烃源岩母质之海洋有机质的生产及其控制作用
2023-12-25黄臣军刘格云
黄臣军, 刘格云
1. 中国石化石油勘探开发研究院, 北京 100083; 2. 河北地质大学, 河北 石家庄 050031; 3. 河北省岩石矿物材料绿色开发重点实验室, 河北 石家庄 050031
在一般人看来, 海洋似乎是无限的, 在其中生活着大量的鱼类和其他生物。 然而, 随着对海洋越来越了解, 就会发现并非所有海域都是一样的。 事实上,大部分海洋几乎没有海洋生物, 而其他地区, 特别是沿某些海岸的地区, 生产力要高得多。 在主要大陆的西海岸发现了生产力最高的区域, 这些区域仅占全球海洋面积的1%左右, 却生产了全球约20%的鱼类,这是由于上升流系统带来了丰富养分的结果。 四个主要的西海岸上升流系统位于北太平洋的加利福尼亚—俄勒冈州—华盛顿、 南太平洋的秘鲁和智利、 北大西洋的西北非洲, 以及南大西洋的南非和纳米比亚。 在全球海洋中还存在许多其他的上升流系统, 其中一些是全年性的, 而另一些是季节性的。 它们在全球生产力、 生物地球化学循环和全球气候变化下的食物网动态中具有重要作用。
1 海洋有机质生产作用
阳光是地球上几乎所有生命(包括深海中的生命) 的直接或间接能源。 然而, 光被海水吸收和散射, 很少有光能穿透约80 m 的深度以下(在最低生产力的亚热带地区深达150 m, 但在高生产力和沿海地区仅深达10 m) (图1)。 因此, 光合作用主要限于海洋的上部透光表层。 在大部分海域, 包括热带、 亚热带和温带, 吸收阳光导致表层水比深海水温暖得多。 暖水比冷水浮力大, 这导致阳光透射表层水在密度更大的深层水上漂浮, 两者之间的过渡层称为“密跃层” 或“温跃层”。 因此, 海水及其溶解的化学物质在阳光透射的表层与黑暗的深层之间的传输是滞缓的。 光的这种双重作用(光合作用和海水浮力) 对海洋浮游植物的成功至关重要。 光合作用限制了活跃的浮游植物生长在海洋的表层, 而海水密度分层阻止了它们混入黑暗的深海。 因此, 大多海洋生物, 包括浮游植物、 浮游动物和游泳动物, 主要分布在海洋表层约200 m 的深度范围。
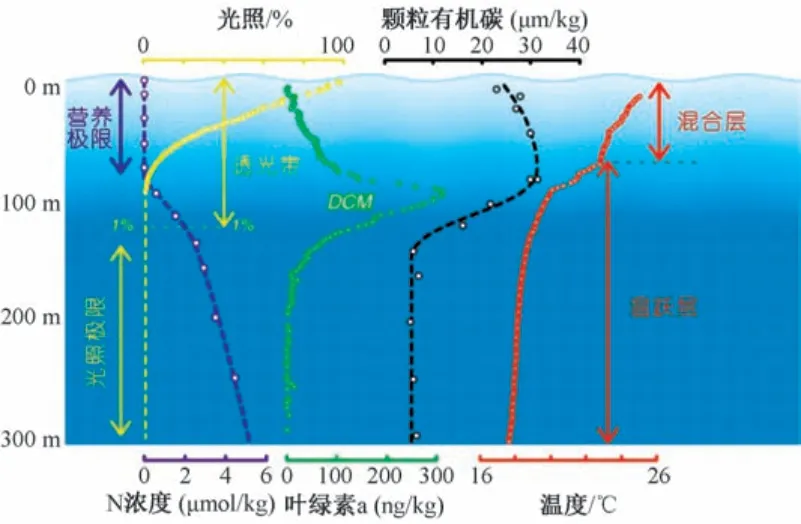
图1 亚热带开阔海洋光合作用环境特征(据文献[1] 修改)Fig.1 Characteristics of photosynthetic environment in subtropical open ocean
1.1 海洋生产力要素
海洋生产力主要是指“浮游植物” (悬浮在海洋中的植物, 其中大部分是单细胞藻类) 产生的有机物质。 浮游植物是“光合自养生物”, 其吸收光能以将无机碳转化为有机碳, 并将这种有机碳提供给不同的“异养生物”, 即仅通过有机物质的呼吸作用获取能量的生物。 开阔的海洋异养生物包括细菌、 以及更复杂的单细胞和多细胞“浮游动物”、 游泳动物(鱼类和海洋哺乳动物) 和海底生物群落。
浮游植物生长需要一套化学物质, 而那些在表层海水中可能稀缺的物质通常是“营养物质”。 重要的营养元素包括氮 (N)、 磷 (P)、 铁 (Fe) 和硅 (Si)。浮游植物对氮和磷的需求相对统一。 在1900 年代初期, 海洋学家阿尔弗雷德·雷德菲尔德 (Alfred Redfield) 发现浮游生物以~106 ∶16 ∶1 的C ∶N ∶P化学计量比建立生物量, 称为雷德菲尔德比 (Redfield ratio)。 铁在生物质中的含量仅为微量, 但它在生物体中用于多种基本目的, 铁的稀缺性通常会限制或影响广海的生产力, 尤其是那些高纬度地区。 硅是特定浮游生物分类群—硅藻(自养, 浮游植物)、 硅鞭毛藻和放射虫(异养, 浮游动物) 的营养物质, 即被用来制造蛋白石硬质体。
初级生产是从无机底质中产生新的有机物, 正是这种有机物成为大多数海洋消费者食物网的基础。 初级生产通常是指光合作用, 或根据反应由植物和藻类利用光能生产化学燃料:
在这里, 以光能为动力, 二氧化碳和水结合在一起产生葡萄糖和氧气。 但是, 在没有光照的情况下,细菌也会进行初级生产化学合成, 其能量不是由光提供反应的能量, 而是来自无机材料的氧化, 例如硫化氢。 光合作用在总海洋生产力中所起的作用远大于化学合成。
负责海洋初级生产的生物包括各种各样的海洋植物和藻类。 海洋中最大的光合作用来自微观藻类(浮游植物)。 生产者创造的有机质总量称为初级生产力。但是, 初级生产者自己通过呼吸消耗了一部分这种有机物质。 因此, 剩下的用以支持海洋中的消费者的总量称为净生产力(总生产-呼吸)。 总生产可以分为两个部分, 新生产和再生生产。 新生产是通过上升流或洋流从本地生态系统外部引入营养支持。 再生生产是生态系统中养分循环的结果。
1.2 上升流供给养分
对于浮游植物而言, 生产力取决于光和养分的利用率。 光对于光合作用至关重要, 浮游植物和其他主要生产者仅限于海洋表层, 那里的光量足以维持反应。 随着深度的增加, 光强度会降低, 一直到无法再进行光合作用的深度为止。 足够光合作用的光可以穿透的深度称为透光带, 通常向海面以下延伸至约80 m水深。
除进行光合作用外, 浮游植物还呼吸, 消耗了它们产生的一些有机化合物。 呼吸速率与光无关, 呼吸发生在所有深度及光照水平。 因此, 随着深度的增加, 光合作用的速率会随着光的减弱而降低, 一直达到光合作用的速率等于呼吸速率的平衡深度。 这个深度是补偿深度, 它标志着净初级生产结束。 在此深度以下为净呼吸。
所有海洋初级生产者都需要营养。 浮游植物所需的主要营养物质是氮和磷, 其形式为硝酸盐()、 亚硝酸盐()、 铵(NH4+) 和磷酸盐()。 许多浮游植物, 特别是硅藻, 还需要二氧化硅来形成壳。 所有这些养分都少量地存在于海水中。因此, 在大多数情况下, 它们通常是浮游植物生长的限制因素, 尤其是氮化合物。
营养在整个水柱中不均匀分布。 在表层水中的营养物质迅速被浮游植物利用, 因此, 表层水通常是缺乏营养的。 但是随着浮游植物的消耗或死亡, 它们转变为有机物颗粒, 例如粪便颗粒或尸体, 这些颗粒沉入更深的水中。 一旦进入深水中, 这些物质的分解会将养分释放回水柱中。 由于没有生产者可以深入利用它们, 因此, 深层水中的养分更加丰富。
这些深层营养物质是浮游植物无法到达的层面。温跃层水柱的密度分层通常会阻止营养丰富的深层水与表层水混合。 但是, 在特定条件下, 这种富含营养的深层水可以通过上升流被带到表层。 因此, 浮游植物可以利用这些输入的养分, 从而形成高生产力。
养分再生在上升流区域起着重要作用。 它主要通过两个过程发生, 即沉积物—水界面和水体中的细菌再生, 以及食藻动物的牧食活动(图2 说明了再生过程)。 氮作为溶解硝酸盐的供应, 使得能够区分由硝酸盐驱动的“新” 生产和由回收铵和尿素驱动的“再生” 生产。 在开阔的海洋中, 营养再生利用率通常约为 10%。 然而, 在海岸上升流地区, 它可能高达80%[2]。 上升流地区的生产力提高直接源于新生产中上升流硝酸盐的持续可用性, 而其他沿海地区或广海则依赖较少量的循环氮。

图2 海洋上升流区磷、 氮和二氧化硅循环及生物吸收和再生模式(据文献[3] 修改)Fig.2 Phosphorus, nitrogen and silica circulation and biological absorption and regeneration patterns in the marine upwelling zones
2 上升流机制及其分布
上升流是将更深的水带到表层的过程, 其重要意义是带来了富含营养的深层水, 从而促进初级生产。海洋中的营养物质浓度通常随着水深的增加而增加,因此, 将营养丰富的海水从上陆坡越过陆架坡折处搬运到大陆架上的动力作用, 可以大大提高上升流区域的生物生产力。 一些上升流系统, 例如, 在秘鲁/智利、 加利福尼亚、 非洲西北部和非洲西南部发现的世界上最大的上升流区域, 在大陆架坡折深度显示出富含营养的底流, 既是上升流水的来源, 也是一个营养汇。 上升流的水营养丰富, 因此导致高生产力。 海洋中许多生产力最高的地区都位于上升流区。
表层水的运动在深水垂直运动中起着重要作用。上升流发生在表层洋流离散的地方。 随着表层水的离散, 需要有深层的水上升至表层加以补偿, 从而形成上升流。 在赤道太平洋, 南、 北半球信风吹向赤道并逐渐偏转至平行赤道方向, 埃克曼传输使表层水在南、 北半球中分别向南和向北移动, 形成赤道发散带, 是发生上升流和形成富营养表层水的区域(图3)。
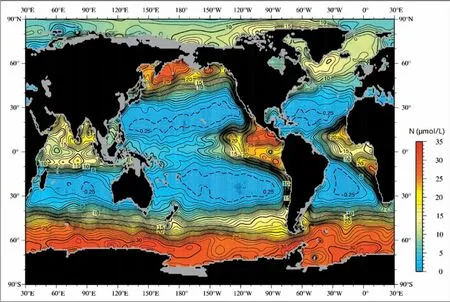
图3 现代海洋100 m 深的年平均氮含量等值线图[4]Fig.3 Contour map of the annual average nitrogen content at depth of 100 m in the modern ocean
当风将表层水移离海岸时, 上升流也发生在海岸地区。 一些生产力最高的商业捕鱼场即与海岸上升流有关。 沿着加利福尼亚海岸, 当地盛行的风向南吹。埃克曼传输将表层水偏转90°到风的右边, 这意味着净埃克曼传输是离岸方向。 海岸附近移走的表层水被冷的、 营养丰富的深层水代替, 从而提高了生产力。
2.1 风力上升流机制
上升流能保持相当长的时间(几天到几周), 足以将水团提升超过100 m 或更大的垂直距离。 风驱动海岸上升流机制, 创造了世界海洋中最大和最持久的上升流区域。 风力驱动上升流的主要形式是(a) 海岸上升流(b) 赤道上升流和(c) 冰缘上升流(图4)。 地球的自转及其相关效应, 例如科里奥利力, 在这三个系统的上升流动力学中都起着主导作用。
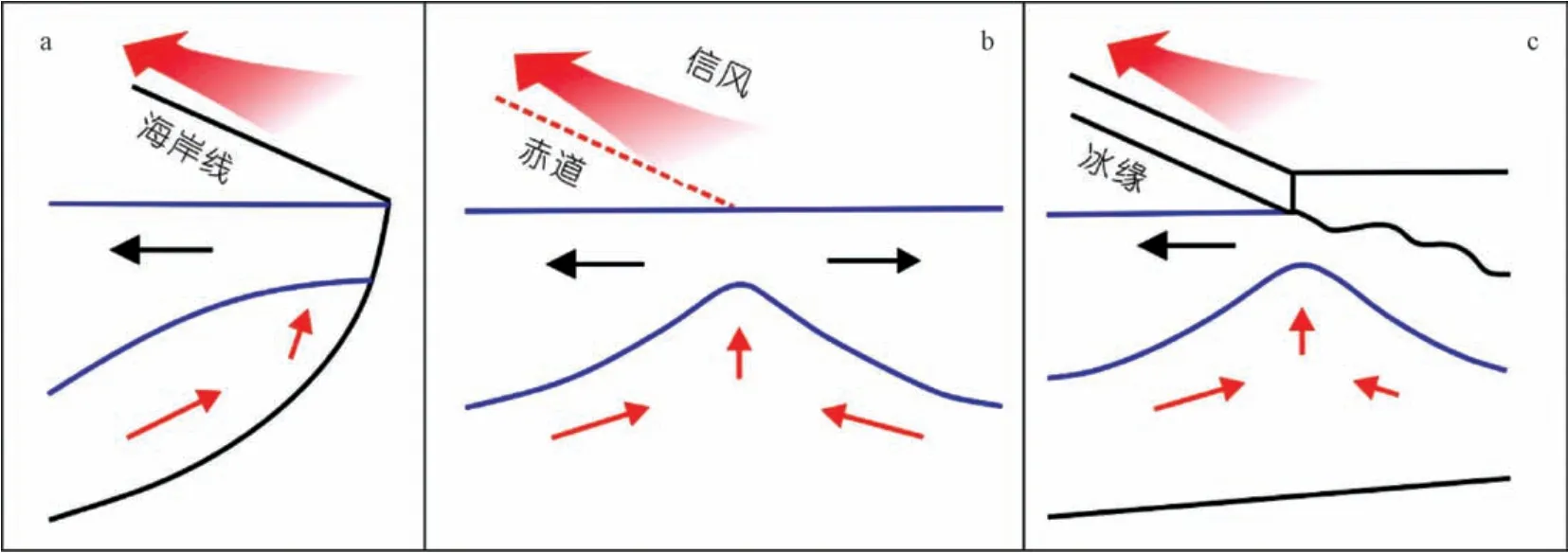
图4 风力上升流机制类型: (a) 海岸上升流、 (b) 赤道上升流和(c) 冰缘上升流(据文献[5] 修改)[红色宽箭头表示盛行风相对于海岸(南半球情形, 南风)、 赤道或冰缘的方向]Fig.4 Types of wind upwelling mechanisms: (a) coastal upwelling, (b) equatorial upwelling and (c) periglacial upwelling
根据一个地区的典型风况, 海岸上升流可以是准永久性的, 即全年性海岸上升流; 也可以是季节性的, 即季节性海岸上升流。 虽然上升流可以沿直线海岸线发生, 但由于海岸线和海底经常不规则, 风驱动的海岸上升流事件通常是局部的。 因此, 上升流在上升流中心的某些地区比其他地区更为明显。 上升流中心通常与强烈的锋面流有关, 上升射流(Jet stream)分化成中尺度(10 ~20 km) 环流模式, 称为涡流。大部分初级生产力发生在海岸上升流中心的内部和下游不远处。
上升射流在数天到数周的时间尺度上迅速变得动态不稳定, 并转化成中尺度涡流。 中尺度涡流是圆形流动模式, 其中地转流围绕与升高(降低) 海平面相关的高压(低压) 中心。 在低压涡流中心主要维持上升水流。 沿海海洋中的涡流直径可小至10 ~20 km,而广海涡流直径可达300 km。 作为涡流倾泻的结果,海岸上升流带的宽度通常沿上升射流的方向增加。 充分发展的涡流场表现出特定的路径, 称为纤流(Filaments), 上涌的水沿着这些路径形成离岸平流(图5)。 纤流可以是准静止或瞬态特征, 通常作为有机质向广海的输出机制运行。 涡流还可以将上升流中心的特性(如: 热异常、 有机质和浮游动物和鱼类幼虫) 分散到近海。
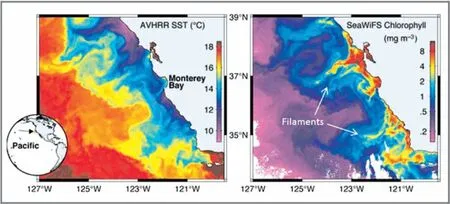
图5 加利福尼亚上升流系统中海面温度(左) 和叶绿素a 浓度(右) 的卫星图像,表现了一个完全发展的涡流场和上升流纤流[6]Fig.5 Satellite images of sea surface temperature (left) and chlorophyll a concentration (right)in the California upwelling system, showing a fully developed vortex field and upwelling filaments
2.2 其他上升流机制
除了风力驱动的海岸上升流作用外, 还有许多其他物理机制可以在沿岸海洋中产生上升流。 包括: 埃克曼泵、 动力上升流、 潮汐引起的上升流、 岛屿之后的上升流、 地形引起的上升流(如在陆架坡折峡谷中), 以及由沿岸受迫波浪通道引起的瞬时上升流。
埃克曼泵是指与空间变化的水平风应力场相关的密跃层的垂直调整, 称为风应力旋卷。 除了海岸的边界效应外, 它还引起水平埃克曼传输的发散。 虽然海岸上升流作用涉及埃克曼层动态固有的跨陆架传输,但埃克曼泵仅引起垂直流动。 埃克曼泵对西海岸上升流区的垂直养分通量有显著贡献(〉25%)。 风应力和风应力旋卷效应之间的重要区别在于, 后者引起的埃克曼泵发生在离岸更远的地方, 通常在陆架边缘, 并且在垂直输送方面比埃克曼传输发生的速度更快。 因此, 它有别于近海埃克曼传输, 可以为系统提供额外的营养输入。
埃克曼泵及其相关的向上养分通量可见于加利福尼亚洋流[7,8]、 本格拉洋流[9]、 东班达海[10], 以及Cabo Frio 周围的上升流系统[11]。 Chavez 和Messié[12]认为这种类型的上升流具有不同的生化意义, 因为它不会吸收与乳浊层相互作用的陆架和斜坡水, 仅能将较低浓度的铁离子带入透光区。 因此, 海岸上升流作用刺激了较大生物(硅藻、 桡足类、 磷虾) 的食物网和生物量的积累, 而风应力旋卷引起的离岸较远的上升流导致相对高营养低叶绿素区域的低生物量和生态系统特征。
动力上升流(也称陆架坡折上升流) 是一种沿大陆东海岸(沿岸海风通常不利于上升流) 的近岸边缘运行的上升流机制。 东海岸流是与温跃层从海洋向海岸急剧上升相关的地转锋流。 作为大型海洋环流的一部分, 它们的动力在很大程度上与当地的风况无关。一般而言, 东海岸流位于大陆架的近岸处, 它们的影响不会深入到远岸海洋, 它建立了以局部风和温盐压强为主的独立环流。 水流位置的变化(例如由涡流形成引起) 可能导致更深的水上升到外陆架上。 该作用适用于所有东海岸流, 可见于两个半球的所有海洋。
潮汐引起的“上升流”, 也称为潮汐泵, 是一种区域性作用。 在这种作用中, 振荡的潮汐流有规律地将富含营养的更深水移动到陆架坡折处。 潮汐泵在位于陆架坡折带的岛屿之间的潮汐通道中的强潮汐流区域最为明显。 虽然这个过程是振荡而不稳定的, 但它导致外陆架上有规律地出现富含营养的水, 因此构成了一种陆架坡折上升流。
陆架坡折峡谷是穿过陆架坡折的海底峡谷。 沿着峡谷方向, 大陆架坡折深度的环境流可以触发较深的水从上陆坡到陆架的局部上涌[13,14]。 这种局部上升流源自与地形罗斯比波(Rossby waves) 传播方向相反的环境流动。 也就是说, 在北(南) 半球的左侧(右侧) 具有较浅水(和海岸线) 的环境流。 地形罗斯比波是由水柱拉伸/挤压地形起伏造成的准地转流动扰动。 虽然这些波浪本身只能在一个特定的沿岸方向传播, 但环境流动可以通过产生驻波模式来抵消这种传播, 驻波模式由跨大陆架坡折的陆上和海上流动的固定但空间交替区域组成。 因此, 峡谷上升流作用可以解释为静止地形罗斯比波的特征, 这可能会在峡谷下游形成的“波峰” 附近引起局部陆上流动[15]。从技术上讲, 坡折峡谷的局部上升流也是陆架坡折上升流的一种形式。
海岸受迫波浪是季节内变化(30~60 天周期) 的波浪, 在海岸附近达到最大振幅, 并与跨大陆架边缘的陆上/海上流动一起引起瞬时上升流/下降流。 这些波浪通常由沿岸风产生, 但也可能由撞击沿赤道东侧海岸的赤道开尔文波产生。 海岸受迫海浪可以沿着海岸线传播数千公里。 在强烈的上升流事件之后, 上升流中心附近海洋压力场的区域性扰动(包括海平面和密度异常) 可能会引发沿陆架坡折传播的海岸受迫波, 并沿途引起瞬时上升流。 这在厄瓜多尔、 秘鲁和智利海岸的沿海潮汐计海平面记录中, 以及在智利中北部系泊处测量的洋流中观察到[16,17]。
2.3 主要上升流区域
当运用上升流是由海岸平行风在表面埃克曼层中驱动离岸传输产生的规则时, 大多数主要的全年性和季节性的海岸上升流系统可以根据表面风型图(图6) 推断出。 这种直观的方法确定了加利福尼亚、 秘鲁、 智利、 非洲西北部和西南部沿海的主要上升流区域及其相关的上升流(即加利福尼亚洋流、 秘鲁—智利洋流、 加那利洋流和本格拉洋流) (图7)。 这些上升流区被称为主要海岸上升流区或西海岸上升流区。虽然这些上升流区的盛行风吹向赤道方向, 但这些系统的一个显著特征是存在向极地的底流, 其在这些区域的营养物质和溶解氧的分布中发挥着重要作用, 也影响海洋生物的横向迁移。

图6 地球表面风型模式的季节性变化[18]: (a) 冬季, (b) 夏季Fig.6 Seasonal changes of wind patterns on the earth's surface: (a) winter, (b) summer
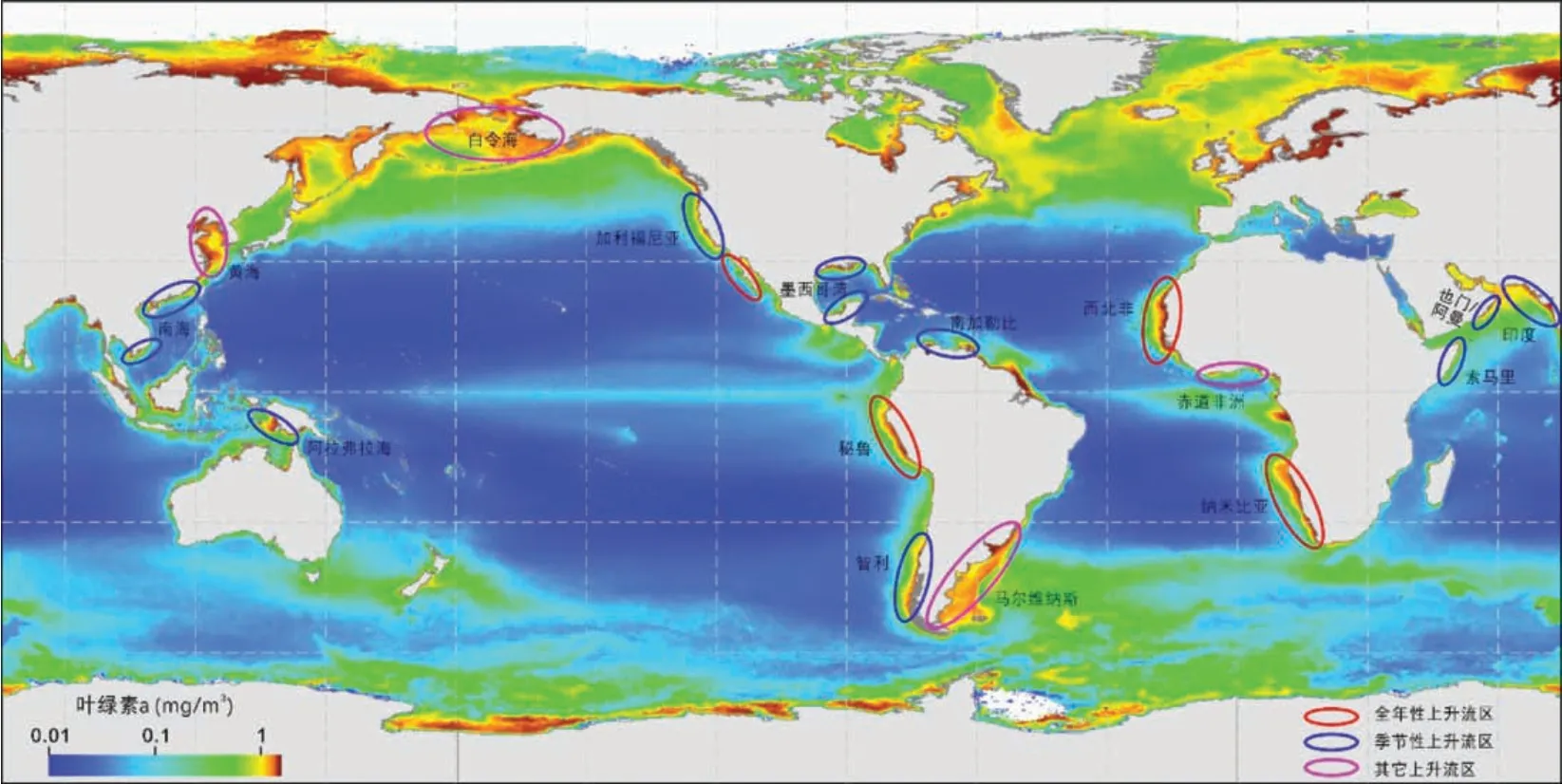
图7 现代海洋表层叶绿素a 年平均浓度(数据来源[19], 代表生产力大小) 及主要的上升流区分布图[5]Fig.7 Annual average concentration of chlorophyll a on the surface of modern oceans (data source from [19];representative of productivity) and the distribution map of main upwelling areas
依据这种方法还确定了索马里、 也门/阿曼、 苏门答腊和南中国海沿岸的季节性上升流系统。 沿着印度西南海岸和澳大利亚南部大陆架也有季节性的海岸上升流系统, 这在图6 所示的风型分布中不太明显。风型图还包括其他类型的上升流/高生产力沿海地区。例如: 白令海的“绿化带”、 格兰德滩、 厄加勒斯滩和马尔维纳斯海流的陆架坡折区, 上升流为动态隆起或陆架坡折上升流; 在爱尔兰海和黄海, 潮汐混合锋导致养分富集。
3 结 论
海洋有机质是烃源岩母质的主要类型之一, 海洋有机质生产力受控于浮游植物对光照和养分的利用率。 足够光合作用的光照深度约80 m, 称为表层透光带。 透光带温水与深部冷水之间形成密跃层而阻碍营养物质向上补给。 因此, 海洋透光带通常是贫营养的, 不利于发生光合作用。
上升流是一种水团的垂向运动, 其可以突破密跃层而将深部营养物质携带至透光带, 从而促进光合作用来提高有机质生产力。 上升流的一种主要机制类型是风力上升流, 又分为全年性和季节性上升流。 这些重要上升流系统分布在低纬度主要大陆的西海岸(例如: 北太平洋加利福尼亚海岸, 南大西洋纳米比亚海岸等), 在那里具有显著的高生产力。
