Effects of acute and chronic thermal stress on survival,apoptosis, and transcriptional responses of Scapharca broughtonii*
2023-12-23DeshengZOUWeianCAOGuilongLIUJunhaoNINGXiaLUJinjingWANGMinCHENBoLIUJinshengZHANGChundeWANG1
Desheng ZOU, Weian CAO, Guilong LIU, Junhao NING, Xia LU,Jinjing WANG, Min CHEN, Bo LIU, Jinsheng ZHANG, Chunde WANG1,,
1 Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai 264003, China
2 University of Chinese Academy of Sciences, Beijing 100049, China
3 Yantai Spring-Sea AquaSeed Co., Ltd., Yantai 264006, China
4 Qingdao Agricultural University, Qingdao 266109, China
5 Zhaoyuan Xiadian Agricultural Technology Promotion Station, Yantai 265415, China
Abstract Ocean warming is altering the habitats of marine invertebrates, which has resulted in an increased physiological stress to marine molluscs, especially those intertidal bivalves, such as the ark shell Scapharca broughtonii.We investigated the physiological and transcriptional responses of ark shells to acute and chronic thermal stress results showed that at 33 °C, a significantly higher cumulative mortality(55.7%) occurred under acute thermal stress than chronic thermal stress.The apoptosis rate of hemocytes was sustained at higher levels and the necrosis rate was increased significantly in a time-dependent manner under acute thermal stress.However, under chronic thermal stress, the apoptosis and necrosis rates exhibited similar change trends: a rapid increase followed by a gradual decline and sustained at a relatively high level until the end of the experiment.The expressions of heat shock protein genes (HSP20 and HSP90), apoptosis-related genes (TRAF6, GRP78, NIX, and Casp-3), antioxidative-related genes(GST and MRP) and cellular detoxification-related genes (HbⅡB, NOS-1, HO-1, and ENO-1) were upregulated significantly under both acute and chronic thermal stresses.These results demonstrated that the anti-apoptotic system, antioxidant defense system, cellular detoxification system, and heat shock proteins(HSPs) played vital roles for ark shells in response to thermal stress.As acute thermal stress can result in irreversible damage to marine molluscs, it is thus advised that chronic thermal stress should be used to select thermal-resistant ark shell strains.
Keyword: Scapharca broughtonii; thermal stress; apoptosis; necrosis; gene expression
1 INTRODUCTION
As a result of global warming, the average surface temperature of the global sea has risen by almost 1 °C in the last 40 years.Simulations by the Intergovernmental Panel on Climate Change (IPCC)stated that global average sea surface temperatures could rise as much as 4.8 °C by the end of the 21stcentury if these trends continue (IPCC, 2013).Extreme ocean warming events in the summers of three consecutive years from 2016 to 2018 were reported in the East China Sea and South Yellow Sea (Gao et al., 2020).Marine organisms inhabiting nearshore areas, unlike terrestrial animals, are usually unable to effectively evade from high-temperature environments(Burrows et al., 2019).
Temperature is a prominent environmental stress to marine organisms, especially intertidal marine molluscs, whose survival, growth, development,reproduction, metabolism, immunity, and geographical distribution can be seriously affected.Proper temperature regulation is beneficial to the growth, development, and reproduction of intertidal organisms, as well demonstrated in the snailPlicopurpurapansa(Naegel and López-Rocha, 2007).However, if the threshold temperature is exceeded, it will adversely affect many aspects of life.It has been reported that fluctuations in temperature greatly affected the survival rate ofCrepidulafornicatain Nahant,Massachusetts (Pechenik et al., 2020).To a certain extent, thermal stress tends to cause physiological imbalance in animals, directly stimulating the metabolism of the body and leading to excessive accumulation of reactive oxygen species (ROS)(Lesser, 2006).Consequently, key cellular components(e.g., proteins, DNA, and membrane lipids of the body) are damaged, leading to the disruption of the physiological performance and gene expression in marine organisms (Li et al., 2020; Clark et al.,2021), which triggers a series of cellular defense events (Meng et al., 2014).Studies have shown that temperature change adversely induced body damage by activating the enzyme defense system,antioxidant system, immune system, and apoptotic pathway in marine organisms (Zhou et al., 2019;Ning et al., 2021).Molluscs inhabiting in intertidal waters are often exposed to acute or chronic thermal stresses, and the adaptability of intertidal organisms to climate warming is poor (Stillman, 2003).Therefore,studying the biological responses of these intertidal molluscs to thermal stress may provide insights into the mechanism of adaptation of these intertidal molluscs to high temperatures.
The ark shell,Scapharcabroughtonii, is a large intertidal marine mollusc widely distributed in the Sea of Japan, Bohai Sea, Yellow Sea, and East China Sea.It is highly demanded in the market due to its delicious meat, high protein, and low fat(Wang et al., 2013).In recent years, due to overfishing and habitat destruction, the natural resources ofS.broughtoniihave been dramatically declining.With the global warming, rising of water temperature is also becoming one of the most important factors threatening their survival, especially in hot summers (Ning et al., 2021).Therefore,exploring the apoptosis, physiological properties,and gene expression ofS.broughtoniiunder different thermal stresses will help to comprehensively understand the thermal tolerance of ark shells and provide a scientific basis for the selection of new heatresistant strains.
2 MATERIAL AND METHOD
2.1 Experimental animal
HealthyS.broughtonii(34.30±3.09 mm in shell length, 14.58±4.14 g in whole weight) were obtained from a seafood market in Yantai, China (37°51′N,121°46′E).The ark shells were acclimated in 200-L tanks with filtered seawater (salinity 29±0.5, pH 8.13±0.2) at 14±0.5 °C for one week prior to experimentation.The animals were fed daily withChaetocerosmuelleriand the seawater was changed twice a day during this period.
2.2 Determination of the upper median lethal temperature
After acclimation, 480 healthy animals were randomly divided into eight groups, a control group at 14 °C and seven experimental groups at 17, 20,23, 26, 29, 32, and 35 °C, respectively, with 3 replicates in each group.The water in each group was heated to the designated temperature at a rate of 1 °C/h by a heating rod and maintained at that temperature for 96 h.The ark shells were checked once every 12 h and those did not respond to mechanical touch at the mantle were deemed to dead and selected out.The seawater in the tanks was replaced by clean seawater heated to the same temperature in advance, twice a day.Determination of median lethal temperature (LT50) within 96 h by two-point method (Liu et al., 2007) (Fig.1).
2.3 Experimental design and sample collection
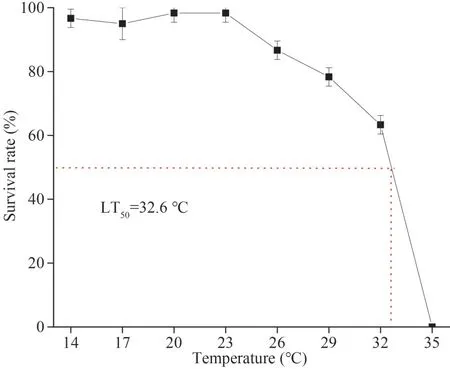
Fig.1 Determination of 96-h LT50
During the experiment, 1 200 animals in healthy condition were randomly chosen and divided equally into four groups, including the acute heating control group (AHCG), the acute heating experimental group (AHEG), the chronic heating control group(CHCG), and the chronic heating experimental group (CHEG).Three replicates were set for each group.The ark shells were checked once every 12 h and those did not respond to mechanical touch at the mantle were deemed to dead and selected out during the experiment.
In the AHEG group, seawater temperature was heated from 14 °C to 33 °C at a rate of 1 °C/h.After the water temperature reached 33 °C, nine animals were randomly selected for sampling at 1, 3, 6, 12,and 24 h (Fig.2a).In the AHCG group, ark shells were cultured at room temperature (14±0.5 °C) and nine animals were randomly collected at 1, 6, and 24 h.Animals were not fed during the experiment.
In the CHEG group, seawater temperature was heated slowly from 14 °C to 33 °C at a rate of 1 °C/d,and nine animals were randomly sampled on Days 1,2, 3, 4, and 7 after the water temperature reached 33 °C (Fig.2b).In the CHCG group, the animals were cultured at room temperature (14±0.5 °C) and nine animals were randomly sampled on Days 1, 3,and 7.During the experiments, animals were fed with concentrated microalgae diet twice a day, and the seawater was changed using preheated seawater 2 h after each feeding.
The sampled ark shells were dissected to collect their hemocytes and gills immediately after sampling.From each animal, 0.5 mL of hemolymph was collected using a sterile syringe filled with 0.5 mL of precooled anticoagulant buffer (336-mmol/L NaCl, 27-mmol/L sodium citrate, 9-mmol/L EDTANa2, 115-mmol/L glucose, pH 7.0).The hemolymphs collected from per animal at the same time point were pooled together and filtered with a 300-mesh sieve.The hemocytes centrifugated at 1 000×gat 4 °C for 5 min were used for measurement of apoptosis rate and necrosis rate immediately.Meanwhile, the gills from 3 animals of the same time point were pooled together, thoroughly washed with phosphate-buffered saline (PBS, pH 7.4), and stored at -80 °C for further analyses.
2.4 Apoptosis and necrosis rate of hemocytes
The hemocyte pellets were washed twice with PBS (pH 7.4), resuspended in 195-µL 1×binding buffer solution buffer, and stained with 10-µL PI dyes and 5-μL annexin V-FITC at 20 °C in dark for 20 min.Analyses of cell suspensions by the flow cytometry (CytoFLEX, USA).The apoptosis rate was expressed as the percentage of the number of PI negative and V-FITC positive cells in the total number of cells.The necrosis rate was represented as the percentage of the number of PI positive and VFITC positive cells in the total number of cells(Schutte et al., 1998).
2.5 RNA extraction and cDNA synthesis
Total RNA was extracted from the gill samples by TRIzol reagent (Invitrogen, USA), genomic DNA was digested with RNase-free DNase I (Vazyme,China).The purity and concentration of the RNA samples were analyzed using Nanodrop 2000 device(Thermo, USA), and RNA integrity was determined by 1% agarose gel electrophoresis.Synthesis of cDNA with the HiScript®III RT Super Mix for qPCR Kit (Vazyme, China) and stored at -20 °C until further analyses.
2.6 Gene expression analysis
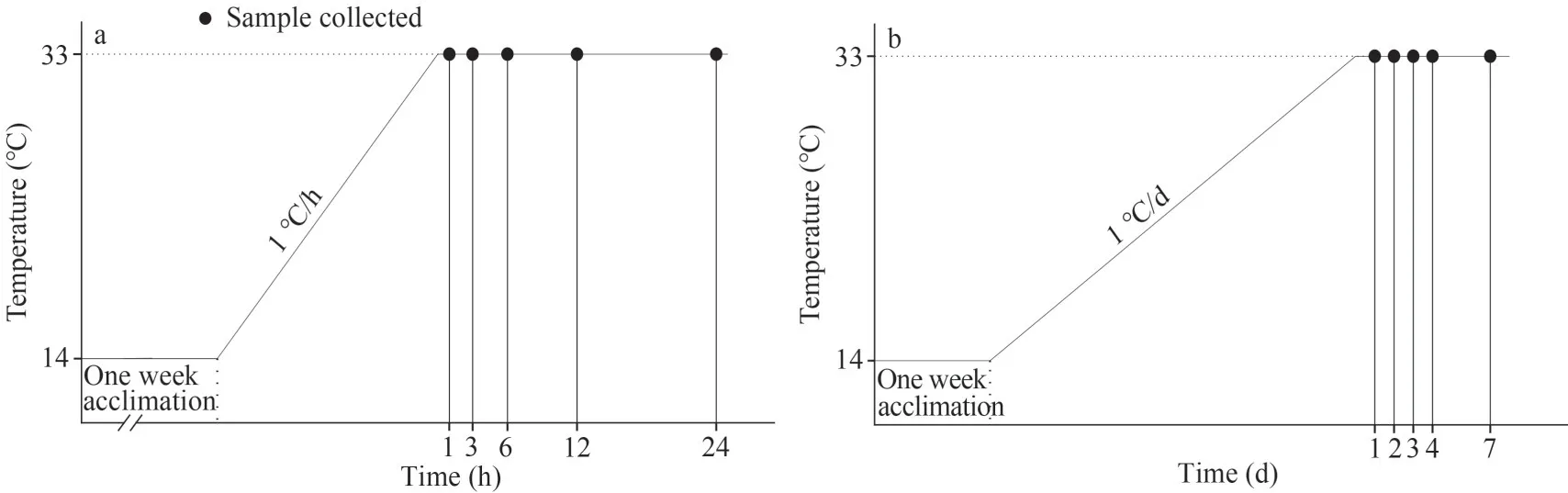
Fig.2 Schematic outlines of the acute (a) and chronic (b) heating treatment experiment
The expression levels of twelve genes, including heat shock proteins (HSP90andHSP20), apoptosisrelated genes (TRAF6,GRP78,NIX, andCasp-3),antioxidative-related genes (GSTandMRP) and immune-related genes (Hb ⅡB,NOS-1,HO-1, andENO-1), were selected to verify their responses to thermal stresses.Primers used for the qRT-PCR analyses are listed in Table 1.18S rRNA was used as the internal reference for the normalization of gene expression.The reactions were performed on the QuantStudioTM5 Real-Time PCR Instrument(Applied Biosystems, USA), and fluorescent dye was SYBR green II.PCR amplifications were performed using a 10-μL reaction mixture containing 0.2 μL of each primer (10 μmol/L), 1 μL of the template cDNA, 3.6-μL ddH2O and 5 μL of 2×Taq Pro Universal SYBR qPCR Master Mix (Vazyme,China).The PCR program was set as follows: 95 °C for 60 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 15 s.The amplification specificity was confirmed by a melting curve analyses.All reactions were performed in triplicates (Ning et al., 2021).The relative expression of the selected genes was analyzed with the 2-ΔΔCtmethod (Livak and Schmittgen,2001).

Table 1 Primers used in this study
2.7 Statistical analysis
SPSS 17.0 software was used for statistical analyses.Significant differences were analyzed using one-way analyses of variance and Duncan’s test.The statistically significant differences were considered asP<0.05 (*) andP<0.01 (**).
3 RESULT
3.1 Determination of 96-h LT50
In this study, the upper LT50ofS.broughtoniiwithin 96 h was determined to be 32.6 °C (Fig.1).
3.2 The cumulative mortality of ark shells during acute and chronic heating treatment
In the AHEG, the cumulative mortality of ark shells increased linearly with the extension of stress time and reached 55.7% within 96 h.However, in the CHEG, the cumulative mortality was much lower than that in the AHEG, with a cumulative mortality rate of only 21.67% within 96 h.There were almost no deaths during the experiment in the AHCG and CHCG (Fig.3).
3.3 The apoptosis and necrosis of hemocytes under acute and chronic heating treatment

Fig.3 Cumulative mortality of ark shells during the acute and chronic heating experiment
In the AHEG, the apoptosis rate of ark shell hemocytes was increased significantly and reached the peak value (18.97%) at 3 h, then decreased slowly and restored to the initial level at 24 h(Fig.4a).However, the necrosis rate of hemocytes showed a linear increase with time and reached the maximum value (11.86%) at 24 h (Fig.4b).However,in the CHEG, the apoptosis and necrosis rate of hemocytes showed a similar change trend: increase significantly and reach the maximum value on Day 1, then a linear decrease from Days 1 to 4 followed by a slight increase on Day 7.Moreover, the apoptosis and necrosis rates of hemocytes in the CHEG group were higher than those in the control group (Fig.4c–d).
3.4 Transcriptional response of ark shell gills to acute heating treatment
During the acute heating treatment, the expressions of heat shock protein genes (HSP20andHSP90),apoptosis-related genes (TRAF6,GRP78,NIX, andCasp-3), antioxidative-related genes (GSTandMRP) and immune-related genes (Hb ⅡB,NOS-1,HO-1, andENO-1) were up-regulated significantly under acute thermal stress, and the peak values were mostly observed at 1 and 12 h.However, the expressions ofHSP90andCasp-3were downregulated manifestly at 24 h (Fig.5).
3.5 Transcriptional response of ark shell gills to chronic heating treatment
During the chronic heating treatment, the expression levels of the twelve selected genes were up-regulated significantly, with the maximum values mainly on Days 3 and 4.However, the expression ofMRPwas down-regulated significantly on Day 1(Fig.6).
4 DISCUSSION
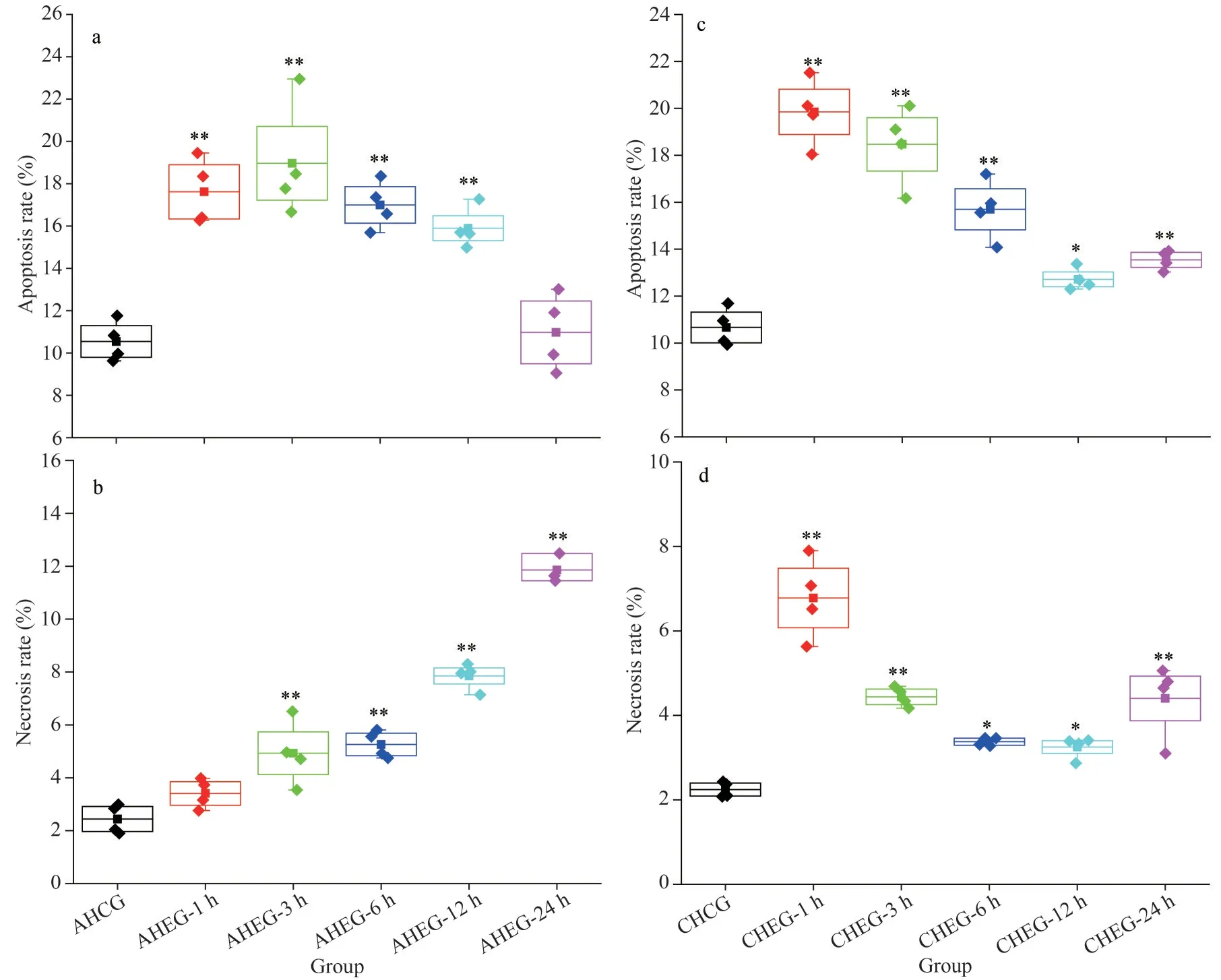
Fig.4 Apoptosis and necrosis rates of ark shell hemocytes under acute and chronic heating stress
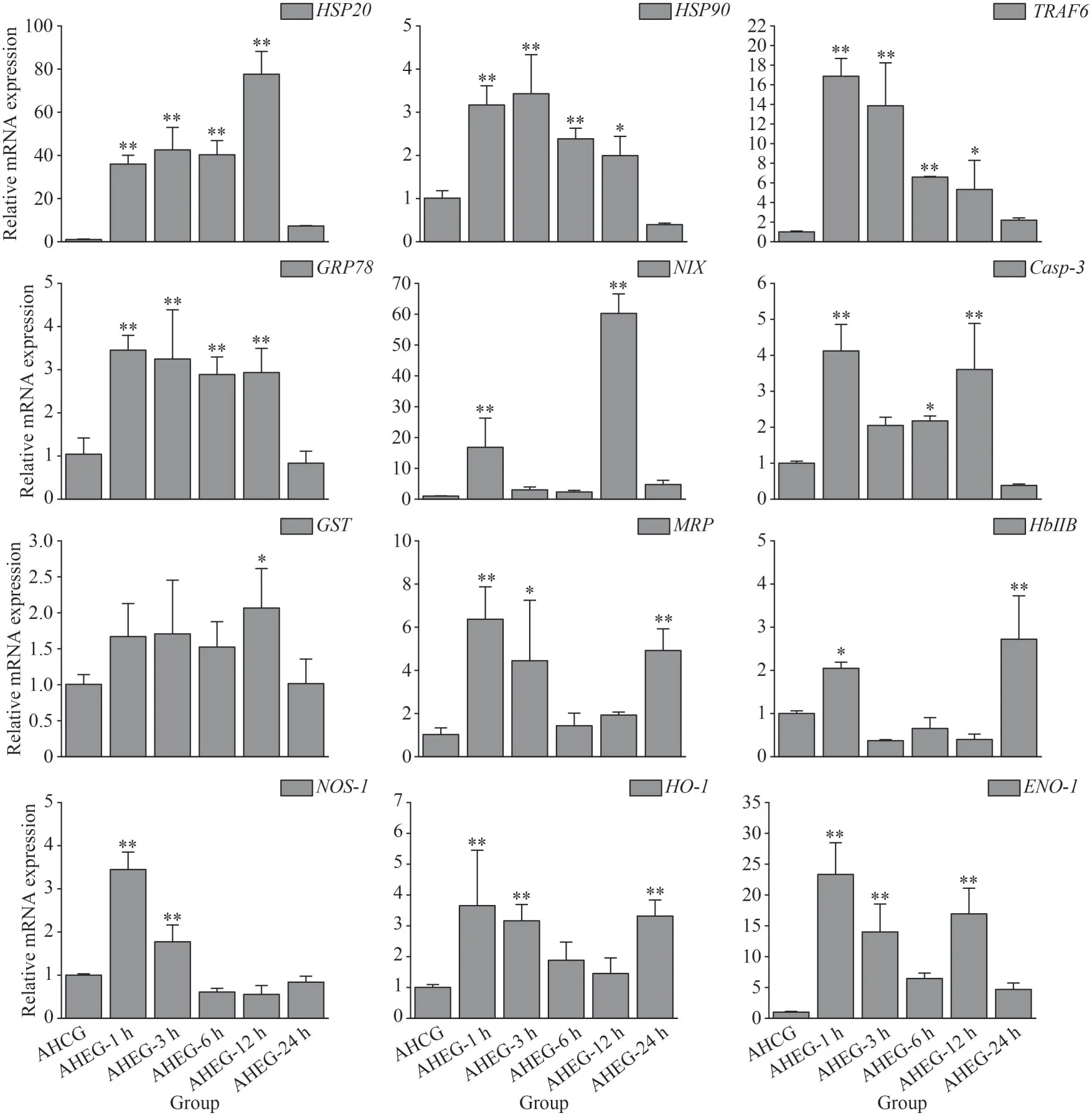
Fig.5 Transcriptional response of ark shell gills to acute thermal stress
In recent years, ocean warming has led to an obvious decline in marine fisheries productivity and sustainable fishing potential, and overfishing has reduced the adaptive capacity of many marine invertebrates to a warmer ocean (Free et al., 2019),which may have contributed to the frequent occurrence of invertebrate deaths.Therefore, it is necessary to investigate the physiological responses and adaptive mechanism of marine invertebrates,especially animals with poor mobility, to thermal stresses.In this study, the upper LT50ofS.broughtoniiwithin 96 h much lower than the maximum temperature in the north inshore of China,especially the intertidal flats where ark shells normally dwell (Ning et al., 2015).Obviously, the ark shells may be subjected to severe heat stresses in hot summers.

Fig.6 Transcriptional response of ark shell gills to chronic thermal stress
It has been confirmed that apoptosis and necrosis are important physiological processes of organisms in response to environmental stress, which could be induced significantly by thermal stress (Cheng et al.,2018; Zhou et al., 2019; Zou et al., 2021).Similarly,in this study, the apoptosis and necrosis rates of hemocytes were also increased significantly when ark shells were subjected to thermal stress.Under acute heat stress, the apoptosis rate remained at a high level and the necrosis rate increased in a timedependent manner, indicating that the apoptosis and necrosis pathways of ark shells has been activated and the stress injury might be irreparable with the prolongation of stress duration.This result is consistent with previously published information inCrassostreagigas(Yang et al., 2017) andS.subcrenata(Zou et al., 2021).However, the difference was that the apoptosis rate and the expression of some related genes in AHEG can return to their original state at 24 h.Related results have also been reported inPatinopectenyessoensis,Litopenaeusvannamei,andHippopotamus, suggesting that the internal environment of these animals has been disturbed(Jiang et al., 2016; Wang et al., 2019; Pan et al.,2022).Moreover, the significant up-regulation of apoptosis-related genes was also in support of the above viewpoint.However, under chronic thermal stress, the apoptosis and necrosis rates of hemocytes showed a similar trend, a significant increase followed by a slow decline, indicating ark shells might gradually build the adaptability to a suboptimal temperature.The manifestly lower mortality of ark shells under chronic thermal stress compared with that under acute thermal stress also supported the above viewpoint.It is speculated that local adaptation and phenotypic plasticity play crucial roles in the modification of thermal tolerance of organisms (Sanford and Kelly, 2011; Yampolsky et al., 2014).
Heat shock proteins (HSPs) are a family of highly conserved proteins that play key roles in rapid responses of organisms to environmental changes, and the expression ofHSPs is often considered as an important indicator to analyze the responses of organisms to temperature stress (Feder and Hofmann, 1999; Srivastava, 2002).Among HSPs, HSP90s are essential chaperone proteins in response to thermal stress in many marine molluscs(Kim et al., 2009; Lim et al., 2016).HSP20s, also named stress induced protein 1, could prevent aggregation of stress-induced proteins and facilitate regeneration of denatured proteins when animals suffer thermal stress (Nover and Scharf, 1997; Ning et al., 2021).In this research, the expression ofHSPs (HSP20andHSP90) was rose remarkably under both acute and chronic heat stresses, which was consistent with our previous results (Ning et al.,2021; Zou et al., 2021).It is suggested that the significant up-regulation ofHSPs would help to strengthen the high temperature tolerance of ark shells.Similar results were also reported in Pacific oysterC.gigas(Lim et al., 2016) and pufferfishTakifuguobscurus(Cheng et al., 2018).
Apoptosis is a key physiological protective mechanism of organisms in response to environmental stresses (Cheng et al., 2018; Zhou et al., 2019).Caspase 3 (Casp-3) is responsible for the cleavage of structural proteins and regulatory proteins participating in apoptosis, which ensures the normal progression of apoptosis (Qin et al., 2020).Nip3-like protein X (NIX) is a key member of the BNIP 3 subfamily, which promotes either apoptotic or nonapoptotic cell death (Chinnadurai et al., 2008).Moreover, the anti-apoptotic pathways in molluscs could be activated to counteract the injuries caused by various stresses (Zhang et al., 2012).For instance, Glucose regulated protein 78 kD (GRP78)could reduce the damage caused by apoptosis by inhibiting the activation ofCasp-7(Reddy et al.,2003).Tumor necrosis factor receptor-associated factor 6 (TRAF6) could inhibit the apoptosis of oyster hemocytes through activation ofPellino(Lin et al., 2020).The up-regulation of apoptosis-related genes (Casp-3,NI X,GRP78, andTRAF6) indicated that the apoptotic and anti-apoptotic systems existed in ark shells and may work simultaneously under heat stress, which is consistent with the previous studies reported inS.subcrenata(Ning et al., 2021;Zou et al., 2021).
As poikilothermal animals, molluscan body temperature changes with the ambient temperature,and thus inevitably affects their metabolism and physiological processes (Konstantinov et al., 2003;Brucet et al., 2012).For instance, reactive oxygen species (ROS) can be produced by respiratory burst of phagocytes under thermal stresses, resulting in severe injury to organisms (Bachère et al., 1995; De La Fuente and Victor, 2000).Moreover, elevated seawater temperature might promote the decomposition of ark shells secretions into toxic substances(Eriksson et al., 2013).It has been reported that glutathione S-transferase (GST) and multidrug resistance protein (MRP) are involved in antioxidant defense and cell detoxification (Hayes and Pulford,1995; Mejdoub et al., 2017; Ning et al., 2021).HbⅡB and Heme oxygenase-1 (HO-1), as the major components of haemoglobin, play important roles in cell detoxification (Geuken et al., 2005; Bao et al.,2013).The significant up-regulation of these genes was observed in this study, suggesting antioxidant defense system provides vital protection for ark shells from irreversible injury caused by thermal stresses.This is consistent with the conclusion found inDicentrarchuslabrax(Hachfi et al., 2012)andS.subcrenata(Zou et al., 2021).Nitric oxide synthases (NOS) are responsible for catalytic production of nitric oxide in mitochondria, which is a vital biological process for maintaining cell metabolism (Haynes et al., 2004).NOS is involved in a variety of functions in invertebrates, and upon stimulation, the expression levels ofNOSin midintestine, hepatopancreas, and hemocytes were significantly increased (Li et al., 2012).In the present study, the expression ofNOS-1showed a similar trend, a significant increase followed by a quick decline, under both acute and chronic thermal stress.Our results also showed that the expression ofEnolase-1(ENO-1) was also up-regulated by thermal stress.ENO-1 is an enzyme of the glycolytic pathway that interacts with plasminogen and suggests ENO-1 as a candidate marker for monitoring health status in teleosts (Santana et al.,2021).The significant up-regulation of α-ENOin kidney and spleen of the rainbow trout was also found when challenged withYersiniaruckeri(Kumar et al., 2018).Therefore, we speculate thatENO-1can also be used as an indicator of invertebrate response to high temperature stimulation.
5 CONCLUSION
In summary, the upper LT50of adult ark shells was determined to be 32.6 °C.The higher mortality in ark shells under acute thermal stress suggested they suffered more injuries and gradually turned irreversible under acute thermal stress.The apoptosis and necrosis rates of ark shell hemocytes were increased significantly under both acute and chronic thermal stresses, and significant up-regulation of apoptosis-related genes was observed herein,suggesting the apoptotic and anti-apoptotic systems were activated in response to thermal stresses.Moreover, the expression of the genes involved in antioxidant defense, cellular detoxification and homeostasis was increased significantly to protect ark shells from oxidative damage.These results may provide new clues for management strategy of ark shell husbandry and further understanding of the tolerance to thermal stresses in ark shells.
6 DATA AVAILABILITY STATEMENT
The data that support the findings of this study are available from the corresponding author upon reasonable request.
7 ACKNOWLEDGMENT
We are grateful to all the laboratory members and anonymous reviewers for their technical advice and helpful discussions.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Trends of carbon and nutrient accumulation through time in the Andong salt marsh, Hangzhou Bay, China*
- Physical processes determining the distribution patterns of Nemopilema nomurai in the East China Sea*
- Comparison in structure and predicted function of epiphytic bacteria on Neopyropia yezoensis and Neopyropia katadae*
- Interaction between macroalgae and microplastics: Caulerpa lentillifera and Gracilaria tenuistipitata as microplastic bio-elimination vectors*
- Lake regime shift from submerged macrophyte to phytoplankton affected phosphorus speciation in sediment and eutrophic state in Caohai Lake, Guizhou, China*
- Temporal characteristics of algae-denitrifying bacteria co-occurrence patterns and denitrifier assembly in epiphytic biofilms on submerged macrophytes in Caohai Lake, SW China*