瘤胃纤维素降解细菌的研究进展
2023-12-06李永丽燕亚平胡建华刘占英
高 星, 李永丽, 燕亚平, 胡建华, 刘占英*
(1.内蒙古工业大学 化工学院,内蒙古 呼和浩特 010051;2.内蒙古自治区发酵产业节能减排工程技术研究中心,内蒙古 呼和浩特 010051;3.生物发酵绿色制造内蒙古自治区工程研究中心,内蒙古 呼和浩特 010051)
纤维素(cellulose)是地球上分布最广泛、储存量最大的有机可再生资源,具有广阔的开发前景[1]。由于纤维素的特殊木质化和结晶化使得纤维素难以被降解,如何高效利用纤维素是当前研究的一大热点[1]。目前,能将纤维素高效降解成可被利用的还原糖的方法主要有物理降解、化学降解和微生物降解等[2]。化学和物理降解不仅消耗大量的能源,同时在分解的过程中产生大量的污水、尾气等,而微生物降解主要存在效率较低的问题。反刍动物瘤胃作为一种独特的消化系统,其中细菌在反刍动物瘤胃中种类较多、数量最大,因此可以有效的降解纤维素,是瘤胃纤维降解的主体。本文从瘤胃微生态的视角探究瘤胃纤维降解细菌的功能,进而深入揭示纤维降解细菌对纤维素的降解机制,对于全面发掘瘤胃纤维降解细菌和纤维素酶资源有较大的理论价值与现实意义。
1 瘤胃纤维降解细菌的主要种类
1.1 产琥珀酸丝状杆菌
产琥珀酸丝状杆菌(Fibrobactersuccinogenes)为革兰阴性菌,作为主要的纤维降解细菌在瘤胃中普遍存在。大量研究表明产琥珀酸丝状杆菌在木质纤维素的降解过程中发挥着重要的作用,但不能够降解木聚糖。Maglione等[3]对产琥珀酸丝状杆菌的相关问题进行研究,结果显示,产琥珀酸丝状杆菌若处于非生长期不分泌产生纤维素酶,从而无法分解纤维素。在此基础上,研究了碳源对产琥珀酸丝状杆菌降解纤维能力的影响,通过对比实验对纤维降解速率进行分析,分析结果表明,以纤维二糖作为培养底物时,产琥珀酸丝状杆菌可以快速的分解纤维素;而硫代纤维二糖使得产琥珀酸丝状杆菌无法降解纤维素,推断产琥珀酸丝状杆菌产生的纤维素酶对反馈抑制较为敏感。同样,Field等[4]在底物对产琥珀酸丝状杆菌降解能力的影响研究中,通过设置以纤维二糖和纤维素作为碳源进行对比分析,表明菌体生长速度和代谢产酸的速度基本相同,但是菌体对纤维素底物的降解速率不同,最终确定了产琥珀酸丝状杆菌降解对纤维素的存在形式具有选择性。产琥珀酸丝状杆菌在纯培养下具有很强的纤维降解能力,王梦芝等[5]在蛋白质来源对瘤胃细菌影响的相关问题进行研究中,发现产琥珀酸丝状杆菌分泌的纤维素酶有一个特殊的纤维催化区和纤维素结合区,从而使得该菌可以迅速的降解一些其余菌种所不能分解的纤维素。
1.2 白色瘤胃球菌
白色瘤胃球菌(Ruminococcusalbus)属于严格厌氧的革兰阳性菌,在瘤胃中作为主要的纤维降解菌广泛存在。在培养白色瘤胃球菌时,需要一些维生素等生长因子。周非帆[6]从绵羊瘤胃中筛选出4株菌,对纤维降解特性的研究发现,其中白色瘤胃球菌具有最高的糖苷水解酶酶活力。此外,Hoda等[7]分析了白色瘤胃球菌分泌的纤维素酶结构、功能和系统发育,结果表明,白色瘤胃球菌产生的纤维素酶具有高度的稳定性,蛋白质主要由随机螺旋和α螺旋组成,说明白色瘤胃球菌分泌的纤维素酶具有较高的耐热性,并且90.1%的氨基酸处于有利区域,从酶结构上解释了白色瘤胃球菌降解纤维素的高效能力。Yeoman等[8]在对产琥珀酸丝状杆菌、白色瘤胃球菌、黄色瘤胃球菌(Ruminococcusflavefaciens)之间的竞争作用研究中发现3株菌在纤维素上黏附生长并且进行竞争,当接种黄色瘤胃球菌后,产琥珀酸丝状杆菌逐渐减少,但纤维素的降解并没有减少,说明产琥珀酸丝状杆菌与黄色瘤胃球菌采用不同但同样有效的方法分解植物细胞壁。
1.3 黄色瘤胃球菌
黄色瘤胃球菌作为三种主要纤维降解菌之一,在瘤胃中大量存在。研究表明,黄色瘤胃球菌所需要的营养物质主要为纤维素,具有高效降解纤维素的能力。王卫云等[9]从内蒙古绵羊瘤胃中筛选出两株黄色瘤胃球菌H1、H2,H2的滤纸酶活力达到了0.21 μmol/(mL·min),具有良好的纤维素降解能力。此外,刘玉承等[10]从绵羊瘤胃中筛选出32株菌,经鉴定两株黄色瘤胃球菌CCQ、LYQ滤纸降解率分别达到12.5%、14.3%。黄色瘤胃球菌可以降解难以降解的纤维如棉花纤维等,Krause等[11]在对瘤胃球菌的多样性研究中,测定了26株瘤胃球菌的基因,结果表明仅有3株瘤胃球菌Y1、LP-9155和AR67基因型相同,在此基础上研究了对纤维的消化能力,结果显示部分瘤胃球菌没有纤维的降解能力,其中Y1菌株纤维消化最高可达86.4%,消化能力和基因型的多样性表明自然界中可能存在较强纤维降解能力的菌株。Mondal等[12]将黄色瘤胃球菌所分泌纤维素酶在大肠埃希菌中克隆表达后对其测定,结果显示重组酶宽pH范围内(3.5~9.0)表现出良好的活性和稳定性,具有高温酶的特性,具有很大的生物技术潜力。
1.4 其他瘤胃纤维降解细菌
1.4.1 溶纤维丁酸弧菌 溶纤维丁酸弧菌(Butyrivibriofibrisolvens)为丁酸弧菌属的模式种,革兰阴性杆菌,严格厌氧菌,单生、成链或丝状体,丝状体可呈螺旋状。研究表明,溶纤维丁酸弧菌的相关纤维降解基因在瘤胃宏基组中纤维降解基因占比较大[13]。Sukharnikov等[14]研究表明,在瘤胃纤维降解中,溶纤维丁酸弧菌发挥着重要作用。Hespell等[15]筛选纯化出30株溶纤维丁酸弧菌,并在此基础上筛选了几株酯酶酶活高的菌株,发现一些菌株含有木聚糖酶和乙酰木聚糖酯酶活性。刘占英等[16]筛选纯化出一株纤维降解细菌,经鉴定为溶纤维丁酸弧菌,并研究了该菌对纤维素降解的性质,结果表明滤纸降解率达到16.81%。溶纤维丁酸弧菌可利用的底物较多,Elianna等[17]在半纤维素(木聚糖、葡聚糖和木葡聚糖)对溶纤维丁酸弧菌的影响研究中,发现溶纤维丁酸弧菌可以有效地发酵木聚糖,而对其他半纤维素(木葡聚糖、葡聚糖)的作用效果较差,证实了溶纤维丁酸弧菌降解半纤维素的生产能力。和瘤胃球菌相似,溶纤维丁酸弧菌降解半纤维素需要生长底物的诱导,Sechovcová等[18]研究了不同碳源对溶纤维丁酸弧菌酶活力的影响,研究结果显示木聚糖诱导溶纤维丁酸弧菌产生β-内切葡聚糖酶和β-木糖苷酶,不同底物主要影响溶纤维丁酸弧菌的蛋白表达量,这些蛋白质参与糖酵解、蛋白质合成和丁酸盐的合成。除此之外,溶纤维丁酸弧菌作为一株益生菌,可以减轻由于弯曲杆菌(Campylobacter)引起的小肠结肠炎等影响[19]。
1.4.2 大肠埃希菌 内蒙古自治区发酵产业节能减排工程技术研究中心和生物发酵绿色制造内蒙古自治区工程研究中心(本课题组)庞建等首次从牛瘤胃中分离出一株可分解纤维素的大肠埃希菌(Escherichiacoli)ZH-4,其菌落为圆形、光滑、湿润、乳状,菌株ZH-4具有内切葡聚糖酶、外葡聚糖酶和β-葡萄糖苷酶的全部活性,这解释了该菌株能有效降解纤维素的原因。此外,研究还发现细胞外酶活性随着培养时间的延长而增加,并能分解玉米秸秆、沙柳等纤维素生产乙醇和氢气[20]。在此基础上,庞建等[21]对大肠埃希菌分泌的纤维素酶进行鉴定,结果表明最适pH为6.0,最适温度为50 ℃,并发现Mg2+可增强内切葡聚糖酶(BcsZ)活性,而Zn2+、Cu2+、Fe3+会抑制BcsZ酶活性,证实大肠埃希菌ZH-4的胞外纤维素降解能力是由BcsZ所致。由于大肠埃希菌的复杂双膜结构导致蛋白的分泌水平较低,通常不产生和分泌纤维素酶。庞建从大肠埃希菌ZH-4的生理表型特征、基因组、转录组和比较基因组学进行分析,提出了一种大肠埃希菌纤维素酶分泌机制[22]。
1.4.3 粪肠球菌 本课题组刘占英等[23]从绵羊瘤胃中分离纯化出一株粪肠球菌(Enterococcusfaecalis),在此基础上研究了该菌对纤维素的降解能力,结果表明该菌分泌的羧甲基纤维素酶活力较高,滤纸降解率达到了24%,可有效降解纤维素。此外,有研究表明,纤维素酶基因也存在于肠球菌,Robert等[24]分离纯化出一株肠球菌,经鉴定纤维分解菌株的最近亲缘关系为粪肠球菌,具有纤维素降解活性。李娅婕[25]在对粪肠球菌源β-葡萄糖苷酶的结构与功能的研究中,将β-葡萄糖苷酶的基因进行了克隆并表达,并对其酶活性进行分析,结果表明,最适pH在8.5~9.0之间,最适温度在40~60 ℃之间。此外,刘占英等[26]通过紫外诱变粪肠球菌,发现菌株对微晶纤维素的利用率提高了9.93%,对玉米秸秆的利用率提高了6.58%。
1.5 主要瘤胃纤维降解细菌特性
由于瘤胃纤维降解细菌丰富的代谢途径,瘤胃中纤维降解细菌有着许多相同的特性和特异性,特异性主要体现在对纤维素类型的选择和代谢产物方面(表1)。相似的特性主要分为四个方面:①大多数瘤胃纤维降解细菌具有变异性,即饲料中的成分不同,细菌数量也会随之变化,如纤维含量低则纤维降解菌的数量会减少。此外,有研究表明,pH的改变对纤维降解细菌数量也有相应的影响[27]。②通常情况下,能够降解纤维素的细菌可以利用半纤维素,而降解半纤维素的菌株不能利用纤维素。③大多数瘤胃纤维降解细菌对于纤维素的降解需要底物的诱导,如黄色瘤胃球菌对半纤维素和果胶的降解程度需要生长底物的诱导。④瘤胃纤维降解细菌一般都为固相微生物,指在降解过程中一般附着于饲料颗粒,黏附阶段时主要通过表面的多糖-蛋白复合物进行黏附,对于黏附草料或饲料的位置主要体现在菌株差异上。

表1 瘤胃内主要纤维降解细菌的特异性[28]Table 1 Specificity of main fiber degrading bacteria in rumen[28]
2 瘤胃纤维降解细菌降解纤维素的过程
瘤胃细菌因其分布广泛和生物量的庞大以及更多的代谢途径,普遍认为瘤胃细菌在纤维降解过程中发挥主要作用。研究表明,纤维素在瘤胃中被利用生成挥发性脂肪酸(VFA)、甲烷等经历两个阶段:第一阶段,瘤胃微生物会分泌纤维素酶将纤维素分解成甲酸盐、琥珀酸盐、H2和CO2等(图1);第二阶段,纤维分解的第一步产物经过各种微生物的利用,转化为丙酮酸,这些菌的各种代谢途径再进一步利用,终产物有丁酸、甲烷、乙酸、丙酸等(图1)[29]。

图1 瘤胃内纤维素降解过程[30]Fig.1 Rumen cellulose degradation process[30]
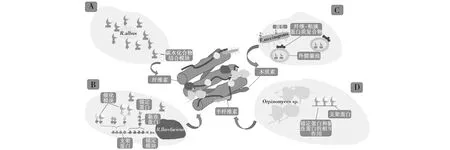
图2 瘤胃微生物降解木质纤维素示意图[41]Fig.2 Schematic representations of cellulose-degrading machineries of rumen microorganisms[41]A:白色瘤胃球菌与含有CBM37的酶一起产生纤维小体复合物;B:黄色瘤胃球菌产生高度结构化的纤维小体复合物;C:产琥珀酸丝状杆菌产生外膜囊泡;D:厌氧根囊鞭菌属产生真菌纤维小体复合物A:R.albus produces cellulosome complex along with CBM37-containing enzymes;B:R. flavefaciens produces a highly structured cellulosome coplex;C:F.succinogenes produces outer membrane vesicule;D:Orpinomyces sp. Produces fungal cellulosome complex
2.1 瘤胃纤维降解细菌对纤维的黏附和降解
纤维物质在瘤胃中的消化是一个连续过程,首先微生物进行黏附,然后进行穿透,产生一系列的纤维素酶,最后实现对纤维的降解和利用[31]。在草料或饲料进入瘤胃时,瘤胃的规律性蠕动使得瘤胃纤维降解细菌快速与新摄入的草料或饲料发生最初的物理性接触[32]。随着附着过程的进行,黏附素的形成使得纤维降解细菌进行更为紧密的特异性附着[33]。瘤胃微生物通过不同方式实现对纤维的黏附和降解见表2。

表2 瘤胃微生物黏附及降解纤维方式和特性Table 2 Rumen microbiota adhesion and fiber degradation modes and characteristics
研究表明,瘤胃球菌黏附和降解纤维过程通过纤维小体进行,不同的黏连蛋白-锚定蛋白群之间存在识别偏好[33]。黄色瘤胃球菌FD-1至少有225个包含锚定蛋白的开放阅读框(ORF)编码多种催化模块[39]。同时,黄色瘤胃球菌可以将一些自由酶和纤维素酶进行结合,进行更有效率的纤维降解。与瘤胃球菌不同,产琥珀丝状杆菌的未知模块可能发挥与CBM或支架蛋白类似的作用,而瘤胃厌氧真菌则是通过假根分泌水解酶来分解纤维素[40]。虽然已知瘤胃纤维的一般分解途径,但瘤胃细菌和真菌纤维小体的组成、模块组装机制和功能作用尚不明确,还有待进一步研究。
2.2 瘤胃纤维降解中微生物之间的互作
由于木质纤维素特殊的结构,仅一种微生物或酶难以将其分解。微生物之间的互作存在多种形式,祁茹等[42]发现瘤胃细菌与瘤胃真菌之间可能存在互补作用,瘤胃厌氧真菌的假根可将植物细胞壁穿透,降解植物颗粒,有利于瘤胃纤维降解细菌的黏附并使得酶反应位点增多。吴宏忠等[43]研究表明,相比瘤胃真菌,瘤胃细菌可以有效的降解纤维,瘤胃细菌与真菌共培养时纤维降解效果最好。Odenyo等[44]发现白色瘤胃球菌与黄色瘤胃球菌共培养时,白色瘤胃球菌会产生一种蛋白质,这种蛋白质会抑制黄色瘤胃球菌的生长。Fondevila等[45]在研究产琥珀酸丝状杆菌、普雷沃氏菌(Prevotellaruminicola)、黄色瘤胃球菌之间的协同和抑制作用时发现,普雷沃氏菌与另外两种纤维降解菌产琥珀酸丝状杆菌、黄色瘤胃球菌共培养时,对饲料纤维的降解明显比产琥珀酸丝状杆菌和黄色瘤胃球菌单独培养时高,并且黄色瘤胃球菌会产生一种抑制物质来抑制产琥珀酸丝状杆菌的生长,从而使得共培养时纤维降解能力的下降。
3 瘤胃纤维降解菌组学研究
3.1 宏基因组学的研究
宏基因组学是以环境样品中全部微生物的DNA为研究对象,提取全部DNA并进行分析来构建基因文库的新颖方法,用于进一步研究微生物的多样性和群落功能[46]。通过对比基因文库,能够筛选出大量瘤胃微生物的功能性基因,Brulc等[47]从海子水牛瘤胃筛选出大量具有纤维素酶活性的糖基水解酶基因。姚健等[48]从牛瘤胃中通过功能驱动法筛选出一个木聚糖酶基因,并测定了其最佳反应温度及pH。Wang等[49]在对Fosmid文库的分析中,筛选出大量的木聚糖和纤维素水解酶相关基因。Stewart等[50]在研究牛瘤胃微生物宏基因组时,通过对比CAZy数据库预测得到235 001个糖苷水解酶、6 834个多糖裂解酶。Gharechahi等[51]通过对骆驼瘤胃进行宏基因组学的研究,发现一种新的内切葡聚糖酶-CelCM3,证明瘤胃中存在大量的未被开发的纤维素酶资源。通过宏基因组学手段,对比基因文库,分析样本中功能性基因的丰富度和表达水平,是研究微生物群落功能的重要手段,对未来的瘤胃微生态研究具有重要意义。
3.2 宏转录组学
宏转录组学是一种提取环境样品中特定时间和空间下的全部RNA,从转录水平上分析基因组转录情况和调控规律的方法,可以有效证明微生物的生长状态和生长规律,研究微生物群落代谢能力[49]。田彦[52]在对奶牛瘤胃微生物的分析中发现瘤胃球菌属、纤维杆菌属和普雷沃氏菌属为主要的纤维降解菌。Meng等[53]研究了麝牛瘤胃中的植物细胞壁降解酶,通过元转录组的方法从2 500多个序列重叠群中鉴定出植物细胞壁降解酶模块,包括糖苷水解酶、碳水化合物酯酶和多糖裂解酶。Comtet-Marre等[54]在转录组学揭示奶牛瘤胃中细菌和真核纤维降解菌群落的研究中,分析鉴定出主要为GH家族组成的12 237个碳水化合物活性酶(CAZymes),大部分CAZymes来源于普雷沃氏菌属、瘤胃菌属和纤维杆菌属,证实这些菌属在纤维降解中的关键作用,此外,作者发现真菌和纤毛原生动物对降解多糖也发挥着重要作用。由于宏转录组学需要以微生物的全部RNA为研究对象,所以对微生物的检测状态具有很高的要求,为减小这种偏差,需要与其他分子手段进行共同分析。
3.3 宏蛋白质组学
宏蛋白质组学是指特定时刻下,环境微生物所表达的所有蛋白。蛋白质组学对揭示环境微生物的活性有重要作用,同时可以进一步揭示瘤胃中纤维降解的机制。Toyoda等[55]从绵羊瘤胃内容物中直接分离并利用宏蛋白质组学的方法鉴定了纤维素结合蛋白(CBPs),CBPs对羧甲基纤维素有内切葡聚糖酶活性,而分离出的5个CBPs中有4个来自产琥珀酸丝状杆菌,表明琥珀酸丝状杆菌显著参与了瘤胃纤维素的降解。Deusch等[56]在研究不同日粮和微环境对瘤胃微生物菌落结构和功能的影响中,通过蛋白质组学进行分析发现大多数表达的糖苷水解酶以GH57和GH2两个家族为主。相比宏基因组学,宏蛋白质组学不仅可以直接获得微生物基因的表达情况,而且可以深入解析微生物间的相互作用,此外,宏蛋白质组学在特殊蛋白质的开发、生态元素循环等研究领域也有着重要作用。
4 瘤胃纤维降解细菌的开发利用
反刍动物种类、日粮和瘤胃微生物、饲料消化率之间存在着动态变化关系,对瘤胃纤维降解细菌的降解过程和瘤胃微生物区系进行分析,对提高饲料利用率和降低饲养成本有着重要作用。于满满等[57]优化了氨化秸秆与苜蓿组合,以此增强瘤胃内主要纤维降解菌的活性,20%苜蓿干草和80%的氨化秸秆组合有效提高了产琥珀酸丝状杆菌和黄色瘤胃球菌的活性,促进瘤胃微生物的发酵。王超等[58]研究了绵羊体外瘤胃发酵全混合饲料对纤维降解和瘤胃纤维降解细菌数量的影响,结果表明,发酵全混合饲料组挥发性脂肪酸生成量显著高于饲喂未发酵全混合饲料组(P<0.05)。此外,瘤胃纤维降解细菌在生产丁酸和生物燃料方面也具有重要作用,黄恒猛[59]研究了溶纤维丁酸弧菌发酵产丁酸和乙醇,为工业生产提供了方案。除此之外,粪肠球菌有着提高瘤胃微生物活性、改善瘤胃pH环境的作用,开发具有纤维素降解能力的粪肠球菌对饲料的降解效率和增强反刍动物免疫力有着重要作用。
5 展 望
前人对细菌附着纤维、酶的组成和特性以及菌群的相互关系等方面进行了大量的研究,结果表明,在复杂的瘤胃微生态中,细菌被认为是纤维降解的主体,具有较强的纤维降解能力,瘤胃纤维降解细菌经过附着、黏附、降解和利用实现对纤维的利用,另一方面,瘤胃纤维降解细菌之间的相互作用使得纤维降解更为有效。由于特殊的瘤胃生态环境、微生物群体之间复杂的相互关系、以及研究方法的限制等问题,当前仍有三个亟需解决的问题:①对瘤胃纤维降解细菌对纤维的分解机制尚不明确,瘤胃纤维降解细菌产生的类似纤维小体复合物的结构、作用机理等问题还需通过分子生物学进一步研究。②由于特殊的瘤胃环境、体内实验和瘤胃微生物分离的难度以及对瘤胃内纤维降解细菌在纤维降解中的贡献尚不明确,对于菌种之间的相互协作和相互制约关系研究仍存在不足之处。③瘤胃微生物种类繁多、数量巨大,瘤胃内仍存在大量的纤维素酶资源未被开发,通过宏组学手段开发利用新的特异性强、酶活力高的纤维素酶资源对畜牧业和工业生产具有重要意义。随着研究的深入,对瘤胃纤维降解细菌的认识将更加深刻,进一步研究纤维降解细菌的降解机理、菌群之间的竞争和协同关系,开发新的酶源,为纤维降解机制的明确和提高反刍动物饲料利用率提供参考。
