Environmental hypoxia induces apoptosis in large yellow croaker Larimichthys crocea via both intrinsic and extrinsic pathways*
2023-12-23ShengyuLUOChengLIUXinmingGAOJingqianWANGYiboZHANGJieDINGCongcongHOUJunquanZHUBaoLOUWeiliangSHENXiongfeiWUChundanZHANG
Shengyu LUO, Cheng LIU, Xinming GAO, Jingqian WANG, Yibo ZHANG,Jie DING, Congcong HOU, Junquan ZHU,**, Bao LOU, Weiliang SHEN,Xiongfei WU, Chundan ZHANG
1 Key Laboratory of Applied Marine Biotechnology by the Ministry of Education, School of Marine Sciences, Ningbo University, Ningbo 315211, China
2 State Key Laboratory for Managing Biotic and Chemical Threats to the Quality and Safety of Agro-products, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China
3 State Key Laboratory of Large Yellow Croaker Breeding, Ningbo Academy of Oceanology and Fishery, Ningbo 315012, China
Abstract Hypoxia has become an unfavorable factor affecting the sustainable development of the large yellow croaker Larimichthys crocea, an economically important mariculture fish in China.Apoptosis is a consequence of hypoxia on fish.However, the effects of hypoxia stress on apoptosis in L.crocea remain largely unknown.We investigated the effect of environmental hypoxia on apoptosis in L.crocea.Results show that hypoxia induced apoptosis in L.crocea both in vivo and in vitro.The mitochondrial membrane potential was significantly reduced in large yellow croaker fry (LYCF) cells.The expression levels of Bcell lymphoma/leukemia-2 (Bcl-2) mRNA and protein were also significantly decreased in the liver and LYCF cells during 96 h and 48 h of hypoxia stress, respectively, whereas the expression level of Bcl-2 associated X (Bax) mRNA, Casp3 mRNA, and activity of caspase-3/7/9 were significantly increased,indicating that hypoxia induced caspase-dependent intrinsic apoptosis in L.crocea.The expression level of the apoptosis-inducing factor (AIF) protein was significantly increased in the liver and LYCF cells.The level of AIF protein was significantly decreased in the cytoplasm but increased in the nuclei of L.crocea,demonstrating that hypoxia induced the AIF-mediated caspase-independent intrinsic apoptosis.In addition, the activity of caspase-8 was significantly increased, indicating that hypoxia stress induced extrinsic apoptosis in L.crocea.Therefore, hypoxia induced apoptosis in L.crocea through both the intrinsic and extrinsic pathways.The present study accumulated basic biological information to help elucidate the mechanism of hypoxia response in marine fish.
Keyword: hypoxia; large yellow croaker; mitochondrial apoptosis pathway; death receptor apoptosis pathway
1 INTRODUCTION
Dissolved oxygen (DO), i.e., molecular oxygen dissolved in aquatic environments, is the primary oxygen source for various aquatic organisms.The diffusion rate of oxygen in water is only 1/10 000 of air, and the oxygen content in the same volume of water at the same specific pressure is only 1/30 of air (Vuori et al., 2004; Sun et al., 2011).Unlike the terrestrial environment, hypoxia with DO<2 mg/L occurs frequently in aquatic environments (Diaz,2001).It is worrying that the frequency and severity of hypoxia in the marine environment is expected to continue to increase due to climate change,environmental pollution, and elevated density of aquatic organisms (Breitburg et al., 2018).Hypoxia can distort the homeostasis of the internal environment of fish, leading to reduced feed intake, growth rate and reproduction, and even death (Martínez et al.,2011; Schulte, 2014).
Under normal physiological conditions, apoptosis can remove damaged cells from the organism to maintain the homeostasis of the internal environment.In addition, when an organism is under excessive stress or pathological conditions, pro-apoptotic factors are overactivated, triggering excessive apoptosis or even secondary necrosis, eventually leading to damage and the death of the organism (Ren et al., 2006; Wang et al., 2006).Apoptosis may be triggered by numerous stressors, such as hypoxia, nutrient deprivation, high or low temperature, high or low pH, changes in osmotic pressure, and protein misfolding (Grilo and Mantalaris, 2019).In mammals, intrinsic (also known as the mitochondrial pathway) and extrinsic(also known as the death receptor pathway) pathways are the two important pathways of apoptosis induced by hypoxia stress (Pan et al., 2014; Lohberger et al.,2016).For instance, under hypoxia stress in cells,electrons in the mitochondrial electron transport chain (ETC) have been shown to escape before their transfer to complex IV and react with O2to generate large amounts of reactive oxygen species (ROS)(Halliwell, 1992; Semenza, 2011), which lead to DNA damage (Hammond et al., 2002), altered mitochondrial membrane permeability (Crompton,1999; Hausenloy et al., 2003), and the release of apoptotic proteins such as cytochrome C (CytC).It is worth noting that aquatic environments are more susceptible to hypoxia than terrestrial environments.The relevance of apoptosis to hypoxia response in aquatic animals has received much attention, and studies on the effects of hypoxia stress on apoptosis in fish have been reported inHypophthalmichthys molitrix(Zhao et al., 2016; Ding et al., 2018),Danio rerio(Li et al., 2017),Megalobramaamblycephala(Wu et al., 2016),Mudskipperboleophthalmus(Ren et al., 2018),Micropterussalmoides(Zhao et al.,2020),Micropogoniasundulatus(Ondricek and Thomas,2018),Oryziasmelastigma(Tse et al., 2015), andAcipensershrenckii(Lu et al., 2005).
Apoptosis is mainly mediated by the mitochondrial pathway and death receptor pathway.The mitochondrial pathway has been reported to be predominantly regulated by the B-cell lymphoma/leukemia-2 (Bcl-2) protein family (Chao and Korsmeyer, 1998).In normal cells, members of the Bcl-2 anti-apoptotic protein have been found to antagonize the proapoptotic effects of Bcl-2 associated X (Bax) and Bcl-2 antagonist/killer (Bak).In intrinsic apoptotic cells, the transcription and translation of members of the BH3-only subfamily have been shown to both enhance and inhibit the anti-apoptotic function of Bcl-2 and indirectly activate Bak and Bax (Youle and Strasser, 2008), which induce a decrease in mitochondrial membrane potential (MMP) and alterations in outer mitochondrial membrane permeability(Knudson et al., 1995; Wei et al., 2001; Reed, 2006).These changes are known to lead to the translocation of CytC from the mitochondria to the cytoplasm (Liu et al., 1996; Ow et al., 2008), where it binds to apoptotic protease activating factor-1 (Apaf1) and recruits procaspase-9 to form the apoptosome (Yuan and Akey, 2013).Subsequently, complex-bound procaspase-9 is cleaved into active caspase-9 (Casp9),activating in turn downstream caspase-3 (Casp3) and caspase-7 (Casp7), which can act as apoptotic“executors”, cleaving thousands of cellular substrates(Acehan et al., 2002).In addition, following the altered mitochondrial membrane permeability, the apoptosis-inducing factor (AIF) translocates from the mitochondria to the cytoplasm and enters the nucleus to execute caspase-independent apoptotic programs (Susin et al., 1999).The death receptor pathway is known to be activated by death ligands such as Fas ligand (FasL), tumor necrosis factor-α(Tnf-α), and tumor necrosis factor (TNF) superfamily member 10 (Tnfsf10, also known as TRAIL), which bind to specific receptors on the cell surface to generate the death-inducing signaling complex(DISC), which cleaves procaspase-8 into active caspase-8 (Casp8) (Ashkenazi and Dixit, 1998).Consequently, activated Casp8 cleave procaspase-3 to its active Casp3 form that performs the apoptotic program (Tummers and Green, 2017).Thus, the intrinsic and extrinsic pathways of apoptosis are interlinked and work together to induce apoptosis.
The large yellow croakerLarimichthyscroceais an economically important fish distributed along the southeast coast of China.However, few studies have investigated the effects of stress conditions on apoptosis inL.crocea.Only Wang et al.(2020b)reported the effects of treatment with hydrogen peroxide (H2O2) on oxidative stress and apoptosis in large yellow croaker head kidney cells.Studies on the effects of hypoxia stress on apoptosis inL.croceahave not been reported.Here, we examined for the first time the effect of hypoxia stress on apoptosis inL.croceaboth in vivo and in vitro to accumulate basic biological information that would facilitate the elucidation of the mechanism of hypoxia response in fish.
2 MATERIAL AND METHOD
2.1 Fish experiments and sample collection
Large yellow croakers (length 15.90±1.52 cm,body weight 63.61±6.63 g) were commercially obtained from Fufa Aquatic Products Co., Ltd.(Ningde, China).The fish were domesticated in aerated natural seawater (DO 7.8±0.5 mg/L; salinity 29; temperature 22±0.5 °C; pH 8.1) for two weeks prior to the experiment.A total of 240 fish were randomly divided into six tanks (800 L each).Three of them were used as the hypoxia groups, whereas the remaining three tanks were used as the normoxia group.For the hypoxia group, the DO in each tank was lowered to 2.0 mg/L by infusing nitrogen within 10 min of beginning the experiment and then maintained at 2.0±0.1 mg/L for 96 h.For the normoxia group, treatment conditions were in agreement with the domestication conditions.The HACH DO probe system (HACH LDO II, HACH, Loveland, CO,USA) was used to monitor and maintain DO in realtime.The samples were collected at 0, 3, 6, 12, 24,48, and 96 h after starting the experiment.At each time point, three fish were collected from each tank and sacrificed.Dissected liver samples were stored at -80 °C or fixed with 4% paraformaldehyde (PFA)solution.
2.2 Cell culture and hypoxia challenge
The large yellow croaker fry (LYCF) cell line was kindly provided by Dr.Youhua HUANG from the South China Agricultural University (Guangzhou,China).LYCF cells were cultured in Leibovitz’s-15 Medium (Gibco, Grand Island, NY, USA)supplemented with 10% (v/v) fetal bovine serum(Gibco) and 200 µg/mL of penicillin-streptomycin(Gibco).For the normoxia groups, cells were cultured at 27 °C in an incubator.For the hypoxia groups, LYCF cells were cultured under hypoxia conditions at 1% O2and 99% N2in a MIC-101 modular incubator (Billups Rothenberg Inc.,Del Mar, CA, USA) for 0, 3, 6, 12, 24, and 48 h.
2.3 Detection of apoptotic rates
The annexin V-FITC/PI apoptosis detection kit(Beyotime, Shanghai, China) was used to stain LYCF cells according to the manufacturer’s instructions.Stained cells were immediately examined and photographed using a confocal laser microscope(LSM880, Carl Zeiss Meditec, Jena, Germany) or subjected to flow cytometry (Becton Dickinson, San Jose, CA, USA) to detect the apoptotic rates.Acquired data were analyzed using the FlowJo 10 software (Treestar, Ashland, USA).
2.4 TdT mediated dUTP nick end labeling(TUNEL) immunofluorescence staining
The one-step TUNEL apoptosis assay kit(Beyotime) was used to perform TUNEL immunofluorescence staining on frozen sectioned liver samples and cells seeded in confocal dishes according to the manufacturer’s instructions.Immunostaining images were obtained using a laser confocal microscope (LSM880, Carl Zeiss, Jena,Germany).
2.5 Transmission electron microscopy (TEM)
Preparation and staining of ultrathin sections of LYCF cells after 24, 48, and 72 h of hypoxia were performed according to the method described by Quignard et al.(2012).The sections were examined and photographed using a transmission electron microscope (TEM; H-7650; Hitachi Medical Co.,Chiba, Japan).
2.6 Detection of mitochondrial membrane potential
The MMP of LYCF cells was assessed using the potentiometric dye tetramethyl rhodamine methyl ester (TMRM; MedChem Express, Shanghai, China)at a final concentration of 0.5 μmol/L for 20 min at 27 °C.
2.7 Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
Expression patterns ofBax,Bcl-2, andCasp3were assessed with RT-qPCR using a LightCycler 480 instrument (Roche Diagnostics, Penzberg, Germany).The primers used for RT-qPCR are listed in Table 1.cDNA synthesis and RT-qPCR analysis were performed as previously described (Luo et al., 2019).
2.8 Detection of cleavage activity of caspase-3/7
The cleavage activity of Casp3/7 in LYCF cells was determined using a GreenNucTMCaspase-3/7 assay kit for live cells (Beyotime) according to the manufacturer’s instructions.
2.9 Measurement of activities of caspase-3, 8, and 9
The activities of Casp3, Casp8, and Casp9 were measured both in vivo and in vitro using Casp3,Casp8, and Casp9 activity assay kits (Beyotime),respectively, according to the manufacturers’instructions.

Table 1 Primers used for RT-qPCR analysis of Bax, Bcl-2, Casp3, and β-actin
2.10 Western blot
Whole tissue or cell lysates were prepared using RIPA buffer (Beyotime) containing the protease inhibitors phenylmethanesulfonyl fluoride (PMSF;Beyotime).Protein concentration was measured using a bicinchoninic acid protein assay kit (Cwbio,Beijing, China).After boiling, the supernatants were subjected to sodium dodecyl sulfate polyAcrylamide gel electrophoresis (20-μg protein per lane) and transferred to polyvinylidene fluoride membranes.After blocking with 5% non-fat milk, membranes were successively incubated with primary and horseradish peroxidase-conjugated secondary antibodies before visualizing bands using a chemiluminescence image analysis system (Tanon 5200, Tanon,Shanghai, China).
2.11 Nucleoplasm distribution of apoptosisinducing factor
The nuclear and cytoplasmic protein extraction kit (Beyotime) was used to isolate nuclear and cytoplasmic proteins according to the manufacturer’s instructions.Extracted nuclear and cytoplasmic proteins were subjected to western blot analysis.For the subcellular localization of AIF, LYCF cells seeded in confocal dishes were washed with phosphate buffer saline (PBS) twice and fixed with 4% PFA for 30 min at 25 °C.After washing with PBS and permeabilization with phosphate-buffered saline with 0.3% Triton X-100-containing buffered saline (PBST), sections were blocked with 5% FBS in PBS and incubated with rabbit anti-human AIF polyclonal antibody (1꞉100; Beyotime) overnight at 4 °C.The next day, sections were incubated with Alexa Fluor 555 labeled donkey anti-rabbit IgG(1꞉500; Beyotime) at 37 °C for 1 h.Nuclei were counterstained with 2-(4-Amidinophenyl)-6-indolecarbamidine dihydrochloride (DAPI, Beyotime).Stained sections were imaged using a confocal laser microscope (LSM880, Carl Zeiss, Jena, Germany).
2.12 Statistical analysis
Data are expressed as mean±standard error of the means.Statistical analyses were carried out with SPSS software, version 21.0 (IBM, Armonk, NY,USA).Significant differences between 2 groups were determined using the two-tailed independent samplest-test.Significant differences among 3 or more groups were determined using one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test.
3 RESULT
3.1 Change in the apoptotic rate of LYCF cells under hypoxia stress
After 24 h of hypoxia stress, LYCF cells were double-stained with annexin V-FITC/PI.We found that the positive signal of annexin V and PI in the hypoxia group was significantly enhanced compared with that in the normoxia group (Fig.1a).In addition,flow cytometry analysis revealed that the apoptotic rate of LYCF cells in the normoxia group was 8.99%,whereas that of LYCF cells in the hypoxia group was significantly increased to 35.47% (Fig.1b–c).
3.2 Change in late-phase apoptotic levels in the liver and LYCF cells after hypoxia stress
We accordingly observed that after 96 h of hypoxia stress, the TUNEL-positive signal (red)intensity of hepatocyte nuclei in the hypoxia group was significantly higher than that in the normoxia group (Fig.2a).Likewise, after 24 h of hypoxia stress,the TUNEL-positive signal intensity of the nuclei of LYCF cells in the hypoxia group was significantly higher than that in the normoxia group (Fig.2b).

Fig.1 Change in the apoptotic rate of LYCF cells after 24 h of hypoxia stress

Fig.2 Changes in the TUNEL staining in the liver of Larimichthys crocea and LYCF cells after hypoxia stress
3.3 Ultrastructural alteration of LYCF cells under hypoxia stress
The ultrastructure of LYCF cells in the normoxia group is shown in Fig.3a–b.We noticed that LYCF cells were oval in shape, with abundant mitochondria in the cytoplasm; mitochondrial cristae were clearly visible, while the nucleus was oval, containing a nucleolus in its center.After 24 h of hypoxia stress,we found that the nuclear membrane of LYCF cells was sunken inward, and some mitochondrial cristae in the cytoplasm were reduced or disappeared(Fig.3c–d).The ultrastructure of LYCF cells after 48 h and 72 h of hypoxia stress is shown in Fig.3e,f, & g–h.We specifically observed that the plasma membrane of LYCF cells was wrinkled.Compared with the group exposed to 24 h of hypoxia stress,the number of mitochondria with disappeared cristae was increased, and the inward depression of the nuclear membrane was more obvious in cells exposed to 48 h and 72 h of hypoxia stress.
3.4 Change in the mitochondrial membrane potential of LYCF cells after hypoxia stress
We found that after 24 h of hypoxia stress, the MMP (red fluorescence intensity) of the hypoxia group was significantly lower than that of the normoxia group (Fig.4a).In addition, our flow cytometry analysis revealed that the relative mean fluorescence intensity (MFI) of TMRM in LYCF cells was significantly decreased after 24 h of hypoxia stress (Fig.4b), indicating a significant decrease in the MMP in LYCF cells.
3.5 The mRNA expression pattern of Bax, Bcl-2, and Casp3 in the liver and LYCF cells under hypoxia stress
We noticed that the level of expression ofBaxmRNA in the liver of the large yellow croaker was significantly increased under hypoxia stress (Fig.5a).Meanwhile, the level of expression ofBcl-2mRNA was significantly increased in the first 6 h of hypoxia stress and then rapidly decreased to a significantly lower level than that in the normoxia group (Fig.5c).Correspondingly, we found that theBax/Bcl-2mRNA ratio showed an overall upward trend (Fig.5e).We observed a similar trend in LYCF cells under hypoxia stress.The level of expression ofBaxmRNA was significantly increased (Fig.5b),whereas that ofBcl-2mRNA was rapidly decreased to a significantly lower level than that in the normoxia group after an initial significant increase in the first 6 h of hypoxia stress (Fig.5d), with theBax/Bcl-2mRNA ratio showing an overall upward trend (Fig.5f).We also found that the expression ofCasp3mRNA was significantly increased in the liver and LYCF cells under hypoxia stress (Fig.5g–h).
3.6 Change of Casp3/7 cleavage activity in living LYCF cells under hypoxia stress
We found that the cleavage activity (green fluorescence intensity) of Casp3/7 in LYCF cells after 24 h of hypoxia stress was significantly higher than that in the normoxia group (Fig.6a).In addition,our flow cytometry analysis further showed that the cleavage activity of Casp3/7 (the relative MFI of GreenNucTM) was increased 2.24 folds in LYCF cells after 24 h of hypoxia stress compared with that in the normoxia group (Fig.6b).

Fig.3 Ultrastructure of LYCF cells exposed to hypoxia

Fig.4 Change of MMP in LYCF cells after 24 h of hypoxia stress
3.7 Change in the activity of Casp3/8/9 in the liver and LYCF cells under hypoxia stress
Similar to our previous findings, we observed that the activity of Casp3, Casp8, and Casp9 was gradually increased in the liver during 96 h of hypoxia stress (Fig.6c–e).Similarly, we detected that the activity of Casp3, Casp8, and Casp9 was gradually increased in LYCF cells during 48 h of hypoxia stress (Fig.6f–h).
3.8 Change in the protein expression levels of Bcl-2 and AIF in the liver and LYCF cells under hypoxia stress
Change in the levels of expression of the Bcl-2 and AIF proteins in the liver ofL.croceaafter hypoxia stress for 0, 3, 6, 12, 24, 48, and 96 h are shown in Fig.7a, c, & e.We found that the expression level of the Bcl-2 protein showed an overall downward trend (Fig.7a & c), whereas that of the AIF protein showed an overall upward trend (Fig.7a & e)in the liver ofL.croceaafter 96 h of hypoxia stress.
In addition, change in the levels of expression of the Bcl-2 and AIF proteins in LYCF cells after hypoxia stress for 0, 3, 6, 12, 24, and 48 h are shown in Fig.7a, d, & f.We specifically noticed that prolongation of hypoxia stress resulted in an overall downward trend in the level of expression of the Bcl-2 protein (Fig.7b & d), whereas led to an overall upward trend in the level of expression of the AIF protein in LYCF cells (Fig.7b & f).
3.9 Change in the nucleoplasmic distribution of AIF protein in the liver and LYCF cells under hypoxia stress
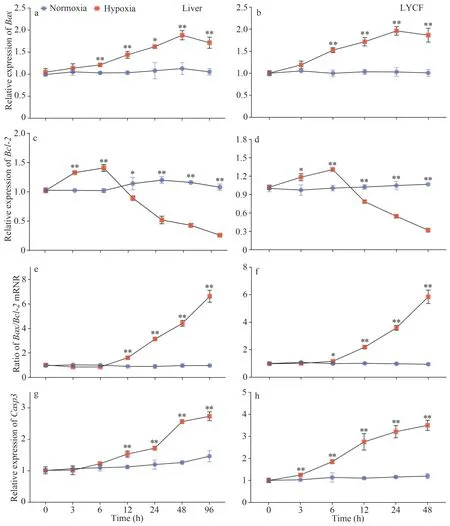
Fig.5 Variations in the expression of Bax, Bcl-2, and Casp3 mRNA in the liver of Larimichthys crocea and LYCF cells under hypoxia stress
Change in the expression levels of the AIF protein in the cytoplasm and nucleus of the liver after 96 h of hypoxia stress are shown in Fig.8a.Compared with the normoxia group, we noticed that the AIF protein was significantly reduced in the cytoplasm, whereas significantly increased in the nucleus of cells in the hypoxia group.Changes in the expression levels of the AIF protein in the cytoplasm and nucleus of LYCF cells after 24 h of hypoxia stress are shown in Fig.8b.Likewise, we detected that the AIF protein was significantly reduced in the cytoplasm, whereas significantly increased in the nucleus of LYCF cells in the hypoxia group compared with those in the normoxia group.Our immunofluorescence (IF) analysis further revealed the stronger presence of AIF in the nuclei of cells in the hypoxia group (Fig.8c).
4 DISCUSSION

Fig.6 Changes in the activity of caspase-3/7, caspase-3, caspase-8, and caspase-9 in the liver of Larimichthys crocea and LYCF cells under hypoxia stress
A number of studies have investigated the hypoxia-induced apoptosis and its underlying molecular mechanism in higher animals (Green and Reed,1998; Murphy et al., 1999; Kunz et al., 2001; Wang et al., 2020a, 2021).Given the frequent occurrence of marine hypoxia, investigating the effect of hypoxia stress on the apoptosis inL.croceacan provide important basic biological information for elucidating the mechanism of hypoxia response in marine organisms.In this study, we found that environmental hypoxia induced apoptosis both in vivo and in vitro inL.croceathrough the mitochondrial and death receptor pathways.
4.1 Hypoxia stress induced apoptosis both in vivo and in vitro in L.crocea

Fig.7 Expression patterns of Bcl-2 and AIF proteins in the liver of Larimichthys crocea and LYCF cells under hypoxia stress
Hypoxia stress is one of the factors known to induce apoptosis in terrestrial animals (Zhang et al.,2013; Pan et al., 2014; Sendoel and Hengartner,2014; Lohberger et al., 2016).Due to the particularity of their respiratory system and habitat, fish might encounter more frequent hypoxia stresses than terrestrial animals.Therefore, the correlation of apoptosis with the response of fish under hypoxia stress has been of special interest to researchers in recent years.Wu et al.(2016) and Ding et al.(2018)examined the apoptotic levels of cardiomyocytes inM.amblycephalaandH.molitrixunder hypoxia stress using the TUNEL method, and found that hypoxia stress induced significant levels of apoptosis in the cardiomyocytes of both fish species.Likewise,Zhao et al.(2016) examined the liver and brain ofH.molitrixunder hypoxia stress using annexin VFITC/PI flow cytometry and the TUNEL assay, and discovered that hypoxia stress induced apoptosis in both the liver and brain ofH.molitrix.To date, no studies have been reported on the effects of hypoxia stress on apoptosis inL.crocea.In this study, we detected for the first time the apoptotic levels in the liver ofL.croceaand LYCF cells after 24 and 96 h of hypoxia stress, respectively, using annexin VFITC/PI flow cytometry and the TUNEL assay.Our results show that the apoptotic rate of LYCF cells in the hypoxia group was significantly higher than that in the normoxia group.In particular, the number of LYCF cells in late apoptosis was significantly higher in the hypoxia group than that in the normoxia group.Similarly, we found that the number of cells in late apoptosis in the liver of fish in the hypoxia group was significantly higher than that in the normoxia group.These results suggested that hypoxia stress induced apoptosis inL.croceaboth in vivo and in vitro, suggesting that apoptosis might be a common response of animals to hypoxia stress.
In 1972, Kerr et al.(1972) first reported the morphological characteristics of apoptotic cells,including cell shrinkage, intact but wrinkled cell membranes, constricted nuclei, condensed nuclear chromatin around the nuclear membrane, and eventual dissolution of cells into several apoptotic bodies with intact membrane structures.In this study, we evaluated the ultrastructure of LYCF cells after 24, 48,and 72 h of hypoxia stress, respectively.We accordingly observed that compared with the normoxia group, the hypoxia group of LYCF cells exhibited the following features: wrinkling of the plasma membrane, constriction of the nuclear membrane by inward depression, breakage or disappearance of some mitochondrial cristae in the cytoplasm, condensation of nuclear chromatin,fragmentation and aggregation around the nuclear membrane.Concomitantly, we found that mitochondrial cristae were broken, mitochondria were vacuolated,nuclei were deformed, and the chromatin was concentrated and fragmented around the nuclear membrane in liver tissue cells after 96 h of hypoxia stress (Luo et al., 2021).These findings further suggested that hypoxia stress induced apoptosis in the liver ofL.croceaand LYCF cells.In accordance to our findings, Sollid et al.(2003) observed cell shrinkage and chromatin fragmentation around the nuclear membrane in the gill tissue ofCarassius carassiusafter hypoxia stress.

Fig.8 Changes in the levels of AIF protein in the cytoplasm and nuclei in the liver of Larimichthys crocea and LYCF cells under hypoxia stress
4.2 Hypoxia stress induced apoptosis in L.crocea through the mitochondrial pathway
The mitochondrial pathway is one of the major pathways of hypoxia-induced apoptosis (Pan et al.,2014; Lohberger et al., 2016).The oligomerization of Bax and Bak in the mitochondria-mediated apoptotic pathway has been reported to induce a decrease in MMP and an increase in outer mitochondrial membrane permeability (Knudson et al., 1995; Wei et al., 2001; Reed, 2006).In this study, we identified that hypoxia stress caused a decrease in MMP in LYCF cells.Combined with the observed in vivo and in vitro induced apoptosis of cells, we postulated that the decrease in MMP was one of the reasons for the occurrence of apoptosis under hypoxia stress.The hypoxia stress-induced reduction of cellular MMP has also been reported in human pulmonary artery smooth muscle cells(hPASMCs) (Hu et al., 2010), human colorectal cancer cells (HCT116) (Dong et al., 2019),hepatocellular carcinoma cells (HCC) (Cai et al.,2019), and human retinal pigment epithelial cells(Feng et al., 2020).The Bcl-2 protein is mainly located in the mitochondrial membrane, playing a role in antagonizing Bax.The increased expression of Bcl-2 is known to results in the polymerization of a large amount of Bcl-2 with Bax toward the formation of a heterodimer, thus preventing the oligomeric activation of Bax and Bak.In contrast,and despite the formation of a heterodimer with Bcl-2, the increased expression of Bax results in the excess Bax continuing to play its pro-apoptotic role with the oligomeric activation of Bak.Therefore, the ratio of the expression of theBax/Bcl-2genes has been used to indicate the level of intrinsic apoptosis(Tsukahara et al., 2006).In this study, we noticed the upregulated expression ofBaxin the liver ofL.croceaand LYCF cells under hypoxia stress.Whereas,the expression ofBcl-2was shown to be downregulated, with theBax/Bcl-2ratio being accordingly significantly increased.Studies on the hypoxia stress-induced elevatedBax/Bcl-2mRNA ratios in fish have also been reported inM.amblycephala(Wu et al., 2016),Ictaluruspunctatus(Yuan et al., 2016), andH.molitrix(Ding et al.,2018).In addition, the present study revealed the significantly reduced expression ofBcl-2in the liver ofL.croceaand LYCF cells under hypoxia stress.Similar results were observed in studies of primary cultured rat cortical neurons (Tamatani et al., 1998)and rat cardiomyocytes (Li et al., 2011).The enhanced mitochondrial outer membrane permeability has been shown to lead to the release of CytC from the mitochondria into the cytosol (Liu et al., 1996;Ow et al., 2008) to bind to Apaf1 and activate Caspase-9 (Yuan and Akey, 2013).Activated caspase-9 cleaves procaspase-3 and procaspase-7 into active caspase-3 and caspase-7, respectively,which in turn triggers apoptosis (Acehan et al.,2002).Here, we found that the activity of caspase-9 and caspase-3/7 was significantly elevated in the liver ofL.croceaand LYCF cells under hypoxia stress.Similarly, the activity of caspase-9 was reported to be significantly increased in the cytoplasm of human corneal epithelial cells (Kurpakus-Wheater et al., 2003), neonatal piglet cerebral cortex cells(Mishra and Delivoria-Papadopoulos, 2006), and mitochondria of neonatal piglet cerebral cortex(Mishra et al., 2006).The hypoxia stress-induced activation of caspase-3/7 has also been reported inCarassius auratus(Poli et al., 2003),D.rerio(Williams et al., 2017) andM.undulatus(Ondricek and Thomas, 2018).The above findings suggested that hypoxia stress induces the mitochondrial pathway-mediated caspase-dependent apoptosis inL.crocea.
In addition to CytC, the AIF pro-apoptotic protein has also been shown to translocate from the mitochondria to the cytoplasm and then to the nucleus following the altered mitochondrial membrane permeability, promoting the fragmentation of genomic DNA for the execution of a caspase-independent apoptotic program (Susin et al., 1999).Studies have shown that hypoxia can induce an increase in the level of expression of the AIF protein (Goel et al.,2010; Kim et al., 2017).In this study, the expression levels of the AIF protein were significantly increased in the liver ofL.croceaand LYCF cells under hypoxia stress; moreover, hypoxia stress induced the nuclear translocation of the AIF protein.Studies on the hypoxia stress-induced nuclear translocation of AIF have also been reported in higher mammals.For example, Brukamp et al.(2007) found that the nuclear translocation of the AIF protein was significantly enhanced in mouse podocytes under hypoxia stress.Likewise, Kim et al.(2010) found that the nuclear translocation of the AIF protein was significantly augmented in human lung adenocarcinoma cells(A549) after hypoxic hypoglycemic stress.These investigators suggested that the release of AIF from the cytoplasm into the nucleus for the promotion of DNA fragmentation is one of the pathways through which hypoxia stress induces apoptosis.Therefore,we postulated that hypoxia stress induces the AIF-mediated caspase-independent apoptosis inL.crocea.
4.3 Hypoxia stress induced apoptosis in L.crocea through the death receptor pathway
The extrinsic apoptotic pathway is known to be activated by death ligands such as FasL, TNF-α, and TRAIL, which bind to specific receptors on the cell surface to generate DISC, which cleaves procaspase-8 into active caspase-8 (Ashkenazi and Dixit, 1998).In turn, the activated caspase-8 cleave procaspase-3 to its active caspase-3 form that executes the apoptotic process (Tummers and Green, 2017).The activated caspase-8 has also been shown to cleave Bid into its active truncated Bid form, which rapidly moves to the outer mitochondrial membrane, promoting the oligomerization of Bax and Bak for the opening of the mitochondrial membrane permeability translocation pore and the formation of the pro-apoptotic protein release channel (Cassidy-Stone et al., 2008).Studies on the hypoxia stress-induced enhancement of the activity of caspase-8 have been reported in rat ventricular myocytes (Gurevich et al., 2001), human cardiac myocytes (Chao et al., 2002), neonatal piglet cerebral cortex (Khurana et al., 2002), and oral cancer cells (Nagarajah et al., 2004).In this study,we examined the activity of caspase-8 in the liver ofL.croceaand LYCF cells during 96 and 48 h of hypoxia stress, respectively, and found that hypoxia stress significantly enhanced the activity of caspase-8 both in vivo and in vitro.These findings suggested that hypoxia induced the death receptor pathwaymediated apoptosis inL.crocea.
5 CONCLUSION
In this study, we performed annexin V-FITC/PI,TUNEL, and TEM assays to show that hypoxia stress induced apoptosis inL.croceaboth in vivo and in vitro.Analyses of MMP, of the activity of caspase-3/7/9, of the level of expression of the Bcl-2 protein, of theBax/Bcl-2mRNA ratio, and of the expression ofCasp3mRNA indicated that hypoxia stress induced the mitochondrial pathway-mediated apoptosis inL.crocea.More specifically, analysis of the level of expression of the AIF protein and its nucleoplasmic distribution indicated that hypoxia stress induced the mitochondria/AIF-mediated caspaseindependent apoptosis inL.crocea.Finally, analysis of the activity of caspase-8 revealed that hypoxia stress also induced the extrinsic pathway-mediated apoptosis inL.crocea.In summary, we found that hypoxia stress can induce apoptosis inL.croceathrough both the intrinsic and extrinsic pathways.
6 DATA AVAILABILITY STATEMENT
The data of this study are available from the corresponding author upon reasonable request.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Trends of carbon and nutrient accumulation through time in the Andong salt marsh, Hangzhou Bay, China*
- Physical processes determining the distribution patterns of Nemopilema nomurai in the East China Sea*
- Comparison in structure and predicted function of epiphytic bacteria on Neopyropia yezoensis and Neopyropia katadae*
- Interaction between macroalgae and microplastics: Caulerpa lentillifera and Gracilaria tenuistipitata as microplastic bio-elimination vectors*
- Lake regime shift from submerged macrophyte to phytoplankton affected phosphorus speciation in sediment and eutrophic state in Caohai Lake, Guizhou, China*
- Temporal characteristics of algae-denitrifying bacteria co-occurrence patterns and denitrifier assembly in epiphytic biofilms on submerged macrophytes in Caohai Lake, SW China*