树莓茎对低温胁迫的生理响应
2023-12-19王鑫月张立业张雪梅
王鑫月,张立业,张雪梅
(河北农业大学林学院,河北保定 071001)
0 引言
树莓(Rubusidaeus)是一种多年生落叶小浆果类灌木,属于蔷薇科(Rosaceae)悬钩子属(Rubus)[1],因含有丰富的矿质元素、维生素且有较高的药用价值,被广泛应用于医药、食品等领域[2]。低温寒害是植物生长过程中面临的主要非生物胁迫因素之一,低温寒害限制植物生长,影响作物产量[3]。树莓栽培品种地上部分(茎)耐寒性差,需采取保护措施才能正常越冬,致使中国北方地区树莓越冬前必须埋土防寒。有些树莓野生资源可以自然越冬,而导致两者越冬性差异的原因尚未明确[4]。
在逆境条件下,脯氨酸既有渗透调节作用,还能保持膜结构的完整性,植物会积累大量的脯氨酸来抵抗逆境带来的损伤[5-8]。可溶性蛋白质有较强的亲水能力,可促进细胞保水,进而提高植物抗寒性[9]。在低温胁迫下,植物通过积累可溶性糖,进而增加细胞液浓度,延缓细胞结冰伤害[10-11]。植物体内活性氧含量会因温度降低而增加,当积累量超过正常水平时,细胞内产生和消除自由基的平衡便会遭到破坏。SOD、CAT和POD 3种酶紧密联系成为植物体内抗氧化防御系统的重要部分[5],同时具有清除植物体内活性氧的抗氧化能力[12-13]。低温胁迫会导致植物细胞膜系统受损,引起细胞膜收缩不均匀,膜脂相变[14-15]。植物细胞膜质过氧化和膜蛋白间会加速聚合,导致细胞内MDA含量增加[16-17]。因此,植物在低温胁迫下受到的伤害程度,可以通过测定电解质渗出率和MDA含量变化来反映。树莓在低温逆境下是否有同样的生理响应需要进一步研究。
为探究抗低温野生树莓资源牛叠肚与低温敏感型树莓栽培品种抗寒性产生差异的原因,本研究从生理特性角度开展系统研究,以期为解析树莓抗寒性突破树莓栽培品种越冬障碍提供研究基础,为农业生产上选育树莓抗寒品种提供理论指导。
1 材料与方法
1.1 试验材料
试验材料均为树莓初生茎,野生资源牛叠肚包括牛叠肚1号和牛叠肚2号,栽培品种有‘波拉娜’、‘菲尔杜德’和‘海尔特兹’。牛叠肚1 号引自张家口市黑龙山林场(116°10′71′′E、41°28′58′′N),牛叠肚2 号引自唐山市迁西县(118°26′34′′E、40°15′38′′N),栽培品种均产自保定市市区(115°45′23′′E、38°83′34′′N)。试材均于2016年移栽至试验田,株行距为1 m×2.5 m,冬季不做防寒处理,使其自然越冬。
1.2 试验设计
试验地位于保定市河北农业大学实验基地(115°25′3.98′′E、38°49′32.33′′N),海拔14.3 m,温带大陆性季风气候。1、2月气温最低,1月日均最高气温为2℃,日均最低气温为-9℃,极低温度可达-22℃左右。越冬期平均气温以及平均空气湿度如表1所示。在表2中的10 个时期,分别选取长势一致、生长健壮的不同品种树莓3株,每株取中部茎用于测定生理指标。

表1 试验地越冬期平均气温及平均湿度
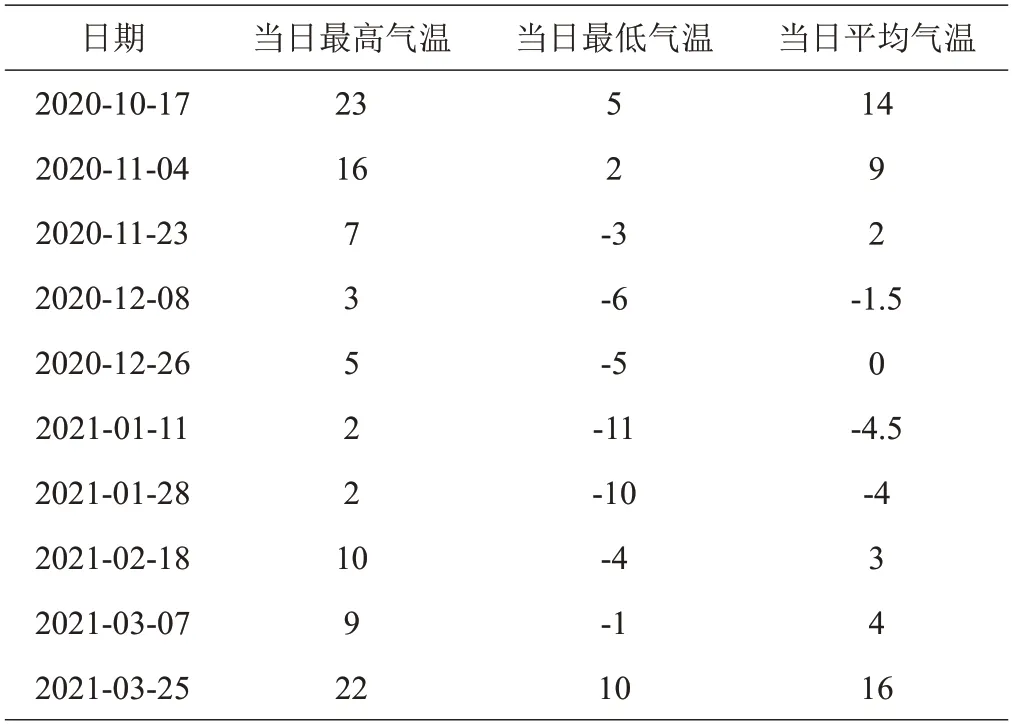
表2 采样时间及温度 ℃
1.3 测定指标及方法
相对含水量采用烘干称重法[18]。
渗透调节物质脯氨酸含量采用磺基水杨酸法,可溶性糖和淀粉含量采用蒽酮比色法,可溶性蛋白质含量采用考马斯亮蓝G-250染色法测定[19]。
超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性测定,分别采用氮蓝四唑(NBT)光还原法、愈创木酚法、过氧化氢法[19]。
相对电导率用电导仪测定,丙二醛含量采用硫代巴比妥酸法测定[19]。
1.4 数据处理及分析
数据分析及作图应用Microsoft Excel,采用Duncan 新复极差法对数据进行方差分析,并应用SPSS 26.0 软件进行各指标的相关性分析及主成分分析,运用主成分分析法对10个生理指标进行树莓茎抗寒能力综合评价。
2 结果与分析
2.1 低温对树莓茎相对含水量的影响
在自然越冬条件下树莓茎的相对含水量变化整体呈先下降后上升趋势(图1)。各品种在1月11日出现低峰值,较10 月17 日牛叠肚1 号、2 号和栽培品种‘波拉娜’、‘菲尔杜德’和‘海尔特兹’分别减少了42.47%、41.11%、79.82%、82.41%和85.53%,其中‘海尔特兹’减少最多,牛叠肚2 号减量最少;1 月下旬,随着气温回升,茎相对含水量呈上升趋势。显著性分析表明,从12月26日开始,树莓野生资源牛叠肚茎相对含水量均显著高于栽培品种。越冬后(3月25日),牛叠肚1号、2号的相对含水量分别为76.34%、72.74%,比‘波拉娜’、‘菲尔杜德’和‘海尔特兹’分别高出了205%、221%、263%和190%、206%、246%。
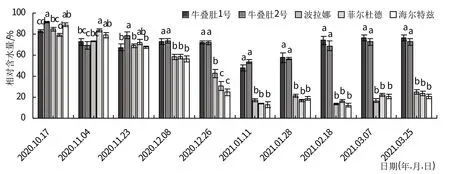
图1 越冬过程中树莓茎相对含水量变化
2.2 低温对树莓茎渗透调节物质含量的影响
随着冬季温度不断降低,树莓茎游离脯氨酸含量先增加后减少(图2)。越冬初期除‘海尔特兹’外,其他品种茎中游离脯氨酸含量均无显著差异,从11月初开始野生资源游离脯氨酸含量显著升高。其中牛叠肚1号茎游离脯氨酸含量在1月11日达到峰值(742.31 μg/g);牛叠肚2 号、‘波拉娜’茎游离脯氨酸含量在1 月末达到峰值,分别为755.21、296.55 μg/g;‘菲尔杜德’、‘海尔特兹’茎游离脯氨酸含量在2 月18 日达到峰值,分别为349.08、159.24 μg/g。
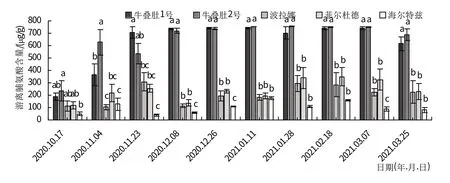
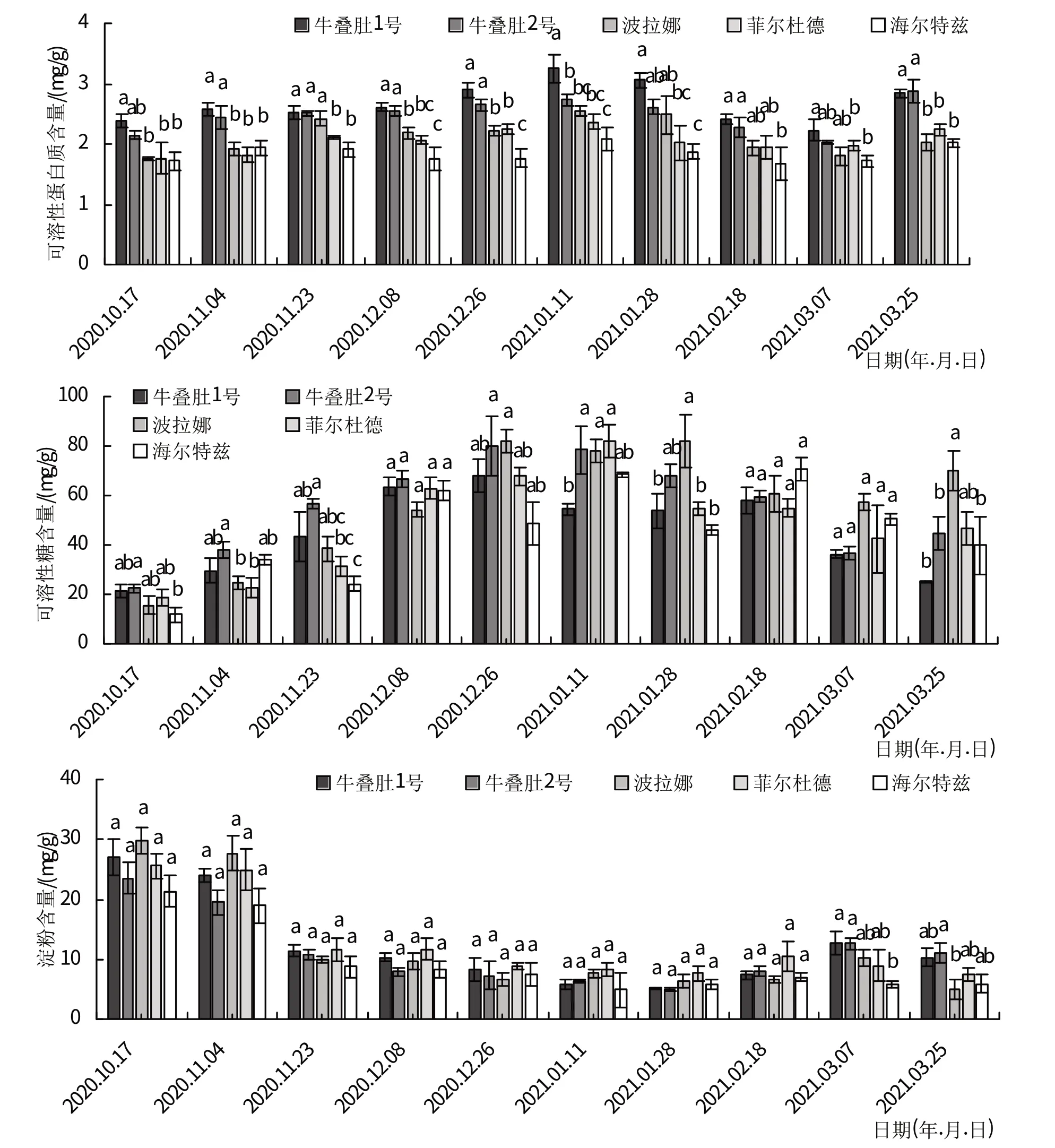
图2 越冬过程中树莓茎渗透调节物质含量变化
可溶性蛋白质含量变化整体呈先上升后下降趋势。随着温度降低,树莓茎可溶性蛋白质含量均在10月中旬—翌年1 月中旬期间逐渐积累,并且在1 月11日达到最高值,牛叠肚1号、牛叠肚2号、‘波拉娜’、‘菲尔杜德’和‘海尔特兹’较10 月17 日分别增加了36.63%、27.35%、45.37%、34.16%和21.42%。此后,随着温度的回升,可溶性蛋白含量明显下降。
牛叠肚1号、牛叠肚2号和‘波拉娜’茎可溶性糖含量均在10 月17 日—12 月26 日期间逐渐积累,并且在12 月26 日达到最高值,峰值分别为68.27、80.14 和82.06 mg/g。而‘菲尔杜德’和‘海尔特兹’茎可溶性糖含量在翌年1月11日达到最高值,较越冬初期分别增加了236%和383%。之后,随着温度的回升,可溶性糖含量呈波动性变化,整体表现出降低趋势。
淀粉含量和可溶性糖含量变化相反。树莓茎淀粉含量在1月28日前均无显著性差异,整体呈先下降后缓慢上升。除‘海尔特兹’品种外,其他品种茎淀粉含量均在10月17日—翌年1月28日期间逐渐下降,并且在1月末含量达到最低值,较上一年10 月中旬分别降低了80.91%、78.94%、78.68%和69.88%。而‘海尔特兹’茎淀粉含量在1月中旬达到最低值,最低值为4.86 mg/g。之后,随着温度的回升,淀粉含量缓慢上升。
2.3 低温对树莓茎抗氧化酶活性的影响
树莓茎SOD 活性的变化呈先下降后上升再下降的趋势(图3)。牛叠肚SOD活性在越冬多数时期均显著高于栽培品种。其中牛叠肚1号和2号在12月8日之前迅速下降,在12月8日达到第1个低峰值,分别为3.70 和3.50 U/g,随后SOD 活性开始上升,分别在2 月18 日和1 月11 日达到峰值。栽培品种树莓茎SOD 活性在11月23日之前迅速下降,‘波拉娜’、‘菲尔杜德’、‘海尔特兹’茎SOD活性分别在1月11日、1月28日和11月23日到达第1个低峰,分别为1.14、1.45、0.91 U/g,随后SOD活性有缓慢上升。
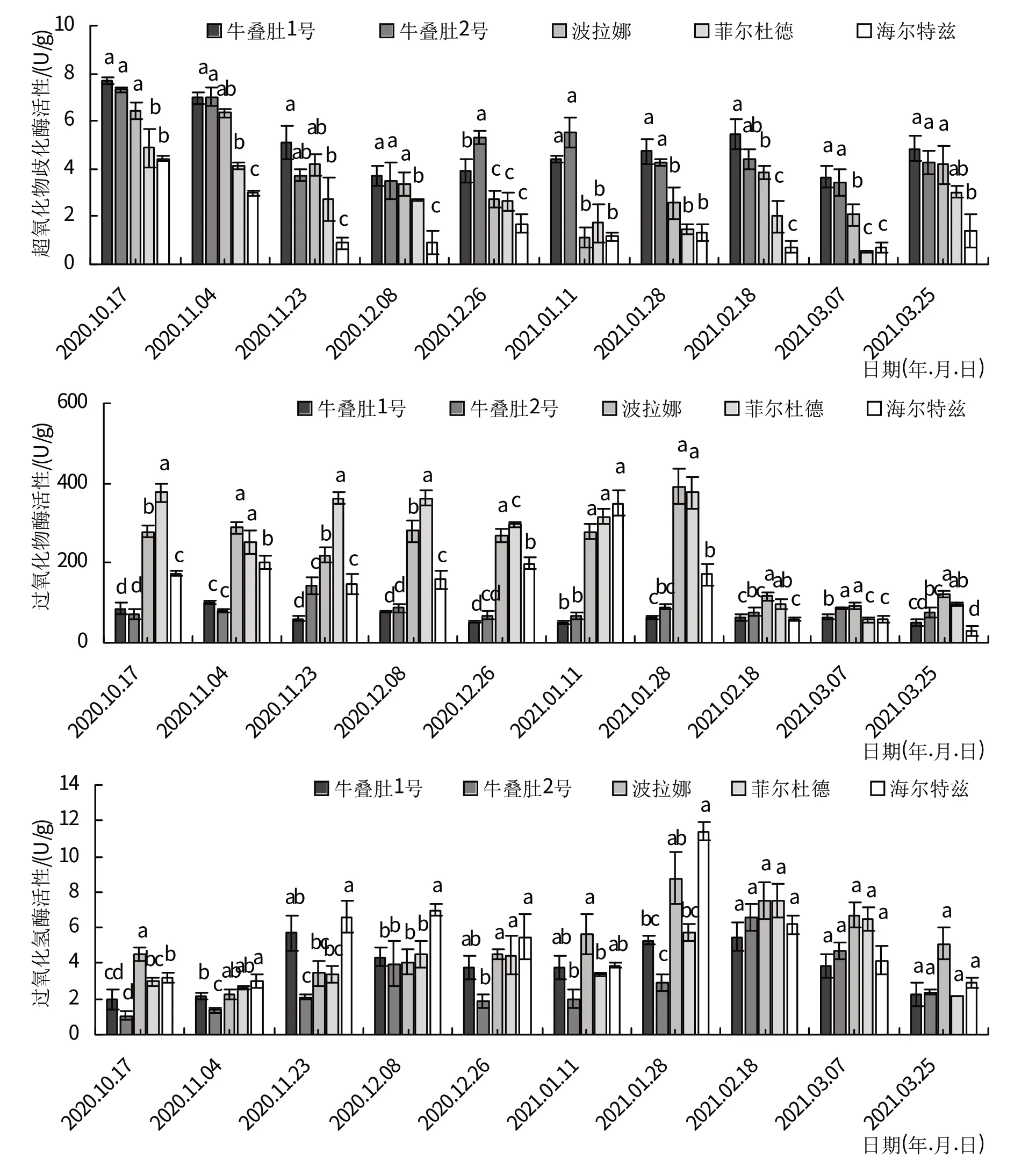
图3 越冬过程中树莓茎抗氧化酶活性变化
树莓栽培品种茎POD活性在越冬期间变化明显,而树莓野生资源变化较小。在2月18日之前,牛叠肚茎POD活性显著低于栽培品种。牛叠肚1号和2号茎POD活性分别在11月4日、11月23日达到第1个高峰值,分别为101.52、142.56 U/g,在1月11日达到了最低值50.75、65.84 U/g。‘菲尔杜德’茎POD活性在11月4日达到第1个低峰值251.23 U/g,‘波拉娜’和‘海尔特兹’在11月23日达到第1个低峰值220.13和147.06 U/g。‘海尔特兹’茎POD 活性在1 月11 日达到了最高值(348.70 U/g),‘波拉娜’(391.51 U/g)和‘菲尔杜德’(377.54 U/g)在1月28日达到了最高值。
树莓茎CAT 活性变化大致呈升—降—升—降的趋势。牛叠肚1 号茎CAT 活性在11 月23 日达到第1个高峰值,牛叠肚2 号、‘菲尔杜德’和‘海尔特兹’茎CAT 活性在12 月8 日达到第1 个高峰值。‘波拉娜’在11月23日达到第1个高峰值。‘波拉娜’和‘海尔特兹’茎CAT 活性在1 月28 日达到第2 个高峰值,而牛叠肚1号、2号和‘菲尔杜德’茎CAT活性在2月18日达到第2 个高峰值,分别为8.76、11.38、5.49、6.59 和7.51 U/g。越冬过程中,‘海尔特兹’茎的CAT 活性增加幅度最大,牛叠肚1号茎的CAT活性增加幅度最小。
2.4 低温对树莓茎细胞膜系统的影响
树莓茎相对电导率变化整体呈先上升后下降趋势(图4)。12 月8 日—1 月28 日,随着温度的降低,树莓茎的相对电导率逐步增加,牛叠肚1 号、牛叠肚2 号、‘波拉娜’和‘海尔特兹’茎的相对电导率在1月28日出现峰值,其中增加最多的是‘海尔特兹’,牛叠肚1号的增量最少;1 月28 日后,随着气温回升,茎相对电导率呈下降趋势。
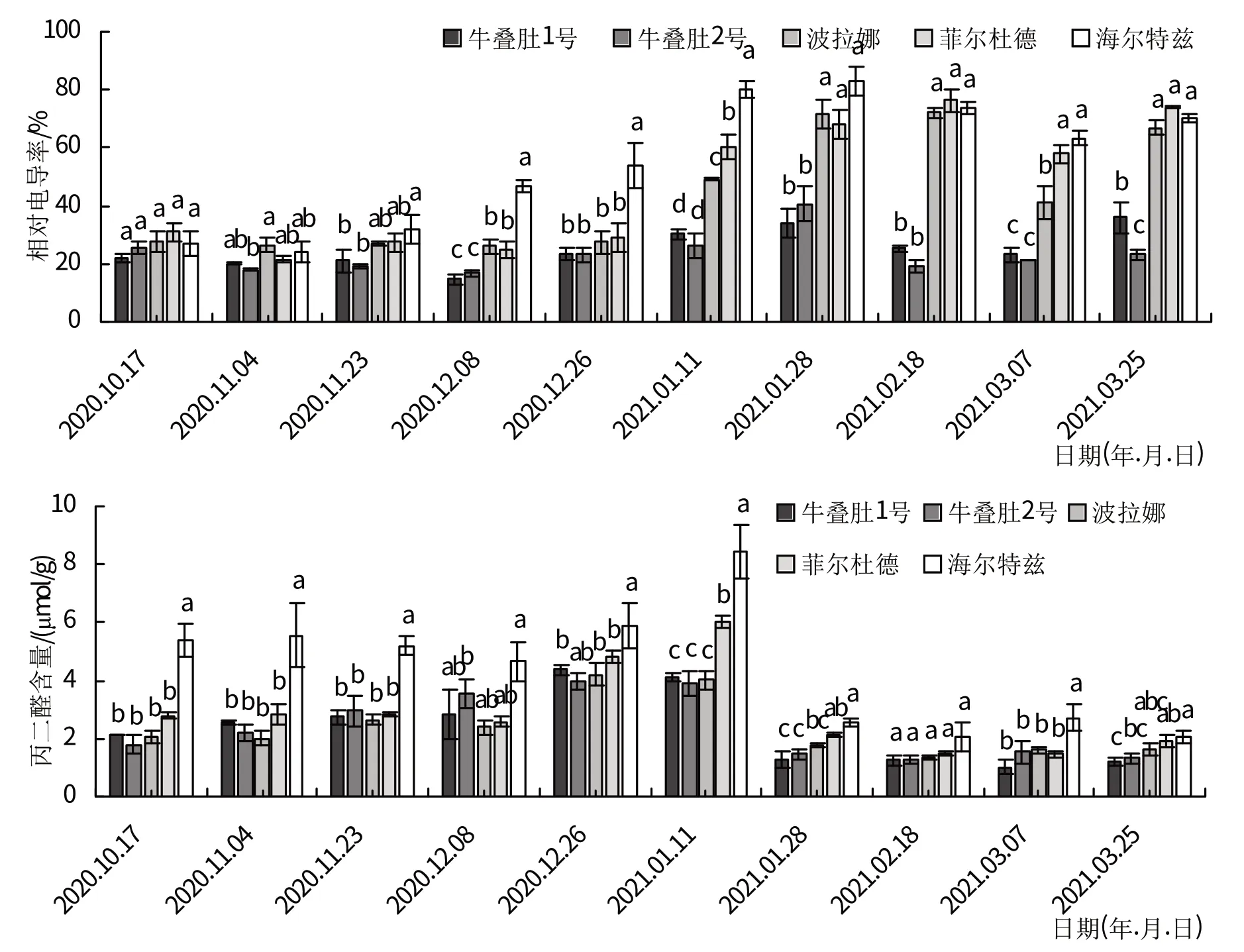
图4 越冬过程中树莓茎细胞膜系统变化
牛叠肚1号、牛叠肚2号和‘波拉娜’茎MDA含量在12月26日之前逐渐增加,并在12月26日达到最高值,峰值分别为4.37、3.97 和4.20 μmol/g;三者茎的MDA 含量在1 月开始下降,1 月末后变化幅度较小。‘菲尔杜德’和‘海尔特兹’茎MDA含量在1月11日之前逐渐增加,并达到最高峰(6.02、8.44 μmol/g),随后急速下降,在1月末后处于稳定阶段。
2.5 抗寒性指标相关性分析
由相关性分析(表3)可见,自然低温胁迫下2个树莓野生资源和3个树莓栽培品种的10个生理指标间均存在着一定的相关性。POD 活性与相对电导率显著正相关(r=0.932);与相对含水量、脯氨酸含量极显著负相关,相关系数分别为-0.984 和-0.983;与可溶性蛋白含量、SOD 活性显著负相关,相关系数分别为-0.895和-0.949。相对含水量与SOD 活性极显著正相关(r=0.984);与相对电导率显著负相关(r=-0.897)。脯氨酸含量与相对含水量、SOD 活性极显著正相关,相关系数分别为0.922 和0.978。相对电导率与MDA 含量显著正相关(r=0.917)。
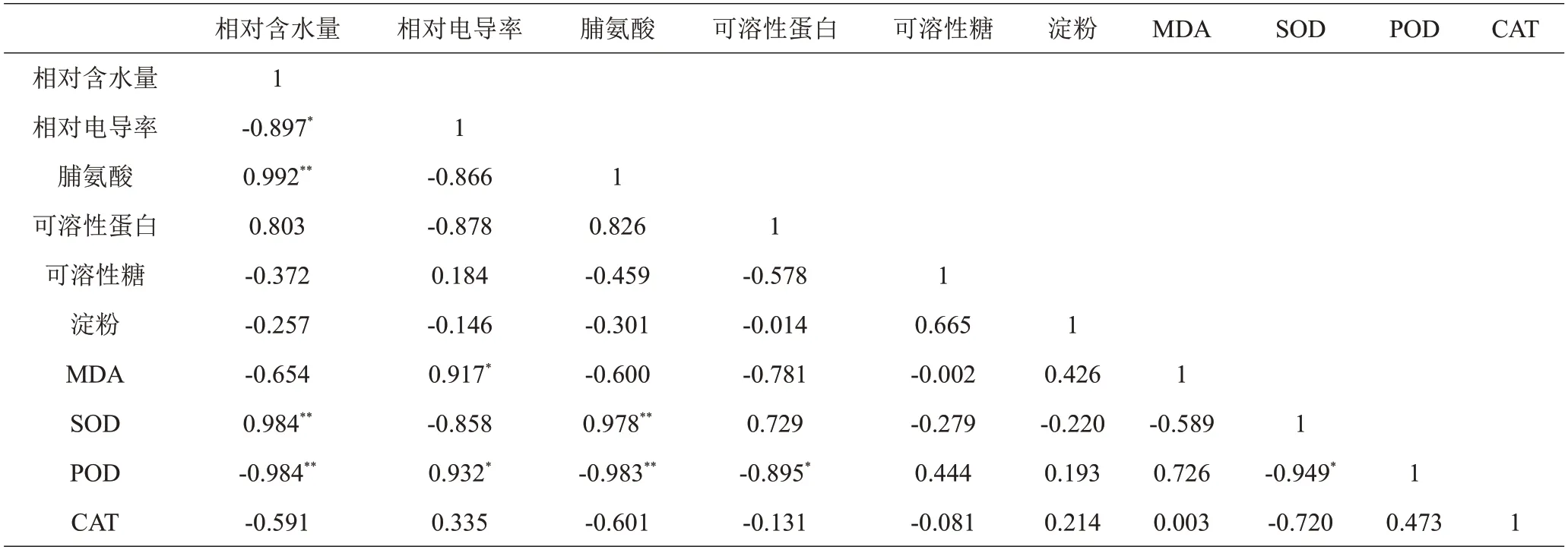
表3 树莓茎抗寒生理指标相关系数
2.6 生理指标主成分分析
以测定的10个指标为依据,应用主成分分析法综合评价低温对不同品种树莓茎相对含水量、渗透调节物质、抗氧化酶活性及膜透性的影响,结果见表4。前3 个主成分的贡献率分别为65.064%、19.417%、13.310%,累积贡献率达到了97.791%。表明前3 个主成分反映了绝大部分信息,可以代替原来10个指标对自然越冬过程中2个树莓野生资源和3个树莓栽培品种抗寒性的评价。第1 主成分特征值为6.506,其中POD活性的指标系数最大,为-0.999;其次是相对含水量、相对电导率、SOD活性和脯氨酸含量,指标系数分别为0.985、0.985、0.958 和-0.937。说明第1 主成分与这几个指标关系最为密切,代表了更多的抗寒评价信息。结合相关性分析可知,第1 主成分中POD 活性、相对含水量、相对电导率和脯氨酸含量均与其他多个指标有较大的相关性,代表了2个树莓野生资源和3个树莓栽培品种抗寒性的大部分信息。
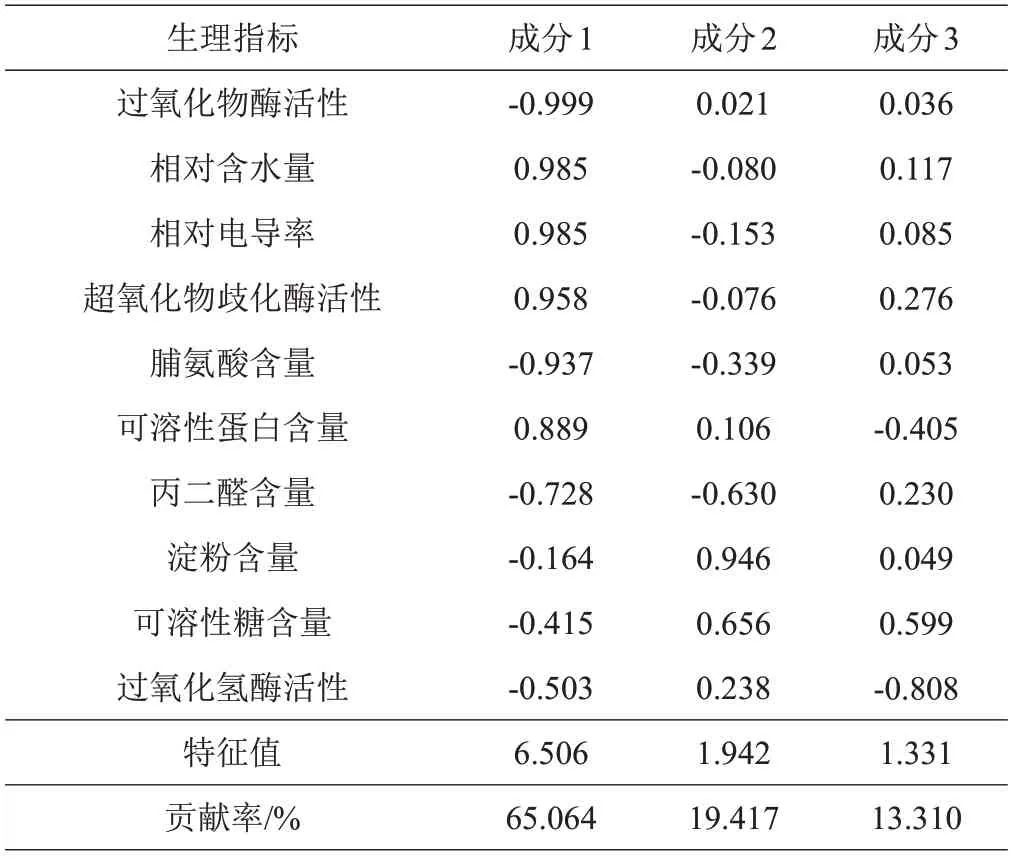
表4 树莓茎抗寒性指标因子负荷
通过表4 所示主成分载荷矩阵,得到以每个载荷量来表示主成分与对应变量的相关关系,通过计算得到主成分表达式[式(1)~(3)]。主成分综合评价模型如式(4)。
式中,F1代表第1主成分,F2代表第2主成分,F3代表第3主成分。Zx1代表过氧化物酶活性标准化后的数值,Zx2代表相对含水量标准化后的数值,Zx3代表相对电导率标准化后的数值,Zx4代表超氧化物歧化酶活性标准化后的数值,Zx5代表脯氨酸含量标准化后的数值,Zx6代表可溶性蛋白含量标准化后的数值,Zx7代表丙二醛含量标准化后的数值,Zx8代表淀粉含量标准化后的数值,Zx9代表可溶性糖含量标准化后的数值,Zx10代表过氧化氢酶活性标准化后的数值。
根据主成分综合评价模型即可计算2个树莓野生资源和3 个树莓栽培品种树莓茎的综合主成分值,并对其按综合主成分值进行综合抗寒能力评价排序(表5),牛叠肚1 号、2号抗寒性较强,‘波拉娜’抗寒较弱,‘菲尔杜德’和‘海尔特兹’抗寒性最差。
3 结论
本试验通过对树莓2 个野生资源和3 个栽培品种自然越冬过程中生理指标的测定,结合主成分分析对其抗寒性进行综合评价,明确了不同品种的抗寒性强弱,其抗寒性由强到弱为牛叠肚2号、牛叠肚1号、‘波拉娜’、‘菲尔杜德’和‘海尔特兹’。不同树莓品种对低温胁迫的响应机制不同,抗寒性产生差异的主要原因是牛叠肚1号和2号在越冬期茎保水能力更高,在低温胁迫下诱导产生渗透调节物质的水平较强,同时抗膜脂过氧化能力也发挥了重要作用。该研究为解析树莓抗寒性提供研究基础,为农业生产上选育树莓抗寒品种提供指导。
4 讨论
水分是植物生长过程中控制植物生命活动的重要组成成分,植物的含水量与其代谢活动有着密切的关系。本试验研究发现,野生资源牛叠肚和树莓栽培品种茎的相对含水量在低温期(2021 年1 月11 日)前呈下降趋势,说明随着温度降低,树莓茎不断失水而抗寒性下降,经过低温期后野生资源牛叠肚茎相对含水量又恢复到越冬期之前水平,但是栽培品种茎相对含水量越冬期后一直处于较低水平,低温敏感型树莓在越冬过程中相对含水量的变化趋势与姜蓓蓓[20]的研究结果一致。
植物能利用自身防御系统,通过调节代谢物质来降低逆境伤害[21]。脯氨酸含量增加可以帮助细胞保水,逆境胁迫下积累脯氨酸含量,有利于细胞恢复生长[22]。本试验结果表明,在越冬期树莓茎游离脯氨酸含量呈先上升后下降趋势,这是因为树莓茎在受到低温伤害时会促进脯氨酸的合成,使脯氨酸含量累积并增加。而在低温来临前野生资源牛叠肚茎脯氨酸含量升高幅度远远超过栽培品种,可以更好地应对低温伤害[23]。在自然越冬条件下树莓各品种茎可溶性蛋白质含量变化整体呈先上升后下降趋势。1月11日前由于植株受到了持续低温伤害,树莓茎可溶性蛋白质含量增加,从而增加细胞中束缚水含量,以减少原生质体结冰,降低伤害[24]。而1 月11 日后随着温度的上升可溶性蛋白质含量下降。但在3月25日树莓茎可溶性蛋白质含量又上升可能是由于春季温度上升,树莓茎水分丧失严重,植株受伤程度再次加重。这与自然越冬时姜花属植物生理指标变化及抗寒性评价结果一致[25]。同时在研究山西梨的抗寒性中发现,枝条可溶性糖含量越高其抗寒性越强[26]。可溶性糖积累使细胞中原生质浓度增加,以防细胞液结冰,增强流动性,是植物体内重要的保护物质[27]。本研究表明,越冬期树莓茎可溶性糖含量先上升后下降,1 月11 日前可溶糖含量逐渐增加,是由于随着温度降低,细胞通过积累可溶糖含量来减少低温对细胞的伤害;1 月11 日—3 月25 日环境温度逐渐上升,树莓茎可溶糖含量降低,而此时‘波拉娜’茎可溶性糖含量较高可能是由于其低温期后恢复能力较差。树莓茎淀粉含量与可溶性糖含量变化趋势相反,是由于淀粉转化为可溶性糖来抵抗低温[28]。
在低温胁迫下,植物代谢失调会产生大量自由基,而SOD 可清除O2-来发挥保护酶的作用[29]。本试验结果表明,野生资源SOD 活性高于栽培品种,是由于野生树莓利用高SOD 活性清除过剩的自由基来缓解低温环境带来的伤害。POD 在保护酶系统中主要的作用是降解过氧化物[30]。本研究发现,树莓野生资源茎POD 活性低于栽培品种,可能原因是,树莓栽培品种对低温更敏感,在越冬初期便启动POD 降解过氧化物,之后随低温胁迫强度加大其活性逐渐增强,促进细胞产生自我保护机制。而野生资源牛叠肚茎POD 活性在整个越冬期变幅不大,可能是牛叠肚含有较高的相对含水量及渗透调节物质使得植株抗寒能力更强,在低温胁迫下体内过氧化物增幅较小,不足以激发POD的响应。本研究结果与张玉霞等[31]对苜蓿抗寒性的研究结果一致。低温胁迫促使植物产生H2O2,而CAT能将其分解成水以减缓植物受伤程度[32]。在越冬期树莓茎CAT活性呈先上升后下降趋势,说明随着低温加剧,细胞内氧化物增加,为保护细胞不受伤害,细胞内CAT 酶活性增强,而后期气温回升,CAT 活性随即降低。野生资源牛叠肚茎CAT活性变化幅度更小,同样表明牛叠肚自身生理抗寒性更强,本结果与对葡萄砧木抗寒性结果一致[33]。
植物遭受低温胁迫,最先受到伤害的是细胞膜[34],破坏膜半透性,提高细胞膜通透性,致使膜内溶质外渗,最终引起植株死亡[35]。电导率能表征植物受低温伤害的程度[36]。本研究表明,随着自然低温强度加大,树莓栽培品种茎相对电导率急速增加,而野生资源牛叠肚茎相对电导率上升幅度不大。说明栽培品种细胞膜受损比野生资源牛叠肚更加严重,细胞电解质外渗程度更大,抗寒性更差。低温期后相对电导率居高不下可能是因为MDA 含量的积累损伤了生物膜[37]。MDA含量可以反映膜质过氧化的程度,膜质过氧化程度越低植物抗逆性就越强[38]。本试验表明,树莓不同品种茎MDA 含量在12 月8 日之前呈上升趋势,可能是越冬初期温度逐渐降低增加了树莓茎细胞膜脂过氧化程度;12月26日之后MDA含量有短暂的下降,可能是由于在低温下细胞内保护酶的应激反应,产生SOD、POD、CAT等物质以清除MDA对植物的伤害。
在中国北方地区,树莓栽培品种自然越冬十分困难。本研究主要是针对不同品种树莓茎越冬过程中相对含水量、渗透调节物质、抗氧化酶系统以及细胞膜系统的变化进行研究,解析野生树莓资源冬季抗寒的生理优势,为今后抗寒基因的筛选提供参考。然而,其他因素对树莓茎抵御低温是否有影响、如何影响需进一步研究,同时相同低温条件下,不同品种树莓茎细胞结构及潜在基因的研究应作为今后研究的重点,以早日消除栽培品种自然越冬障碍。
