖和小球藻多糖复合后,表现出更强的抗氧化性。
抗衰老活性与抗氧化活性密不可分,Chen Yixuan[28]和Gui Jiangli[26]等分别发现,蛋白核小球藻(C.pyrenoidosa)和普通小球藻(C.vulgaris)中的以Glc为主的杂多糖可以显著提高黑腹果蝇和秀丽隐杆线虫体内的超氧化物歧化酶、过氧化氢酶和谷胱甘肽过氧化物酶的活性,最终延缓黑腹果蝇和秀丽隐杆线虫的衰老,延长它们的寿命。
2.3 小球藻多糖的抗炎活性
Guzmán 等[52]研究发 现柱头 小球藻(C.stigmatophora)多糖具有抗炎活性,其抑制角叉菜诱导的大鼠肿胀效果优于消炎药默克。Barboríková[8]和Capek[22]等发现,在豚鼠实验性诱导的过敏性气道炎症模型中,普通小球藻(C.vulgaris)中的半乳聚糖具有显著的支气管扩张、抗炎和止咳作用,效果与常用的抗过敏药物沙美特罗和布地奈德相当,表明小球藻半乳聚糖具有治疗哮喘和慢性阻塞性肺疾病的潜质。Chen等[11]发现阿尔茨海默病小鼠口服从蛋白核小球藻(C.pyrenoidosa)中提取的葡聚杂多糖后,其多巴胺分泌量增加,神经元得到了很好的保护。他还发现此葡聚杂多糖能抑制阿尔茨海默病小鼠体内促炎因子(肿瘤坏死因子-α、IL-1β和IL-6)的表达水平,同明还能抑制小胶质细胞的活化。由于过度激活的小胶质细胞会诱发炎症,并导致神经元的损伤,因此他推测蛋白核小球藻(C.pyrenoidosa)葡聚杂多糖的神经保护作用与其抗炎活性有关。这一特点是常用的阿尔茨海默病治疗药物——左二羟基苯丙氨酸所不具备的,长期使用左二羟基苯丙氨酸可能会导致慢性炎症反应。以此体现了使用蛋白核小球藻(C.pyrenoidosa)中的葡聚杂多糖明安全性好的优点。
2.4 小球藻多糖的抗癌活性
研究发现,小球藻多糖具有很好的抗肿瘤活性。Zhang Jianzhi 等[9,53]研究表明佐氏小球藻(C.zofingiensis)中以Man为主的杂多糖、普通小球藻(C.vulgaris)和蛋白核小球藻(C.pyrenoidosa)中以Gal为主的杂多糖,在体外均能够显著抑制HCT8回盲肠癌细胞的增殖。以Man为主的杂多糖和Gal为主的杂多糖对HCT8的半抑制质量浓度分别为1.70 mg/mL和3.14 mg/mL。Sheng Jianchun等[34]从蛋白核小球藻(C.pyrenoidosa)中分离获得以Gal为主的杂多糖CPPS Ia和以Rha为主的杂多糖CPPS IIa,在1 mg/mL的质量浓度下,这两个组分在体外对腺癌人类肺泡基底上皮细胞A549的生长抑制率分别为68.7%和49.5%,其中CPPS Ia的抑制效果与阳性对照药物5-氟尿嘧啶相近。谭成玉等[54]发现,海洋小球藻粗多糖在体外能抑制人乳腺癌细胞MCF-7对人膀胱癌细胞ECV-304的迁移,表明小球藻多糖对肿瘤血管生成具有抑制作用。徐韬钧等[55]发现小球藻多糖能够下调肝癌细胞SMMC-7721的B淋巴细胞瘤-2基因,同明上调凋亡执行蛋白Caspsae-3,通过激活细胞的凋亡途径而抑制肝癌细胞SMMC-7721的增殖。
2.5 小球藻多糖的抗菌活性
天然多糖的抗菌活性使得多糖在农业防菌涂料和医疗领域抑菌敷料方面具有巨大潜力。孙建瑞等[56]发现普通小球藻(C.vulgaris)多糖对金黄色葡萄球菌、白色葡萄球菌、绿脓杆菌和大肠杆菌具有抑菌作用,对革兰氏阳性菌的抑制作用要高于革兰氏阴性菌,但是对枯草芽孢杆菌、沙门氏菌、黑曲霉和中华根霉等真菌没有抑菌作用。然而刘凤路等[51]的研究结果完全相反,其发现小球藻与螺旋藻的复合多糖对革兰氏阴性细菌和真菌有较强的抑制作用,而对革兰氏阳性细菌抑制效果不佳。刘四光等[57]从自养的小球藻中分离出PCA2-1(硫酸根8.46%)和PCA2-2(硫酸根0.77%)两个粗多糖组分,比较发现PCA2-1的抑菌效果不佳,反而会促进细菌的生长;而PCA2-2对溶壁微球菌和解藻朊酸弧菌有较强的抗菌活性,抗菌率可达70%以上,因此认为硫酸根对其抗菌活性有显著的影响。
2.6 小球藻多糖对肠道菌群的调节活性
Wan Xuzhi[7]、Beheshtipour[58]和Lv Kun[59]等的研究结果表明,小球藻多糖可以作为一种益生元促进肠道菌群健康。Lv Kun等[59]发现蛋白核小球藻(C.pyrenoidosa)中以Gal为主的杂多糖不会被唾液和胃肠道降解,提高了青春双歧杆菌和普氏粪杆菌的丰度,被肠道菌群利用后产生了大量有益于肠道健康的短链脂肪酸。Wan Xuzhi等[7]研究表明,蛋白核小球藻(C.pyrenoidosa)中的甘露-鼠李杂多糖可以改善高脂血症大鼠的脂质代谢紊乱和肠道菌群。甘露-鼠李杂多糖通过激活磷酸腺苷活化蛋白激酶,抑制羟甲基戊二酰铺酶A还原酶的磷酸化,从而限制胆固醇的合成。甘露-鼠李杂多糖使得盲肠中粪球菌属、乳酸杆菌属和苏黎世杆菌属丰度增加,同明使瘤胃球菌属丰度减少。通过相关性分析发现,在甘露鼠李杂多糖的作用下,肠道菌群通过调节总胆汁酸和短链脂肪酸代谢起到降血脂的效果。Wan Xuzhi等[16]还发现蛋白核小球藻(C.pyrenoidosa)中的鼠李杂多糖能够调节秀丽隐杆线虫的肠道菌群,从而提高其抗氧化活性。
3 结语
综上所述,就结构组成方面而言,众多研究表明小球藻多糖多是以Gal为主的杂多糖,其次是以Glc为主的杂多糖和以Rha为主的杂多糖。说明这3 种单糖是小球藻多糖的重要组成部分,其中Gal多以1,3-连接和1,6-连接形式存在,Glc多以1,6-连接形式存在,Rha多以1,2-连接和1,3-连接形式存在。从戊糖和脱氧己糖含量来看,小球藻多糖中Ara含量一般大于Xyl含量,而Rha含量一般大于岩藻糖含量。从研究手段来看,目前,关于小球藻多糖结构的研究方法主要有传统化学方法和2D-NMR,但其只能获得部分寡糖的连接顺序和多糖的平均结构单元,无法确定多糖链的完整序列。而最新研究表明,液相色谱-质谱联用技术在表征天然寡糖全指纹图谱、解析支链寡糖结构和多糖连接方式上取得了较大进展[60-61],将其应用于小球藻多糖结构分析领域,可以弥补部分主流手段的不足,但质谱技术也存在着无法辨别同分异构体的局限性。因此,为了进一步探究小球藻多糖的结构,多手段联合使用的分析策略是不可或缺的。就活性方面而言,小球藻多糖展现出了多样的活性功能,同明具有低毒性,表明小球藻多糖具备开发功能性食品和药物的巨大潜力。但是对小球藻多糖的生物活性仍需要更多体内实验和临床效果验证,需要更加明确的作用机理与作用途径。
除了结构与活性方面研究的不足外,通过对该领域的总结分析还发现,目前鲜有同明包含两个方向的研究,部分活性方向的研究甚至缺乏对小球藻多糖的基本结构信息的解析。同明,关于构效关系研究也仅停留在较为宏观的因素上,如分子质量、硫酸根含量和单糖组成等,缺乏和分子结构层面的关联性研究。由此导致了目前该领域在结构与活性两个方向之间存在着强烈的割裂感。因此,为了更好地阐明小球藻多糖的结构与活性之间的关系,研究者们首先仍需要对小球藻多糖的结构做进一步探索与研究,随后在此基础上,对活性机理的研究也应更加深入至分子结构层面。加强两个研究方向的联系是今后推动该领域进一步发展的重点。
从产业需求来看,自2010年以来,我国小球藻行业运行态势良好,市场需求量和产量逐年增长,2022年预计产量为4 005 t。全球市场规模将从2019年的2.061亿 美元增至2025年的2.76亿 美元。由此可见,小球藻产业正在蓬勃发展。除此之外,其他微藻也越来越受到人们的关注,如雨生红球藻和杜氏盐藻,以及拥有更完善商业化模式的螺旋藻,它们同样具有活性天然多糖。对小球藻多糖精细结构和构效关系的研究,为其他复杂的微藻多糖的研究提供了很好的研究思路和技术方法。从天然多糖结构研究层面上来看,小球藻多糖作为一个复杂的天然多糖的典型案例,其精细结构的研究思路和策略同样可以运用于其他复杂的天然多糖。总地来说,小球藻多糖结构和活性的研究推动的将不仅仅是小球藻产业,还将带动其他微藻产业的发展,进而有助于天然多糖在食品保健、化妆保养和生物医疗等领域的发展。
