Diversity and geographical distribution of haptophyte Phaeocystis in the Chinese seas based on metabarcoding analysis*
2023-12-23QingshangSONGYueXUEYanyingZHANGJiehuiYINPingpingSHEN
Qingshang SONG, Yue XUE, Yanying ZHANG, Jiehui YIN, Pingping SHEN
Ocean School, Yantai University, Yantai 264005, China
Abstract Phaeocystis is an important bloom-forming species and over 100 blooms have occurred since 1997 along the Chinese coasts, while only the species P.globosa was described.In this project, a total of 246 phytoplankton samples collected from the Bohai Sea, the Yellow Sea, and the East China Sea in April 2021 were analyzed to evaluate the species diversity and geographical distribution of the genus Phaeocystis based on metabarcoding 18S rDNA sequence.Four described Phaeocystis species were recognized, including P.globosa, P.pouchetii, P.jahnii, and P.cordata, and each has distinguished geographical distribution characteristics.P.globosa was the most widespread and abundant species, and P.pouchetii was found in the Bohai Sea and the northern Yellow Sea with substantially elevated abundance.P.cordata was also a widespread species, but its abundance was relatively low, while P.jahnii gathered only in the southeastern East China Sea.Water temperature, phosphate as well as ammonium contents were found to be associated with the abundance of P.globosa, P.pouchetii, and P.jahnii significantly.Moreover, two uncharacterized Phaeocystis species were detected in the Chinese seas,indicating the diversity of the genus remains underestimated worldwide.
Keyword: Haptophyta; Phaeocystis; species diversity; distribution; Chinese seas; amplicon sequence variant
1 INTRODUCTION
PhaeocystisLagerheim (Haptophyta) is one of the most extensively studied genera of marine phytoplankton worldwide and is recognized both as a nuisance and ecologically important microalga(e.g., Schoemann et al., 2005; Medlin and Zingone,2007, and references therein).It plays an essential role in the marine ecosystem, particularly the carbon and sulfur biogeochemical cycles by producing substantial amounts of dimethylsulphoniopropionate(DMSP) and volatile catabolite dimethylsulphide(DMS) which can impact the cloud albedo in the atmosphere, hence climate regulation (Stefels et al.,2007; Verity et al., 2007 and references therein).Members of the genus have a complex polymorphic life cycle alternating between solitary free-living cells and gelatinous colonies where hundreds of cells are embedded in a polysaccharide matrix(Rousseau et al., 2007, 2013; Zingone et al., 2011).Colonial species can form nearly monospecific extensive blooms (contributing>90% of total phytoplankton abundance), which are associated with dramatic ecosystem changes and negative impacts on fisheries and fish farming from the polar to tropical oceans (Hansen et al., 2003; Peng et al.,2005; Schoemann et al., 2005; Smith et al., 2014;Xu et al., 2019).In addition to the well-documented colony-forming species, the non-colonial species ofPhaeocystisspecies have been suggested as potentially important components of open ocean nanoplankton communities (Zingone et al., 1999;Lin et al., 2014; Gran-Stadniczeñko et al., 2017).
Up to now, tenPhaeocystisspecies have been described around the world since the establishment of the genusPhaeocystisin 1893 (Medlin and Zingone, 2007; Andersen et al., 2015; Shen and Qi,2021).Among them, three colonial species,P.globosaScherffel,P.pouchetii(Hariot) Lagerheim,andP.antarcticaKarsten are the most frequently recorded species and are known to form dense blooms in major climatic regions, i.e., the Arctic, the Antarctic, and temperate/tropical regions (e.g.,Schoemann et al., 2005).Compared with the welldocumented colony-forming species, less is known about the occurrence and significance of solitary species, includingP.scrobiculataMoestrup,P.cordataZingone et Chrétiennot-Dinet,P.jahniiZingone, which are supposed to be distributed worldwide (Moestrup, 1979; Zingone et al., 1999).The newly discovered unicellular speciesP.rexAndersen, Bailey, Decelle & Probert was observed from the Arabian Sea for the first time in 2015(Andersen et al., 2015).Other 3 species, includingP.amoeboideaBüttner (1910),P.sphaeroidesBüttner(1911), andP.bruceiMangin (1922) have never been found again after their first description (Medlin and Zingone, 2007).Besides the recognized species,several undescribedPhaeocystisspecies were reported from the South China Sea, and the western Mediterranean Sea based on morphological characteristics (Doan-Nhu and Larsen, 2010;Novarino, 2014).On the other hand, many uncultured clones belonging to this group were detected worldwide based on DNA metabarcoding(de Vargas et al., 2015; Wu et al., 2015; Shih et al.,2019).Most importantly, two unknownPhaeocystisspecies have been found in the Indian and Pacific Oceans to form endosymbiotic associations with the Acantharia (Decelle et al., 2012), and these symbiont sequences form the basal clades in the phylogeny ofPhaeocystis, indicating a wider diversity of the genus need to be characterized.
The firstP.globosabloom in China was recorded in 1997 on the southeastern coast of China (Chen et al., 2002).Since then,P.globosabloom has spread rapidly and occurred repeatedly along the South China Sea (Shen et al., 2018 and references therein).In 2004,P.globosabloom broke out in Bohai Bay for the first time (Qu et al., 2008) and bloomed frequently after that in the Bohai Sea and the northern Yellow Sea (Tu et al., 2011; Li et al.,2022).Until 2018, up to 97Phaeocystisbloom events have been recorded, covering an area of nearly 20 000 km2, but onlyP.globosawas reported and described (Shen et al., 2018; Shen and Qi, 2021;Wang et al., 2021), indicating that the species diversity and spatial distribution ofPhaeocystisare poorly understood in China.Moreover, studies have demonstrated thatP.globosastrains isolated from coastal China display remarkable genetic diversity,which might include cryptic species or different ecotypes (Song et al., 2021; Zhang et al., 2021; Li et al., 2022).Moreover, bloom of unicellular species has been reported in the East China Sea, where two new record speciesP.cordataandP.jahniiwere found at high density (Lin et al., 2014), indicating the solitary species may have been undersampled,primarily due to its small size and offshore location.Therefore, to clarify the species diversity and geographical distribution ofPhaeocystisin coastal China, high-throughput sequencing was used to investigate the species composition and abundance ofPhaeocystisin the Bohai Sea, the Yellow Sea, and the East China Sea in the present study.
2 MATERIAL AND METHOD
2.1 Sample collection
Surface water samples were collected from 82 sites in the Bohai Sea (BS), the Yellow Sea (YS),and the East China Sea (ECS) in April 2021 (Fig.1).The samples of the BS and the YS, and the ECS were collected onboard R/VLanHai101, andXiangyanghong18using SBE 32 Water Sampler with a SeaBird CTD system (SeaBird, USA),respectively.After collection, 3-L water samples were pre-filtered by a 200-μm mesh sieve to remove large particles and then collected using a vacuum pump onto a 2-μm pore size polycarbonate membrane filter (Millipore, USA).A total of 246 samples were obtained and stored under -80 ℃ before DNA extraction.
At each site, in-situ water temperature and salinity were measured by SeaBird CTD during the field sampling.Water samples for dissolved nutrients (nitrate, phosphate, ammonium, nitrite, and silicate) and chlorophyll-a(Chl-a) determination were filtered through 0.45-μm pore size cellulose filters, then both the water and filters were preserved at -20 °C in darkness before further laboratory processing.Nutrient concentrations were determined by SKALAR sa3000/5000 chemistry unit (SKALAR, the Netherlands) for the BS and the YS samples, and SEAL QuAAtro39 nutrient analyzer (SEAL Analytical, Germany) for the ECS samples, respectively.Chlorophyll-acontent was measured by TD10-AU fluorometer (Turner Designs,USA) according to the method of Parsons et al.(1984).

Fig.1 The locations of sampling sites in the Bohai Sea (B01–B19), the Yellow Sea (N02–N26, H01–H38), and the East China Sea (S1–S56)
2.2 DNA extraction and amplification
Planktonic community genomic DNA was extracted using the FastDNA®Spin Kit for Soil (MP Biomedicals, USA) according to the manufacturer’s instruction.The DNA extract was checked on 1%agarose gel, and its concentration and purity were determined with a NanoDrop 2000 UV-vis spectrophotometer (Thermo Scientific, USA).
The hypervariable region V4 of the 18S rDNA was amplified with primer pairs V4-F (5′-GCGG TAATTCCAGCTCCAATA-3′) and V4-R (5′-GATC CCCHWACTTTCGTTCTTGA-3′) (Song et al., 2016)by 9700 PCR thermocycler (ABI, USA).The PCR amplification was performed as follows: initial denaturation at 94 °C for 3 min, followed by 33 cycles of denaturing at 94 °C for 30 s, annealing at 58 °C for 45 s and extension at 72 °C for 45 s, and final extension at 72 °C for 5 min, and end at 4 °C.PCR reactions were performed in triplicate and the PCR products were extracted and purified using the DNA Gel Extraction Kit (Axygen Biosciences, USA),and then quantified using QuantusTMFluorometer(Promega, USA).
2.3 Sequencing and bioinformatics analysis
Purified amplicons were pooled in equimolar and paired-end sequenced on an Illumina NovaSeq PE250 platform (Illumina, USA) according to the standard protocols by Majorbio Bio-Pharm Technology Co.Ltd.(China).The sequences reported in this paper were deposited into the National Center for Biotechnology Information (NCBI) GenBank database (Accession Nos: ON129525–ON129547).The raw 18S rDNA sequence reads were deposited to NCBI under the project number PRJNA876366.
The raw 18S rDNA sequence reads were demultiplexed, quality-filtered by fastp version 0.20.0 (Chen et al., 2018), and merged by FLASH version 1.2.7 (Magoč and Salzberg, 2011).Only overlapping sequences longer than 10 bp were assembled and the maximum mismatch ratio of the overlap region is 0.2.Reads that could not be assembled were discarded.Samples were distinguished according to the barcode and primers,and the sequence direction was adjusted.
Then the high-quality sequences were de-noised using DADA2 (Callahan et al., 2016) plugin in the Qiime2 pipeline with recommended parameters(maxEE=2, truncQ=0).After that, the amplicon sequence variants (ASVs) were obtained and then annotated for species classification through the Protist Ribosomal Reference (PR2) database, setting the comparison threshold at 70%.For the ASVs that could not be annotated by the PR2database, a further taxonomic assignment was used by BLAST against the NCBI nucleotide database.Each ASV was annotated to the species level with the highest percentage identity (PID) at a cutoff level of 99%.The ASVs ofPhaeocystisshould meet the following conditions in sequence alignment with knownPhaeocystis: Query cover=100%,Evalue=0, PID>99%.ASVs information and sequence abundance ofPhaeocystisare shown in Supplementary Table S1.
The Maximum Likelihood (ML) phylogenetic tree was constructed using the MEGA X (Kumar et al., 2018) software package.The phylogenetic tree was inferred by the maximum likelihood (ML)method, using a Kimura 2-parameter model of DNA substitution.The bootstrap consensus tree was inferred from 1 000 replicates.The sampling sites and distribution of species were drawn with Surfer 16 (Golden Software LLC, USA) and GraphPad Prism 8 (GraphPad Software, USA).Spearman correlation coefficient was used to explore the relationship betweenPhaeocystisabundance and environmental factors.The spearman corresponding significance was tested and drawn using the corrplot package in R.Community network analysis among thePhaeocystisand other plankton species was carried out using the Spearman correlation coefficient in the R package psych.Principal component analysis (PCA) of the environmental factors was performed using software Canoco 5 (Microcomputer Power, USA) and the data was lg(n+1) transformed prior to performing PCA.
3 RESULT
3.1 Environmental condition
In April, the surface water temperature varied from 6.4 °C to 24.7 °C, with an average value of 13.2±4.2 °C.It was generally high in the ECS, with the highest value (24.7 °C) recorded at the southernmost station S55, and the lowest value(6.4 °C) occurred in the BS at the northernmost station B09.The salinity ranged from 29.1 to 34.5,with an average value of 32.4±1.3 across the whole studied area.In general, the salinity of the nearshore is lower than that of the open sea (Fig.2).
The concentration of nitrate, phosphate, and silicate showed a similar spatial distribution pattern,with the highest content occurring near the Changjiang(Yangze) River estuary and the ECS coast (Fig.3a,d, & e).The nitrate concentration ranged from 0.02 to 15.12 μmol/L, with a mean of 2.40±3.06 μmol/L.It was higher than 11.56 μmol/L around the Changjiang River estuary and higher than 6.63 μmol/L along the ECS coast.In addition,phosphate concentration ranged from 0.01 to 0.90 μmol/L (mean 0.21±0.17 μmol/L) and silicate concentration ranged from 0.52 to 21.20 μmol/L(mean 5.51±5.17 μmol/L).The nitrite concentrations were between 0.01 and 0.85 μmol/L (average 0.20±0.20 μmol/L), with the high values generally found near the Changjiang River estuary and the ECS coast (Fig.3b).The ammonium levels varied from 0.12 to 14.44 μmol/L, with an average of 1.64±2.15 μmol/L (Fig.3c).Two maximum of ammonium concentrations were found at stations H05 and H15 of the southern Yellow Sea.In addition, chlorophyll-acontent showed remarkable variability, with concentrations varying between 0.51 and 14.47 μg/L(mean 2.47±1.98 μg/L) (Fig.3f).
3.2 Species diversity and distribution of Phaeocystis

Fig.2 The distribution of surface water temperature (a) and salinity (b) along the Chinese seas
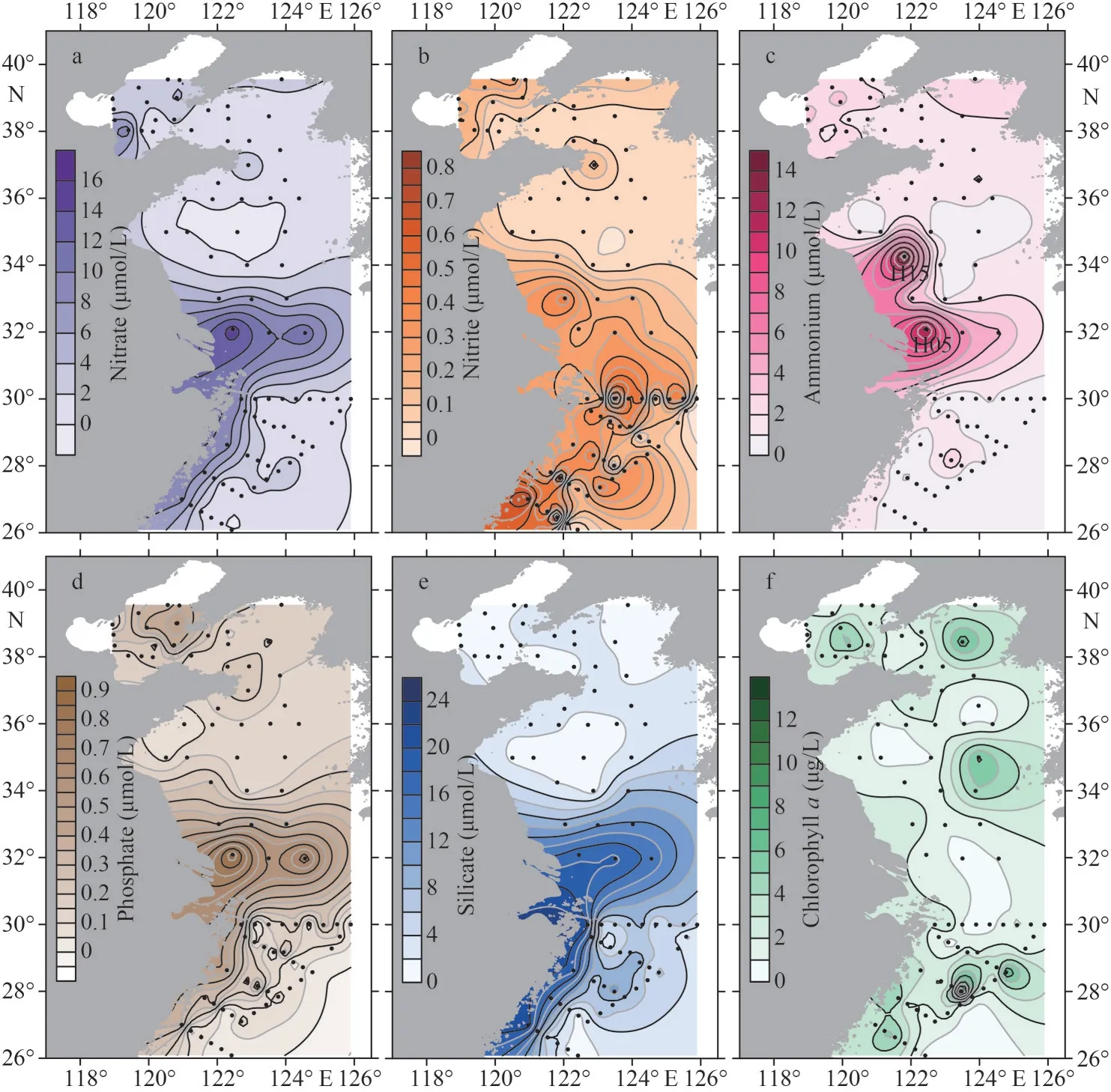
Fig.3 The distribution of nitrate (a), nitrite (b), ammonium (c), phosphate (d), silicate (e), chlorophyll a (f) along the Chinese seas
In this study, a total of 26 381 ASVs were identified from the environment samples in April 2021, and 18 ASVs were annotated to four recognizedPhaeocystisspecies (Figs.4–5), includingP.globosa,P.cordata,P.pouchetii, andP.jahniiusing the DADA2 analysis based on partial 18S rDNA sequences.Among them, the warm-water speciesP.globosadistributed widely throughout the Chinese seas, especially in the BS (Figs.4–5).The highestP.globosaabundance was found at Station H15 in the southern YS (Fig.4a).P.cordatais also a widespread species and occurred at most sampling stations, with the higher abundances at Stations H19 and H37 in the southern YS (Fig.4b).Cold-water speciesP.pouchetiidistributed only in the BS and the northern YS, with a substantially elevated abundance occurring at Station B09 in the BS,which was much higher than that of other stations(Fig.4c).The solitary speciesP.jahniiwas only recorded in the southeastern ECS with relatively low abundance (Figs.4d & 5).
Overall,Phaeocystiswas found at 58 out of 82 sampling stations.From a biodiversity perspective,the number ofPhaeocystisspecies was relatively low in the southern Yellow Sea, which was dominated byP.globosain terms of abundance(Fig.5).In addition, several ASVs from the southern Yellow Sea were annotated yet uncharacterizedPhaeocystisspecies (Fig.4e), which can be clustered into one distinguished clade (Fig.6).Clade 1 is a sister taxon to theP.globosa, but is significantly different from the knownP.globosaspecies,suggesting the existence of possible cryptic species ofP.globosa.
3.3 Correlation between environmental factors and Phaeocystis
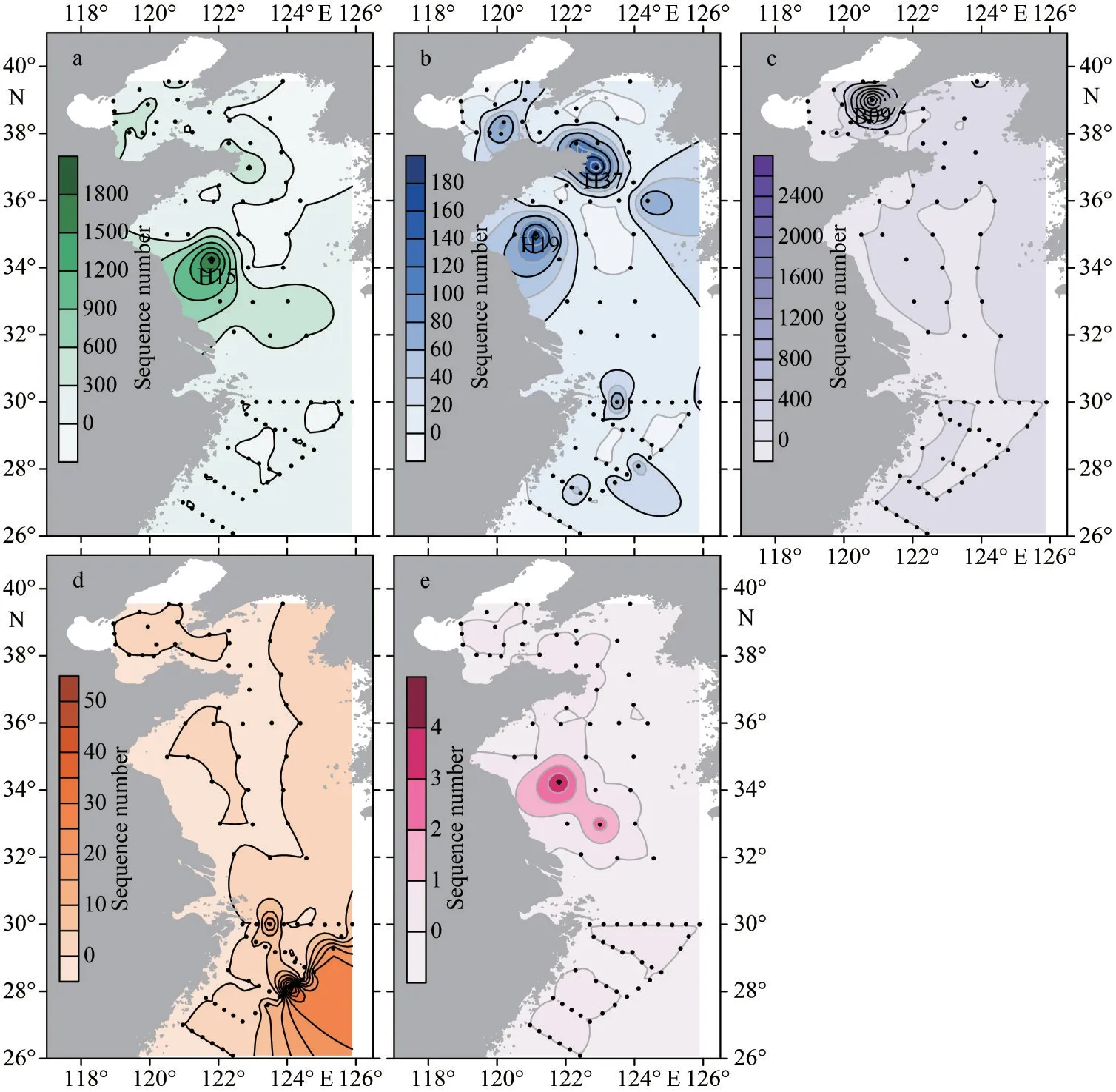
Fig.4 Distribution of different Phaeocystis species along the Chinese seas based on the number of DNA reads
Principal component analysis of environmental factors basically divides the sampling stations into three groups (BS, YS, and ECS), which are consistent with their geographical areas (Fig.7).The results indicate that 56.0% and 21.6% of the variation could be explained by PC1 and PC2,respectively.Samples of the ECS are closely related to high temperature, salinity, and high silicate, while samples from the BS showed opposite characteristics with low temperature, low salinity, but high ammonium and phosphate.No obvious features were observed for the YS’s samples, with half of them characterized by low nutrients (Fig.7).To further explore the impact of environmental factors onPhaeocystisabundance and distribution, the correlation between them was analyzed.TheP.globosaabundance was positive significantly related to phosphate and ammonium concentrations(P<0.05,n=82), but negatively related to the water temperature, salinity, and silicate concentration (P<0.05,n=82), respectively.The correlation between theP.pouchetiiand environmental factors was similar to those ofP.globosa, but the significant levels were much higher, in particular the effect of water temperature and salinity (P<0.001,n=82).On the contrary,P.jahniiwas positively related to the water temperature and salinity (P<0.05,n=82), and negatively related to phosphate and ammonium concentrations (P<0.01,n=82).
In addition, community network analysis among all the eukaryotes species indicated the complicated relationships among the plankton, including competition and predation.In general, differentPhaeocystisspecies showed negative response to ciliates,copepods, and some dinoflagellates and diatom species (e.g.,Noctilucasp.andChaetocerosdecipiens),but usually positive related to phytoplankton, in particular several clades of Syndiniales dinoflagellates(Syndiniales Groups I and II; Supplementary Table S2).
4 DISCUSSION
4.1 Phaeocystis species and their distribution

Fig.5 The relative abundance and distribution of different Phaeocystis species at each sampling site
In the present study, 4 describedPhaeocystisspecies, includingP.globosa,P.pouchetii,P.cordata,andP.jahnii, were detected in the Chinese seas in April 2021.Except forP.globosa, the other 3 species are new records to several seas, implying the species number of the genus is severely underestimated on Chinese coasts (Shen and Qi,2021).Although the South China Sea (SCS) was not investigated this time, previous studies have demonstrated thatP.globosadistributed widely in the SCS (Hai et al., 2010; Shen et al., 2018).Most interestingly, two cold-water speciesP.antarcticaandP.pouchetiiwere found in a cold eddy in the tropical SCS with high abundance (Wu et al., 2015).Moreover,P.scrobiculatawas also observed on the Vietnam coast along with the SCS based on morphological features (Doan-Nhu and Larsen,2010), revealing a much higher diversity of thePhaeocystisthan previously recognized in the SCS,which is now recognized as a hotspot ofPhaeocystisdistribution and biodiversity (Shen and Qi, 2021).
There is no doubt thatP.globosais the most widespread and abundant species in the world’s oceans, including off the China coasts (Schoemann et al., 2005, and references therein).From 1997 to 2022,P.globosahas caused over 100 bloom events in China, including 13 events in the Bohai Sea, 5 blooms in the East China Sea, and most remaining(over 80 events) in the northern South China Sea(e.g., Wang et al., 2021; Li et al., 2022).In late November 2021, a large-scaleP.globosabloom broke out in the southern Yellow Sea for the first time (Li et al., 2022), and just before that in April,the highest abundance ofP.globosawas detected near this region at Station H15 (Fig.4a), suggesting a great potential risk of the bloom outbreak.In addition, the ammonium concentration at this station and during theP.globosabloom in the southern Yellow Sea in late November were both high,implying an important effect of the ammonium on the growth ofP.globosa(this study; Wang et al.,2011; Li et al., 2022).Besides ammonium,phosphate also showed a positive significant influence on theP.globosaabundance in this study.
In fact, numerous studies have demonstrated that nutrients play an essential role in regulating the outbreak and maintenance ofP.globosablooms(Schoemann et al., 2005; Lancelot et al., 2007; Xu et al., 2019), but the results were depending and other effects such as temperature and induced allelopathy should be taken into account.Moreover,although noPhaeocystisblooms occurred in the Bohai Sea in recent years,P.globosais still widely distributed in the Bohai Sea, indicating the high potential risk ofP.globosabloom reoccurrence in this area.
Phaeocystis pouchetiiis another important bloomforming species that has caused dense blooms in the high latitude northern seas, including the New England continental shelf, Massachusetts Bay, and the Arctic Ocean (Saiz et al., 2013; Borkman et al.,2016; Smith et al., 2021).It can survive at water temperatures between -1.7 and 9 °C, but is better adapted to temperature below 5 °C (Schoemann et al., 2005; Simo-Matchim et al., 2017).In this study,extremely high abundance ofP.pouchetiiwas found in the northern Yellow Sea and the Bohai Sea, where the water temperature was the lowest (6.4–9.6 °C).Moreover, a strong negative correlation betweenP.pouchetiiand temperature was detected, indicating that low temperature is critical to the distribution and abundance ofP.pouchetiiin the Chinese seas.On the other hand, based on the high abundance ofP.pouchetii, it is reasonable to speculate that previously reportedPhaeocystisblooms from the Bohai Sea were possibly initiated byP.pouchetiiand/orP.globosasince only limited morphological and molecular evidence is available for most of them (Qu et al., 2008; Tu et al., 2011).Hence,identification of the causative species needs to be strengthened, especially in the Bohai Sea.

Fig.6 Phylogeny of Phaeocystis based on 18S rDNA V4 region sequences
Two new record species,P.jahniiandP.cordata,were further observed in coastal China (Lin et al.,2014).Based on the phylogenetic analysis,P.jahniiandP.cordataare located in the base branch of the phylogenetic tree and differentiated early in this genus, which should be cosmopolitan species in the world’s oceans (Zingone et al., 1999; Lange et al.,2002).The distribution ofP.jahniiandP.cordataon the Chinese coasts confirmed the prediction of widespread species once again (Zingone et al.,1999).Furthermore, it is worthy to note that a bloom with a high density ofP.jahniiandP.cordata(2.2×104cells/mL) was observed in the ECS in the spring of 2009 (Lin et al., 2014).In this study, an elevated abundance ofP.jahniiwas found in the southeastern East China Sea, indicating a high potential risk of solitary cell bloom in this area,where the temporal and spatial distribution ofP.jahniimay have been under-sampled.Moreover,P.jahniiwas positively related to the water temperature and salinity, while negatively related to phosphate and ammonium concentrations.These features may further indicate thatP.jahniiprefers warm open ocean areas; hence, the blooms of solitary cells have been reported on occasion (e.g.,Wassmann et al., 2005; Lin et al., 2014).
4.2 Uncharacterized Phaeocystis species
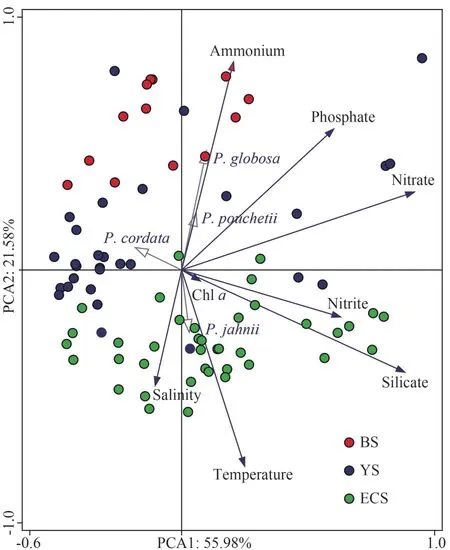
Fig.7 Principal component analysis of environmental factors and Phaeocystis abundance
Besides the above 4 described species, 2 uncharacterizedPhaeocystisspecies have also been detected in the southern Yellow Sea.One is a possible cryptic species ofP.globosa, and the other is a sister taxon to the speciesP.rex.In fact,uncharacterizedPhaeocystisspecies are widely distributed in the world’s oceans (Schoemann et al,2005; Medlin and Zingone, 2007; Shen and Qi,2021).For example, 3 undescribed species and 6 single-cell morphology from the Mediterranean Sea have been reported (Medlin and Zingone, 2007;Novarino, 2014).However, the common problem is that it is uncertain whether these cell types can represent biologically independentPhaeocystisspecies or the cell morphology of the same species at different life stages (Shen and Qi, 2021).Anyway,the existence of uncharacterizedPhaeocystisspecies indicates that the diversity of the genus remains poorly understudied worldwide.It is partly due to the tiny size of the single-cell species ofPhaeocystis, which is easy to be ignored in routine microscopic detection.At present, high-throughput sequencing of rRNA genes combined with metabonomics analysis can be used to investigate species diversity in a wide range of sea areas,especially single-celled algae with complex life cycles (Stoeck et al., 2010; De Vargas et al., 2015).Recently developed analysis methods based on amplicon sequence variants (ASVs) are more sensitive to distinguishing sequence variation.Therefore, the situation of the classification could be improved significantly in the future.
5 CONCLUSION
High-throughput sequencing data from 82 stations expanded our understanding of the species diversity and spatial distribution ofPhaeocystisin coastal China.The results showed that at least 4 describedPhaeocystisspecies, includingP.globosa,P.pouchetii,P.cordata,P.jahnii, and 2 uncharacterized species,coexisted on the Chinese coast.P.globosaandP.cordataare eurythermal and euryhaline species with a wide distribution.The cold-water speciesP.pouchetiihas also been found off the coast of China,particularly in the Bohai Sea, andP.jahniiprefers warm open ocean areas.Furthermore, environmental factors play an important role in affecting the abundance and geographic distribution ofPhaeocystis,especially the temperature.More new species ofPhaeocystisare yet to be discovered, and the metabarcoding analysis is an effective method for detecting species and profiling the biodiversity of thePhaeocystis.
6 DATA AVAILABILITY STATEMENT
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
7 ACKNOWLEDGMENT Samples of the BS and the YS were collected onboard R/VLanhai101implementing the open research cruise NORC2021-01 supported by NSFC Shiptime Sharing Project (No.42049901).Sample collections of the ECS were supported by NSFC Open Research Cruise (No.NORC2021-02+NORC2021-301).These cruises were conducted onboard R/VXiangyanghong18by the First Institute of Oceanography, Ministry of Natural Resources,China.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Trends of carbon and nutrient accumulation through time in the Andong salt marsh, Hangzhou Bay, China*
- Physical processes determining the distribution patterns of Nemopilema nomurai in the East China Sea*
- Comparison in structure and predicted function of epiphytic bacteria on Neopyropia yezoensis and Neopyropia katadae*
- Interaction between macroalgae and microplastics: Caulerpa lentillifera and Gracilaria tenuistipitata as microplastic bio-elimination vectors*
- Lake regime shift from submerged macrophyte to phytoplankton affected phosphorus speciation in sediment and eutrophic state in Caohai Lake, Guizhou, China*
- Temporal characteristics of algae-denitrifying bacteria co-occurrence patterns and denitrifier assembly in epiphytic biofilms on submerged macrophytes in Caohai Lake, SW China*