Cd 胁迫下水稻Cd亚细胞特征及游离氨基酸含量变化分析
2023-12-25林红梅李齐向
林红梅,李齐向
(1.闽江学院地理与海洋学院,福建 福州 350108; 2.福州海洋研究院,福建 福州 350002;3.三明市种子站,福建 三明 365000)
Cd(cadmium,Cd)是元素周期表中第五周期ⅡB的金属元素,地壳中Cd主要以硫Cd矿物的形式存在。随着工业革命后化石燃料燃烧及有色金属的开采增加以来,被释放的Cd通过空气、水等途径进入土壤,造成矿区附近的耕地受到不同程度的Cd污染,而且,农业生产上,污水灌溉、污泥农用等一系列活动进一步加剧农田土壤Cd污染程度。水稻是全球人口的主要粮食作物之一,也是Cd吸附能力较强的农作物之一,人体过量的摄入Cd容易导致肾损伤、骨损伤、肝脏损伤,甚至致癌、致突变等。近年来,水稻受Cd污染的事件越来越受到重视,探索水稻应答Cd胁迫的响应机制有助于降低或防止种植Cd污染的稻米。
植物长期或短期暴露于Cd环境中均能引起植株异常的Cd毒性反应。研究显示,当水稻受到高于0.05 mmol·L-1Cd浓度胁迫时即能产生明显的损伤[1],表现出叶绿体结构破坏、色素合成受阻,光合作用和呼吸作用发生改变,物质和能量代谢发生紊乱。与此同时,植株体内也能产生一系列的生理响应来缓解Cd的毒害。细胞壁是阻挡Cd进入细胞内的第一道屏障[2],而且Cd在植物组织中有选择性的分布能有效减少Cd胁迫对植物产生的伤害。目前Cd胁迫下水稻组织Cd的亚细胞分布研究报道较少。此外,作为多种天然产物的前体,氨基酸在植物受到胁迫时的反应中起到至关重要的作用[3]。在生物与非生物胁迫下,作为机体生长发育最重要的代谢物质-氨基酸及其衍生物通过调节基因表达和蛋白磷酸化过程,改变氨基酸的代谢过程和代谢水平,代谢物响应变化显示了植物体对抗Cd胁迫所做出的防御机制。因此,通过监测氨基酸代谢的变化,可以更好地了解Cd胁迫程度及其相关机制。
因此,本研究以水稻Nipponbare和Lemont为研究材料,分析不同Cd处理水平对水稻叶片Cd的亚细胞分布以及游离氨基酸生理代谢的影响,探讨不同水稻品种对Cd胁迫在物质积累及Cd积累效应上的响应差异,以期揭示水稻应答Cd胁迫的重要防御机制,为低Cd水稻种质资源筛选以及Cd污染农田的生态修复提供参考。
1 材料与方法
1.1 材料与试验设计
供试材料为Nipponbare及Lemont两个水稻品种,Cd为CdCl22.5H2O分析纯试剂。试验于2023年5月闽江学院教学试验基地进行。首先,选取饱满健康的水稻种子经自来水清洗后浸泡24 h,经10%次氯酸钠消毒及双蒸水冲洗后,置30℃培养箱催芽,随后将种子均匀播于装满湿润细沙的砂盆中。待3叶1心时,选取健壮且长势一致的幼苗移栽至塑料盆中。水稻植株由打孔的泡沫板及海绵固定,塑料盆含2 L营养液,营养液配方参照Yoshida等[4]方法配置,pH调至5.6,每盆栽种6棵水稻幼苗。待水稻幼苗恢复生长1周后,以正常水培且未加Cd处理的水稻为对照组(CK),添加5 μmol·L-1(Cd5)、50 μmol·L-1(Cd50)和100 μmol·L-1(Cd100)Cd营养液培养的植株为处理组,试验设置3个重复,处理时间为7 d,处理结束后采其植株叶片,液氮速冻,用于光合色素、Cd亚细胞分布及游离氨基酸含量测定。
1.2 测定方法
1.2.1水稻叶片叶绿素和类胡萝卜含量测定 叶绿素和类胡萝卜素含量测定采用比色法。取0.2 g水稻鲜样置于含少量石英砂和碳酸钙粉的研钵中,加入95%乙醇研磨浸提至组织变白,静置3 min;通过乙醇湿润过的滤纸过滤至25 mL棕色容量瓶中、摇匀,即为色素提取液。取适量提取液移入1 cm光径比色杯,测定470、649和665 nm波长下吸光度,同时以未加样品的95%乙醇溶液为空白对照。
1.2.2水稻叶片Cd亚细胞分布 不同的细胞组分分离参照宋阿琳等[2]方法(差速离心法)略有改动。取0.5 g鲜样于研钵中,加入250 mmol·L-1蔗糖、50 mmol·L-1Tris-HCL缓冲液(pH 7.5)及1 mmol·L-1DTT,低温快速研磨至匀浆,移入离心管,离心,转移上清液1,剩余残渣为细胞壁组分;4℃、16000 r·min-1离心上清液1,转移上清液2,剩余部分为细胞器组分,上清液2为细胞质组分。细胞壁、细胞器和细胞质组分置干燥箱中烘干,参照Lin等[5]方法进行微波消解以及电感耦合等离子体质谱仪(ICP-MS)测定Cd元素含量。
1.2.3水稻叶片游离氨基酸测定 参照郑瑞生等[6]测定水稻叶片游离氨基酸含量。新鲜样品加入70%乙醇,100℃回流浸提4次,浸提液去乙醇、脱色、蒸干,pH2.2、0.1 mol·L-1柠檬酸缓冲液溶解,经0.22 μm滤膜过滤后,采用氨基酸全自动分析仪测定。
1.3 数据处理
数据采用EXCEL整理、DPS统计软件进行统计分析。
2 结果分析
2.1 Cd胁迫对水稻叶片色素含量的影响
由表1可知,随Cd处理浓度增加,叶绿素和类胡萝卜素含量均不断下降;与CK相比,Cd5处理显著降低Nipponbare叶绿素含量,降幅达6.17%,类胡萝卜素含量未发生显著变化;Cd50和Cd100显著降低2种水稻叶绿素和类胡萝卜素含量,其中Nipponbare叶绿素降幅分别为20.13%和50.32%,降幅大于Lemont(分别为14.06%和39.62%);Nipponbare类胡萝卜素含量降幅分别为41.94%和67.74%,降幅大于Lemont(分别为29.41%和52.94%)。对比2种水稻色素含量,Lemont水稻叶绿素和类胡萝卜素含量均大于Nipponbare,且Cd50和Cd100处理时差异达显著水平。
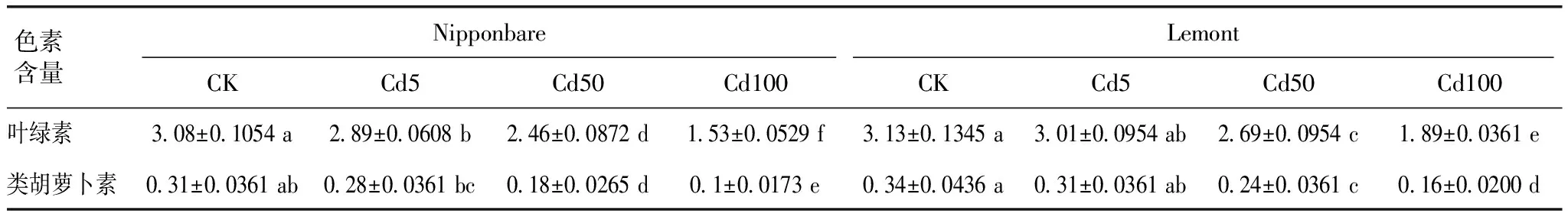
表1 不同Cd浓度处理下水稻叶片叶绿素和类胡萝卜素含量
2.2 Cd胁迫对水稻叶片Cd亚细胞分布的影响
由表2可知,随Cd处理浓度增加,2种水稻各亚细胞组织Cd含量不断增加,与CK相比,当Cd5、Cd50和Cd100处理时,Nipponbare水稻各亚细胞组织Cd含量差异均达显著水平;当Cd50和Cd100处理时,Lemont各亚细胞组织Cd含量也达显著水平;各亚细胞组织Cd含量表现为细胞壁>细胞质>细胞器,表明细胞壁是水稻积累Cd的主要部位;其中,不同Cd浓度处理下Nipponbare细胞壁Cd含量占总Cd比例为86.24%~88.59%,Lemont为88.66%~94.17%。对比2种水稻Cd亚细胞分布可知,Nipponbare水稻显著高于Lemont水稻。
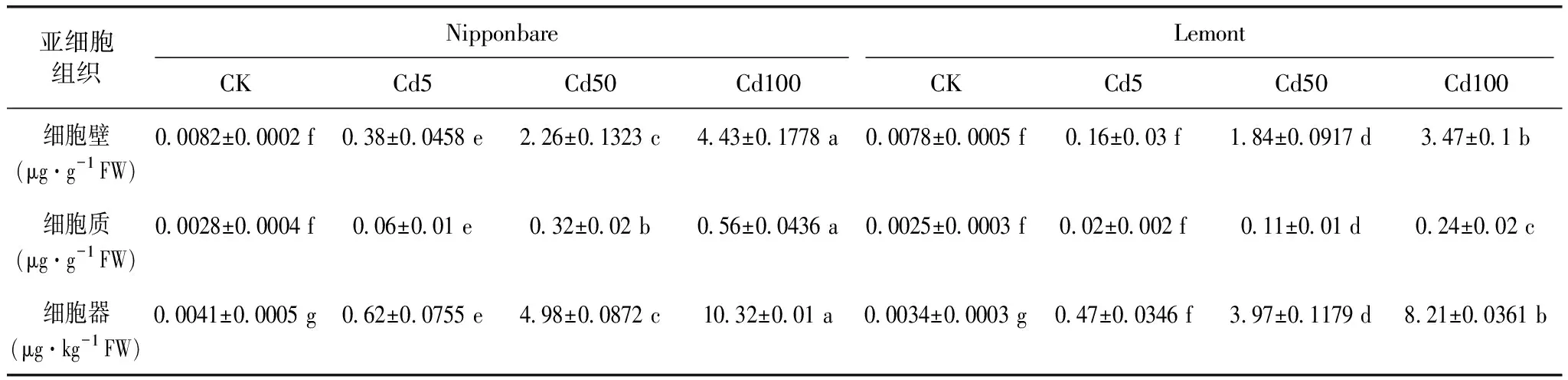
表2 不同Cd处理下水稻叶片Cd亚细胞分布
2.3 Cd胁迫对水稻叶片游离氨基酸含量的影响
由表3可知,当Cd5处理时,Nipponbare水稻甘氨酸和Lemont水稻脯氨酸发生显著改变,其他游离氨基酸含量均未发生显著改变。与CK相比,当Cd50和Cd100处理时,Nipponbare水稻脯氨酸、半胱氨酸及谷氨酸含量随Cd处理浓度增加而显著增加,增幅为77.27%和139.61%、62.5%和112.5%、45.37%和67.22%;当Cd50和Cd100处理时,Lemont水稻脯氨酸、半胱氨酸及谷氨酸增加25.7%和84.92%、57.14%和107.14%、38.69%和68.59%;Cd50处理显著提高两种水稻丙氨酸、苯丙氨酸、甲硫氨酸和甘氨酸含量,但随Cd处理浓度增加到Cd100时,Nipponbare水稻苯丙氨酸、甲硫氨酸以及Lemont水稻丙氨酸、甲硫氨酸和甘氨酸未发生显著性变化;对于组氨酸,仅当浓度为Cd100时才发生显著增加,而苏氨酸在任何Cd处理水平下都未发生显著改变。对比2种水稻在3种Cd处理水平下游离氨基酸含量,结果显示,除苏氨酸外,Nipponbare水稻游离氨基酸含量都大于Lemont水稻。可见,Cd胁迫下,2种水稻细胞通过积累游离氨基酸含量增强其抗性,而且不同水稻品种之间存在一定的基因型差异。

表3 不同Cd处理下水稻叶片游离氨酸含量
3 讨论
3.1 水稻叶片光合色素及Cd亚细胞分布对Cd胁迫的响应
光合作用是植物物质合成的主要途径,而光合色素直接影响光合作用的强度。本研究发现,Cd胁迫导致叶绿素和类胡萝卜素含量下降,低浓度Cd5时表现不太显著,但高浓度Cd50和Cd100时表现显著,表明Cd胁迫导致水稻叶绿素和类胡萝卜素合成受阻,而且随着Cd处理浓度增加,光合色素下降越明显,说明Cd对色素合成的抑制存在剂量效应,该生理变化模式与王霞等[7]发现的研究结果相一致。光合色素合成主要由叶绿体结构和相关酶活性调节,本研究高浓度Cd胁迫下,Nipponbare水稻叶绿素和类胡萝卜素下降幅度大于Lemont水稻,可能是高浓度Cd对Nipponbare水稻叶绿体结构和相关酶活性影响更大,促使色素合成受阻,甚至发生分解。水稻对Cd的抗性不仅与其各器官对Cd的吸收与转运有关[8],而且还与Cd在组织内的亚细胞分布紧密联系[9]。本研究Cd亚细胞组织分布结果显示,各组织Cd含量随Cd处理浓度增加而显著增加,其中Lemont水稻各组织Cd小于Nipponbare水稻,表明水稻组织中Cd在亚细胞内的分布因水稻品种不同而表现差异。此外,所有组织中Cd,绝大部分Cd分布在细胞壁,其中Nipponbare占比最高达88.59%,Lemont占比最高达94.17%,说明Lemont细胞壁对Cd有更强的结合作用,减少Cd2+进入细胞内引起Cd毒害。该结果与Cd胁迫对棉花和碱蓬等[10-11]物种的研究结果一致,说明细胞壁固定Cd是植物抗Cd毒害的重要机理之一。
3.2 水稻叶片游离氨基酸含量对Cd胁迫的响应
氨基酸作为蛋白质组成原料,不仅对适宜环境下植物生长发育具有重要的作用,而且也能在植物逆境响应过程中发挥积极作用。本研究发现不同种类间游离氨基酸含量变化存在较大差异,其中高浓度Cd50和Cd100均提高了两种水稻脯氨酸、半胱氨酸、谷氨酸、丙氨酸、苯丙氨酸、甲硫氨酸和甘氨酸含量,而且Nipponbare水稻显著高于Lemont,表明Cd胁迫可引起抗逆相关的游离氨基酸的合成,Nipponbare水稻表现更为明显。张金彪等[12]研究显示,Cd胁迫引起草莓游离脯氨酸的增加;不仅如此,重金属胁迫导致植物游离脯氨酸的积累在较多植物中普遍存在,游离脯氨酸的积累有助于调节细胞渗透势,维持细胞结构,本研究水稻在重金属Cd胁迫下游离脯氨酸含量增加是水稻应对逆境胁迫的一种生理适应。半胱氨酸、谷氨酸和甘氨酸是合成谷胱甘肽的重要前体物质,而谷胱甘肽是植物细胞内含巯基的一种低分子肽,不仅是一种重要的抗氧化物质,而且能通过巯基与Cd离子结合,缓解Cd产生的毒害[13]。本研究Cd胁迫下半胱氨酸、谷氨酸和甘氨酸含量增加是机体运用谷胱甘肽代谢来增强水稻Cd抗性的策略之一。不仅如此,含硫化合物,如甲硫氨酸等,均可通过其氨基酸代谢调控参与缓解Cd对植株产生的毒害[14]。近年的研究表明,游离氨基酸越来越多地参与植物抗重金属胁迫的过程,游离氨基酸中的功能团(羧基和氨基等)可通过螯合作用螯合有毒的重金属离子,降低重金属对植物产生的毒害,起到解毒作用[15]。
综上所述,2种水稻通过调整体内光合色素生物合成、游离氨基酸合成及Cd亚细胞分布响应Cd胁迫,而且不同水稻品种间在Cd胁迫生理响应中存在显著的基因型差异,该差异可视为水稻对Cd毒害的一种抗逆适应表现。
