典型城乡绿化树种土壤微生物组成和多样性的环境响应差异
2023-12-26王泽锦
冉 堃,王泽锦,李 品
森林培育与保护教育部重点实验室,干旱半干旱地区森林培育及生态系统研究国家林草局重点实验室,城市林业研究中心,北京林业大学,北京 100083
改革开放以来的城市化带来了经济上的蓬勃发展[1],并引起了人口流动[2],二者在环境变化、人口再分布以及区域发展的影响不容忽视。一方面,在城乡梯度的水平方向上形成了环境差异;另一方面,如“热岛效应”“土壤污染”“大气污染”等环境负荷问题也随之而来,此类问题会导致环境条件退化、生物多样性减少等严重的城市生态问题[3]。城市森林作为城市生态系统中重要的组成成分,在缓解环境问题上有着关键作用[4],而土壤环境作为城市森林生长的“摇篮”,通过研究土壤生态系统对环境变化的响应,能够探究出有益于城市森林生长发育的影响因素,进而为缓解城市化环境问题提供科学有效的理论依据。
土壤生态系统中微生物群落的转化速率是植物残体的10倍[5],同时其生态生理特征对土壤养分、温度以及湿度的响应速度超过其它常规指标,如有机碳、全氮、全磷等[6],因此普遍认为土壤微生物群落可以作为反映土壤环境变化的指标。土壤微生物群落在分类上可以分为原核土壤微生物与真核土壤微生物,前者包括细菌、古菌和放线菌等,而后者包括真菌、藻类和原生动物等[7]。在土壤原核微生物中,土壤细菌是丰富度、多样性最高的一个类群[8],它们结构简单且适应环境变化的能力较弱,因此一般通过改变自身的群体结构来适应新环境[9]。目前国内外研究发现土壤细菌极易受到环境因子的影响[10—14],但响应结果并不一致,以土壤pH值为例,Tian等[15]发现土壤pH与土壤细菌群落无显著相关关系,而周丽等[16]发现土壤pH显著影响了研究区域中的土壤细菌种类。此外,气候因子如温度、湿度等[17]和生长在土壤上的植物群落多样性以及植物种类[18]也会对土壤细菌群落产生显著影响。
在土壤真核微生物中,土壤真菌作为另一重要的类群,在土壤养分的分解、转化及循环等方面起着重要作用[19],同时还参与土壤有机质矿化过程[20]。相比土壤细菌群落,真菌群落对于环境因子的适应力较强,因此能够在一定程度上增强土壤的抗盐碱能力与抗逆性,在维持土壤生态系统健康方面有着重要作用[21],同时一些土壤真菌可以与树木根系形成良性共生关系,形成独特的群落结构与分布规律[22]。国内外对于土壤真菌群落与环境因子的响应主要集中在土壤因子方面。与细菌群落相似,大部分土壤理化性质因子也会对真菌群落产生影响,如王峰等[23]发现土壤pH、土壤有机质以及速效养分会对土壤真菌群落结构产生影响。但真菌群落在响应上与细菌群落存在差异,如Han等[11]发现土壤细菌群落多样性随土壤可降解有机碳含量增加而减少,但土壤真菌群落表现为随土壤可降解有机碳含量增加而增加。
最近有研究结合植物群落多样性、植物种类、气候因子以及土壤理化性质因子对土壤微生物群落进行了综合分析,发现这四类因子对微生物存在交叉影响。Schlatter等[24]发现,不仅植物与土壤微生物之间存在相互作用,微生物之间也存在如竞争、拮抗等关系,且微生物之间的相互关系可以抑制多种植物病害,在Liu等[25]的研究中也得出了相似的结论。对于气候因素,Sun等[14]认为目前过于注重气候因子对土壤细菌群落的影响而忽略了气候因子对其他环境因子的影响力;他们发现土壤细菌群落对于各类环境因子并不总是显著相关的,其中气温因子的解释率最低,仅占1.6%—18%,而土壤理化性质因子占4.2%—70.8%,植物因子占7.8%—20.6%。
城市化发展导致了城乡环境差异与生态环境问题。城乡环境差异表现在城乡梯度上环境因子和植物具有显著的差异。土壤环境是城市生态环境中重要的部分,其中土壤微生物会对土壤环境的变化产生响应,因此成为生态领域的研究热点。土壤微生物主要受到气候因子、土壤因子和植物因子的影响,气候因子主要包括年均温与年降水量[14];土壤因子主要包括土壤pH值[26]、土壤有机碳[27]、土壤全氮[28]、土壤全磷[24]以及土壤含水量[29];植物因子主要包括植物种类以及植物多样性[24]。在城乡梯度的研究中,认为从城区至远郊的建立的空间梯度与通过建立时间梯度来研究环境变化的影响具有相似性,因此可以用空间替代时间的方法研究城市化对城市森林生态系统的影响[30]。在研究土壤微生物上,以高通量测序技术中的操作分类单元(OTU)[15]或扩增子序列变体(ASV)[31]表为主要指标进行,此技术能从某个特定环境内的微生物群落进行全面的鉴定分析,进而精准挖掘出关键因子。
本研究选取国槐(Styphnolobiumjaponicum(L.) Schott)林作为研究对象,在北京市设立城区-近郊-远郊城乡梯度带,分析土壤细菌与真菌群落是否在城乡梯度上存在差异,探究影响土壤微生物群落变化的土壤和气候因子。该研究可为缓解土壤环境压力和建设可持续森林城市提供科学依据。本研究拟解决以下问题:(1)分析土壤细菌与真菌群落在城乡梯度上的响应差异;(2)探究出现响应差异的影响因素。
1 研究地区与研究方法
1.1 研究地区与研究方法
研究样地选取北京市景山公园万春亭为中心,并通过测定各样地距离此中心的直线距离,以此距离为参考选取了七块立地条件相似的森林公园为样地,具体样地信息如表1所示。在梯度设定方面参考了Fang等[32]的设定规则,即将样地距离中心的直线距离在0—20km以内归为城区、20—40km归为近郊、40km以上归为远郊。七个样地整体属于温带大陆性季风气候,四季分明,温暖湿润,年平均气温介于8.76—12.49℃,年降水量介于479—595mm,海拔介于53—673m,土壤类型以褐土、棕壤为主,土壤pH介于6.85—8.50,土壤有机碳介于1.32—4.64g/kg,土壤全氮介于0.05—0.34g/kg,土壤全磷介于0.05—0.17g/kg。

表1 样地概括
1.2 实验设计与野外采样
于2021年7月,在7个公园样地中选取胸径和树高接近的国槐(Styphnolobiumjaponicum(L.) Schott)林,用土钻法在3个10m×10m的样方中随机钻4钻土(10cm钻),土壤深度为0cm—10cm,4钻土混合成1个样。去除石头、根系等杂物,过直径2mm土壤筛,并分为三份:一份保存于超低温冰箱中(-80℃),用于后续土壤细菌和真菌群落的扩增工作和高通量测序;一份保存于4℃冰箱,用于测定土壤pH;另一部分样品放于常温下通风晾干,随后使用工具将其碾碎,并使其通过100目土壤筛,以备进行土壤有机碳、全氮和全磷的测定。
1.3 土壤理化性质测定
土壤pH(土液比1∶2.5)采用pH计测定;土壤有机碳(TOC)采用重铬酸钾容量-外加热法测定,全氮(TN)含量采用半微量凯氏定氮法测定,全磷(TP)采用NaOH熔融-钼锑抗比色法测定。其中年均温(MAT)与年降水(MAP)数据来自于网站(http://worldclim.org)[33],记录了从1970至2020年的年均温与年降水数值。
1.4 土壤微生物高通量测序
土壤微生物群落组成和多样性利用MoBio PowerSoil DNA提取试剂盒(MoBio,卡尔斯巴德,加利福尼亚州,美国)提取DNA,用于提取的土壤总DNA的样品用量为0.5g。对于土壤细菌群落,使用338F/806F引物组来扩增16SrRNA基因(338F,ACTCCTACGGGAGGCAGCAG,806R,GGACTACNVGGGTWTCTAAT),每个样品重复扩增3次;对于土壤真菌群落,使用ITS1F/ITS2R引物对扩增rRNA基因ITS1F区(ITS1F, CTTGGTCATTTAGAGGAAGTAA,ITS2R,GCTGCGTTCTTCATCGATGC)。所有的PCR反应均在20μL反应体系中进行,包括5μm的正引物与反引物各0.8μL、10μg的模板DNA以及20μL的H2O。热循环是指首先在95℃的温度下进行30s的循环,之后以55℃的退火温度进行30s的循环,再在以72℃的温度进行45s的循环,最后以72℃进行10min的处理,待温度达到10℃后停止处理。数据已经以登记号PRJNA893845保存在国家生物技术信息中心(NCBI)序列阅读档案(SRA)中。
1.5 数据处理与分析
测序数据用 QIIME分析平台进行分析。并在 97%的相似度水平上进行了物种多样性和系统发育多样性(PD)分析。物种多样性主要使用操作分类单元(简称OTUs)来描述。选择差异显著法(HSD)检验法分析国槐根际土壤细菌和真菌群落组成、物种多样性和系统发育多样性在不同城市化公园之间的差异,采用Pearson相关系数对土壤细菌和真菌群落组成和多样性与土壤理化性质因子以及气候因子进行相关性分析,使用非度量多维尺度分析(NMDS)分析来检测两个微生物群落的结构是否能够在城乡梯度上区分开。在R i386 3.1.1软件中利用Vegan包使用Mantel分析计算土壤细菌和真菌群落与土壤理化性质因子以及气候因子的相关性,再使用决策曲线分析(DCA)决策曲线分析法对数据进行检测,根据曲线结果选择冗杂分析(RDA)或典范对应分析(CCA)方法分析土壤理化性质因子与气候因子对土壤细菌、真菌群落的解释力。
2 结果与分析
2.1 城乡梯度下土壤理化性质分布特征及其相关性
如表1所示,土壤pH、TOC和TN在城乡梯度上存在显著差异。其中,土壤pH在昌平南口公园(NP)点最高,鹫峰森林公园(JP)点最低。TOC在城乡梯度上呈现城区>近郊>远郊的趋势,具体表现为北京大业大学(BFU)、燕山公园(YP)与八达岭森林公园(BP)三处样地的递减趋势;TN在城乡梯度上呈现城区>近郊>远郊的趋势,在近郊的YP有短暂的回升,随后再次降低;TP在城乡梯度上无显著差异。
2.2 城乡梯度下土壤细菌与真菌群落组成特征
如图1所示,样地土壤细菌群落组成主要有41大类群,其中放线菌(Actinobacteriota)(28.46%)、α变形菌(Alphaproteobacteria)(14.64%)、酸杆菌(Acidobacteriota)(11.99%)、绿弯菌(Chloroflexi)(7.16%)、γ变形菌(Gammaproteobacteria)(6.04%)等5大门类是主要的类群,相对丰度均在5%以上,占据了超过85%的总序列数。此外,拟杆菌(Bacteroidota)(2.37%)、粘球菌(Myxococcota)(1.87%)、芽单胞菌(Gemmatimonadota)(1.65%)、厚壁菌(Firmicutes)(1.21%)等4大门类相对丰度比较低,占据了大约7.10%的总序列数,另外还有32个稀有细菌门类被检测到。样地土壤真菌群落组成主要有16大类,其中子囊菌(Ascomycota)(70.16%)、未分类真菌(Fungi)(9.33%)、担子菌(Basidiomycota)(9.20%)、被孢霉菌(Mortierellomycota)(6.48%)等4大门类是主要的类群,相对丰度均在5%以上,占据了超过85%的总序列数。此外,除罗泽真菌(Rozellomycota)(3.68%)门类的相对丰度在1%—5%之间,其余10门类的相对丰度均在1%以下,处于比较低的水平。
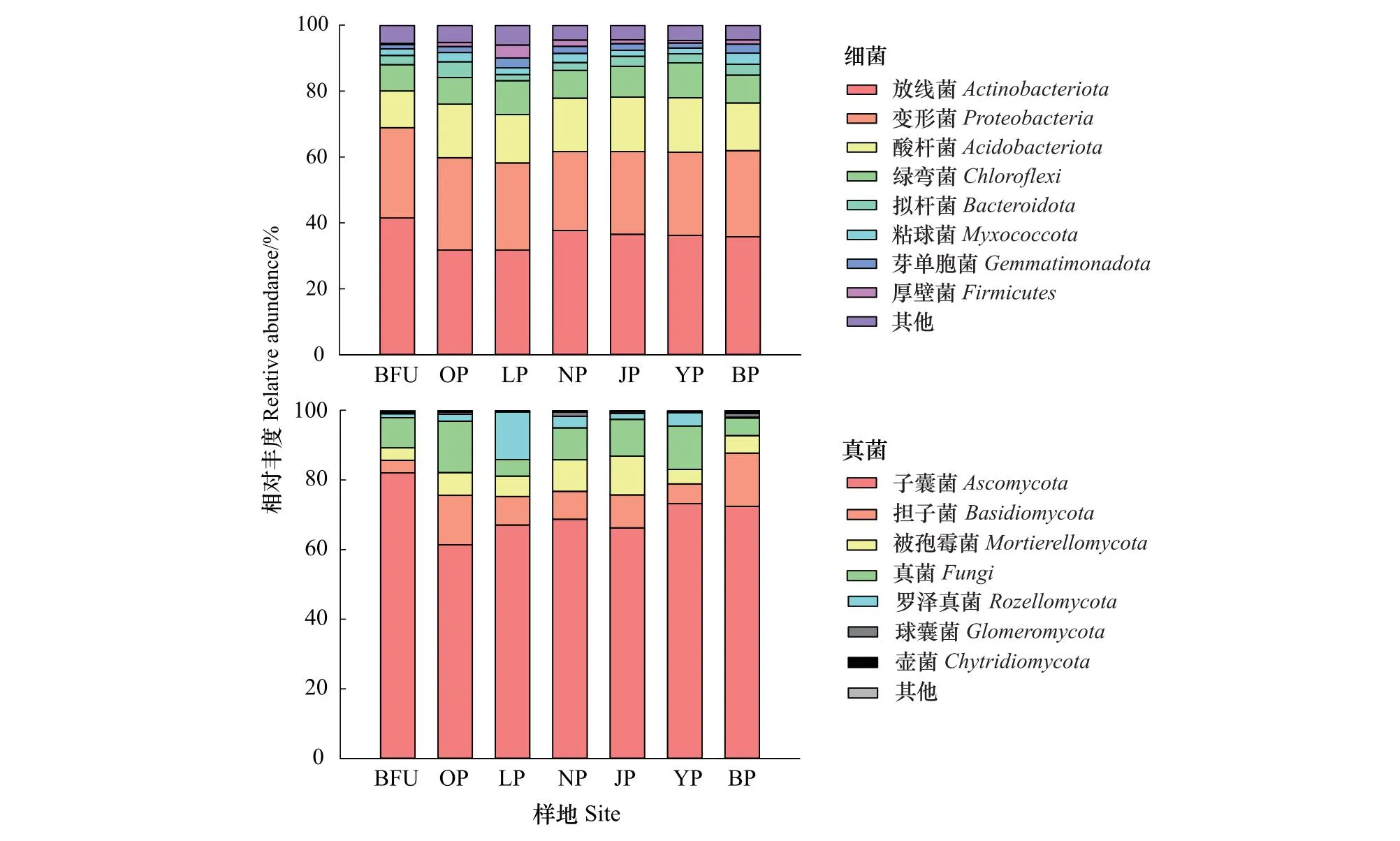
图1 城乡梯度下各样地土壤细菌与真菌主要门类的相对丰度
2.3 城乡梯度下土壤细菌与真菌群落主要门类相对丰度分布特征
如表2所示,城乡梯度下十类土壤细菌主要门类的相对丰度共有五类存在显著差异,分别是放线菌、γ变形菌、拟杆菌、粘球菌与厚壁菌。NMDS分析结果表明(图2),7个样点的土壤细菌群落在近郊样地不能完全分开,而土壤真菌群落可以完全分开。

图2 土壤细菌、真菌群落的非度量多维尺度分析结果

表2 土壤细菌主要门类沿城乡梯度的相对丰度
放线菌的相对丰度在城乡梯度上的趋势表现为随着距离中心的直线距离增加(表2),相对丰度先降低后增加,到达一定距离(30km)后回升到一定水平保持不变;γ变形菌的相对丰度在城乡梯度上表现为城区>远郊>近郊,在城区的奥林匹克森林公园(OP)与近郊的龙潭公园(YP)处出现显著差异;拟杆菌的相对丰度在城乡梯度上整体表现为城区<近郊<远郊,在城区的OP出现最高值,LP出现最低值。粘球菌的相对丰度在城乡梯度上近郊<城区<远郊,其主要差异出现在近郊的YP与远郊的BP中;厚壁菌的相对丰度在城乡梯度上表现为城区>近郊>远郊,在城区的LP出现最高值。
如表3所示,城乡梯度下五类土壤真菌主要门类的相对丰度共有三类存在差异性,分别是未分类真菌、担子菌与罗泽真菌。未分类真菌的相对丰度在城乡梯度上表现为近郊>城区>远郊,在城区中的OP与LP出现显著差异;担子菌的相对丰度在城乡梯度上整体表现为远郊>城区≈近郊,在距离中心最近的BFU与距离最远的BP产生了显著差异;罗泽真菌的相对丰度在城乡梯度上表现为城区>近郊>远郊,其中城区的LP显著高于其他样地。
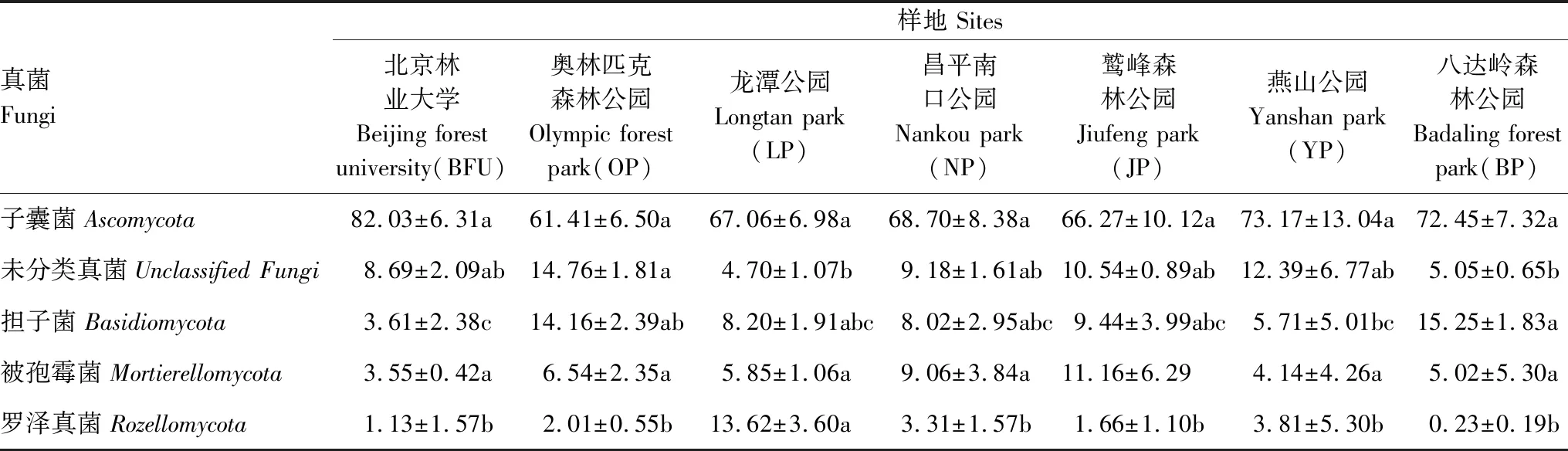
表3 土壤真菌主要门类沿城乡梯度的相对丰度
2.4 城乡梯度下土壤细菌与真菌群落多样性特征
如表4所示, 细菌与真菌群落的OTUs在城乡梯度上均存在差异。对于土壤细菌群落,OTUs表现为远郊>近郊≈城区,具体表现为随着距离中心的距离达到一定数值(30km)后,距离越远,其OTUs数值越高;对于土壤真菌群落,OTUs在7个样地存在显著差异。细菌和真菌群落的PD与Shannon指数在城乡梯度上的变化趋势与OTUs的趋势相似。

表4 各样地土壤细菌、真菌群落物种多样性、系统发育多样性和香农指数
2.5 土壤细菌、真菌群落与环境因子的相关性及响应关系
Mantel分析结果表明(表5),土壤因子(土壤pH、TOC、TP和TN)和气候因子(MAT和MAP)对土壤细菌与真菌群落均产生显著影响。典范对应分析结果表明(图3),TN是土壤细菌与真菌群落最主要的影响因子。

图3 土壤细菌、真菌群落分布的冗杂分析/典范对应分析结果

表5 基于Mantel检验土壤细菌和真菌群落组成与土壤、气候因子的相关性
土壤细菌和真菌群落主要门类的相对丰度、α多样性与环境因子的相关性热图表明(图4),放线菌与土壤pH显著相关;与TOC显著相关的有α变形菌、γ变形菌、芽单胞菌、厚壁菌、子囊菌、担子菌、被孢霉菌以及真菌群落α指数;与TN显著相关的有放线菌、α变形菌、γ变形菌、芽单胞菌、厚壁菌、子囊菌、担子菌以及真菌群落α指数;与TP显著相关的有担子菌;与MAT显著相关的有α变形菌、γ变形菌、拟杆菌、未分类真菌、真菌群落α指数;与MAP显著相关的有粘球菌、担子菌、罗泽真菌以及真菌群落α指数。
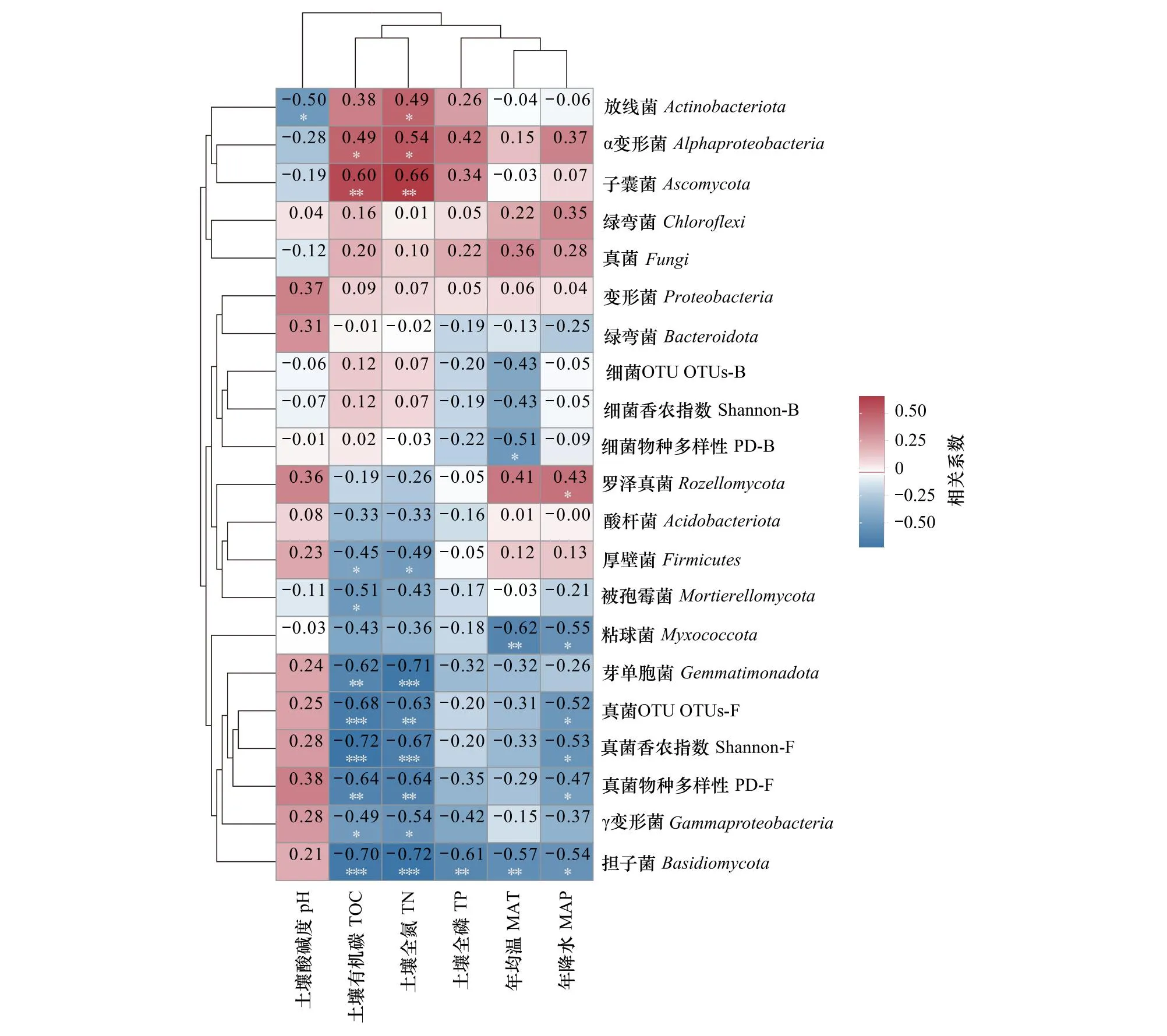
图4 土壤细菌、真菌群落主要门类相对丰度、α多样性与环境因子的相关性热图
3 讨论
3.1 城乡梯度下土壤细菌主要门类的分布特征
从土壤细菌主要门类的相对丰度来看,前五类主要门类分别是放线菌(Actinobacteriota)(28.46%)、α变形菌(Alphaproteobacteria)(14.64%)、酸杆菌(Acidobacteriota)(11.99%)、绿弯菌(Chloroflexi)(7.16%)、γ变形菌(Gammaproteobacteria)(6.04%)。这与前人研究相似[15,18,28]。Tian等[15]发现土壤细菌的主要种类为变形菌与酸杆菌,Hou等[18]发现土壤细菌的主要种类为变形菌与绿弯菌。在这些土壤细菌主要门类中,放线菌、γ变形菌、拟杆菌、粘球菌与厚壁菌在城乡梯度上存在显著差异。
放线菌是一类具有重要生物活性的功能性微生物,其主要功能为产生种类繁杂和富含生物活性的次级代谢产物,能生成各种酶和有机酸等,有助于分解有机物和矿物质,同时具有良好的抗菌活性和耐盐碱作用[34—35],因此能够改良土壤环境,在营养类型菌落中被认为属于富营养菌[36]。本研究发现放线菌在城区BFU、OP以及LP样地存在显著差异,并且受到土壤pH和TN的显著影响。
变形菌被认为在抑制病原菌与胁迫耐受性方面起着关键作用[37]。 Cao等[38]表明变形菌对环境的适应力强,且变形菌属于富营养菌[15]。γ变形菌作为变形菌门中的一类菌群,由于大量植物根基促生菌都属于γ变形菌,因此γ变形菌可以抵抗土壤病原体[31]。本研究发现γ变形菌的相对丰度在城乡梯度上存在显著差异:在城区OP与近郊YP存在显著差异,且OP显著高于YP,同时OP样地的土壤pH和养分指标也均高于YP样地,这与以往研究不一致[18]。Hou等[18]发现变形菌与TOC、TN、TP均为显著正相关、与土壤pH呈负相关关系。本研究发现γ变形菌与TOC和TN呈显著负相关,这与变形菌的富营养性不符合。可能的解释为:一方面,由于变形菌的强适应力,使得相对低养分环境对于γ变形菌造成的环境胁迫力降低;另一方面,在公园管理措施上,OP的人为管理水平高于YP,即使在养分条件劣于YP的情况下,依然出现了OP高于YP的情况。
拟杆菌是土壤环境中养分周转与氮循环的重要菌类,同时参与纤维素的降解过程[3]。在细菌特性上拟杆菌与变形菌相似,对于病原菌和环境胁迫有着良好耐受[37]。在养分需求方面,拟杆菌也属于富营养菌[15]。本研究发现拟杆菌与环境因子之间也无显著相关关系,这与Hou等[18]发现拟杆菌与TOC、TN和TP存在显著正相关而与土壤pH存在显著负相关的结论不一致。可能的原因与变形菌相似,即城乡间不同人为管理方式改变了环境因子对此类有着强环境适应力细菌的影响力。
粘球菌作为一类对土壤养分需求较高的菌群,在营养丰富的条件下会表现出掠夺性,推测它属于一类富营养菌。在表2中,粘球菌的相对丰度表现为远郊BP显著高于其他样地,尤其高于近郊的NP与YP样地。结合样地养分表现为BP≈NP 厚壁菌在土壤细菌群落中属于厌氧菌,可在物料降解中产生对病原微生物有毒的有机酸,有利于土壤健康,在养分充足的情况下会形成良性循环,因此认为此细菌属于富营养菌[39]。同时,厚壁菌在营养相对贫乏的环境中也具有较高的丰度,因此厚壁菌同时具备较强的抗逆性[40]。本研究发现厚壁菌与TOC和TN呈显著负相关,与土壤pH无显著关系,这与Hou等[18]发现厚壁菌与土壤pH呈显著正相关的结论不一致。可能的解释为:本实验中LP的土壤pH仅低于NP,之所以土壤pH未与厚壁菌的相对丰度呈显著相关,可能与因子的变动幅度过小有关,这在Du等[12]中也出现了相似的结果。 本研究发现细菌群落α多样性指数受环境因子影响较小,并且土壤细菌群落在距离中心达到一定距离(30km)时,细菌群落多样性显著上升,这与刘柱等[41]的研究结果相似,认为土壤细菌群落在特定区域的群落组成受到特定环境因子的调控,此特定环境因子可能与土壤容重[42]、细菌之间的拮抗作用[43]以及植物分泌物[24]等因子有关。此外,土壤养分在土壤环境中存在多种形态,其不同菌落对于此类养分也有着不同的偏好[28],可在之后的研究中着重考虑这方面。本研究发现七个样地的细菌群落大部分可在一定程度上完全分开,仅有近郊样地(YP、JP以及NP)有一定重合,表明近郊存在环境重合性,侧面反映出城市化导致环境变化对于细菌群落有着显著影响。另外, CCA结果中横轴解释度为33.69%,纵轴解释度为26.43%,其中TN的线最长,表明TN是影响土壤细菌群落的关键因子。 在土壤真菌主要门类的相对丰度中,前五类主要门类分别是子囊菌(Ascomycota)(70.16%)、未分类真菌(Fungi)(9.33%)、担子菌(Basidiomycota)(9.20%)、被孢霉菌(Mortierellomycota)(6.48%)与罗泽真菌(Rozellomycota)(3.68%)。这样的主要门类相对丰度与前人研究结论相似[13,21,44]。在这些主要门类中,未分类真菌、担子菌与罗泽真菌在城乡梯度上存在显著差异。 子囊菌与担子菌作为典型的陆生型真菌,更适应土壤含水量较低及通气条件好的环境[21],同时均属于腐生营养型菌,在植物纤维和木质素等难以降解的有机质方面承担主要分解角色。早期贫瘠土壤中的真菌群落一般以子囊菌为主,而在后期演替的土壤环境中则以担子菌为主[45]。本研究发现子囊菌的相对丰度在城乡梯度上无显著差异,但以高占比占据了约70%的真菌群落,推测子囊菌相比担子菌其环境耐受力更强。在城乡梯度上存在显著差异的担子菌相对丰度与TOC、TN以及TP均呈现极显著负相关,而无显著差异的子囊菌相对丰度与TOC、TN呈极显著正相关,同时土壤真菌群落α多样性指数也与TOC、TN呈极显著负相关,因此可推测出:(1)由于子囊菌属于营养型菌,因此与TOC、TN等土壤养分因子呈极显著正相关,若此时土壤环境养分充足,菌落对于养分的需求相对会降低,因而可确定此时的土壤环境属于贫瘠前期;(2)担子菌作为与子囊菌同类型的营养菌,其养分相关性与子囊菌相反,这是因为担子菌有着降解顽固有机质的功能[45],因此会与TOC呈显著负相关,而TN的显著相关结果与前人研究成果相同[21],结合子囊菌与TOC和TN的相关关系,表明TOC与TN是显著影响本研究中真菌群落结构分布差异的关键因子;(3)占比极高的子囊菌与TOC与TN呈极显著正相关,而真菌群落多样性与TOC、TN呈极显著负相关,表明占比较小的其余真菌与TOC、TN呈极显著负相关的偏向程度较大,因此可能有其他因子,如真菌与植物形成的共生关系[22]以及植物凋落物影响[21]等影响真菌群落,这些方面有待后续研究。在真菌群落α多样性指数中,OTUs和PD指数均与TOC、TN呈极显著负相关关系,结合真菌群落在城乡梯度上表现为近郊>城区>远郊,同时NMDS结果表明每个样地的真菌群落均能完全分开,因此可推测城市化导致的环境差异对真菌群落的影响与细菌具有明显差异。 与真菌群落相比,细菌群落在城乡梯度上存在显著差异的菌落数量多于真菌群落;细菌群落与土壤pH、TOC、TN以及MAT有着显著相关性,而真菌群落与TOC、TN、TP以及MAP存在显著相关性;细菌群落α多样性指数在远郊处多样性显著上升,而真菌群落α多样性指数在近郊处多样性最高;细菌群落在近郊不能完全分开,而真菌群落可以完全分开。因此可推测出城乡梯度下的土壤细菌群落更容易受到环境因子的影响。 (1)土壤细菌群落与真菌群落在城乡梯度上存在显著差异; (2)土壤pH、TOC、TN、TP、MAT以及MAP对于土壤细菌与真菌群落均有显著影响。其中土壤细菌群落分布的环境因子包括土壤pH、土壤有机碳、土壤全氮以及年均温,而影响土壤真菌群落分布的环境因子包括土壤有机碳、土壤全氮、土壤全磷以及年降水; (3)土壤细菌群落在城乡梯度上不能完全区分开,在近郊处出现群落组成重合,而土壤真菌群落可以完全区分开; (4)土壤细菌群落相比真菌群落更容易受到环境因子的影响。3.2 城乡梯度下土壤真菌主要门类的分布特征
3.3 城乡梯度下土壤细菌、真菌群落分布特征对比
4 结论
