林分因子对神木垒不同森林群落林下植物多样性的影响
2023-12-05何宇婷刘鸿翔郝建锋
王 琴,王 芳,何宇婷,周 宁,刘鸿翔,郝建锋,2*
(1 四川农业大学 林学院,成都 611130;2 水土保持与荒漠化防治重点实验室,成都 611130)
森林植物多样性是森林生物多样性的核心,其经济、社会和生态价值较高[1]。在生态文明建设的背景下,如何提高森林植物多样性,是实现森林可持续发展中所面临的一个重要问题。林下植被层包含了森林90%以上的植物种类,竞争激烈,对于生境变化更敏感,其凋落物的贡献量大,细根发达。林下植物多样性是森林植物多样性的重要部分,对森林的结构与功能的维持发挥了重要作用[2-3]。探究影响林下植物多样性的主要因子,对于提高林下植物多样性,充分发挥林下植物生态功能具有重要意义。
大量研究表明,林下植物多样性受海拔、地形、土壤等环境因素的影响[4-8],也有部分学者认为林下植物多样性可能会受到群落类型[5,9-12]、群落结构[13-15]等群落本身特性的影响。群落所处的环境条件一般比较稳定,难以进行人为调控,因此研究群落本身特性对林下物种多样性的影响对森林经营管理措施的制定更具指导意义。不同森林群落由于其乔木层的树种组成差异,会对林下植物多样性产生影响[9-12]。有研究发现,针叶林林下植物多样性低于阔叶林,但针叶纯林林下植物多样性要高于针阔混交林[11],而范川等[10]则认为混交林群落林下植物多样性高于纯林。群落类型对林下植物多样性的影响也可能通过群落结构发生作用。胡文杰等[13]认为马尾松群落上层林分垂直结构与灌木层物种多样性呈正相关;闫东锋等[14]研究发现,群落冠层结构会通过林下光环境从而影响灌木层物种多样性。
林分因子是森林群落结构的重要组成部分[16],近年来的大量研究均发现林分因子对林下植物多样性有影响[16-22],但不同研究中影响林下植物多样性的主要林分因子不尽相同。朱媛君等[17]对张北杨树人工林的研究认为,林分密度是影响林下草本层物种多样性的主要林分因子;陈笑等[18]在青海省互助北山林场的研究认为平均胸径是影响草本层物种多样性的主要林分因子;张林等[19]在北京山区的研究则认为林分密度与郁闭度均对林下草本层的物种多样性起主导作用。林分因子对草本层与灌木层的影响程度也不同,李婷婷等[16]认为林分因子对灌木层的物种多样性无显著影响,但对草本层物种多样性的影响较大;周润惠等[20]、苏天成等[21]、陈聪琳等[22]的研究发现林分因子对灌木层和草本层的物种多样性均有显著影响。林分因子对林下植物多样性的影响机制较为复杂,在不同环境条件下往往会呈现不同的结果,林分因子对林下植物多样性的影响还需进一步探讨。
神木垒景区隶属于大熊猫国家公园,是国家重点生态保护区,位于全球生物多样性热点地区之一。神木垒植被类型丰富,主要为针叶纯林、针阔混交林和阔叶混交林,然而各森林群落的植物多样性之间是否存在差异、哪种森林群落最有利于当地生物多样性的发展以及林分因子是否是造成各群落间林下植物多样性差异的重要因素等问题并不明确。因此,文章以神木垒林区内的5种主要森林群落为研究对象,对不同森林群落林下植物多样性进行比较与分析,并探究林分因子对林下植物多样性的影响,以期得到影响林下植物多样性的主导林分因子,为神木垒及其他类似区域的森林经营管理提供理论依据。
1 材料和方法
1.1 研究区概况
研究区位于四川省宝兴县硗碛藏族乡西部神木垒风景区(30°41′-30°42′N,102°41′-102°43′E),海拔2 500~2 700 m,属亚热带半湿润气候区,为亚热带向暖温带过渡的湿润季风气候带,山地气候类型,年平均气温8.9 ℃,年降水量776.5 mm,年日照时数1 400 h,土壤以山地黄壤为主。研究区内分布的主要森林群落为云杉林群落、丽江云杉林群落、红杉林群落、针阔混交林群落、阔叶林群落。研究区内植物种类丰富,主要乔木有云杉(Piceaasperata)、丽江云杉(Picealikiangensis)、红杉(Larixpotaninii)、冷杉(Abiesfabri)、亮叶桦(Betulaluminifera)等;主要灌木有银叶杜鹃(Rhododendronargyrophyllum)、粉叶小檗(Berberispruinosa)、峨眉蔷薇(Rosaomeiensis)等;主要草本植物有川西鳞毛蕨(Dryopterisrosthornii)、蛇莓(Duchesneaindica)、早熟禾(Poaannua)及苔藓等。
1.2 样地设置与植被调查
在神木垒林区的调查研究范围内,按照方精云等[23]的研究方法,采用典型样地法,在全面踏查的基础上选取立地条件基本相似、林相整齐、林层丰富、具有代表性的Ⅰ(云杉林)、Ⅱ(丽江云杉林)、Ⅲ(红杉林)、Ⅳ(针阔混交林)、Ⅴ(阔叶林)5种林地各4个样地,面积为20 m×30 m,共计20个,面积总计12 000 m2。在各样地内采用相邻格子法设置10 m×10 m乔木样方6个,共计120个;沿样地对角线设置5 m×5 m灌木样方6个,共计120个;1 m×1 m草本样方12个,共计240个。在各样地内,对乔木进行每木检尺。按乔木层、灌木层和草本层划分层次进行测定、统计。记录乔木的树种名称、胸径、树高、枝下高、冠幅、各树种株数,记录灌木和草本的植物种名、高度、冠幅、株数(丛数)。样地基本情况见表1。
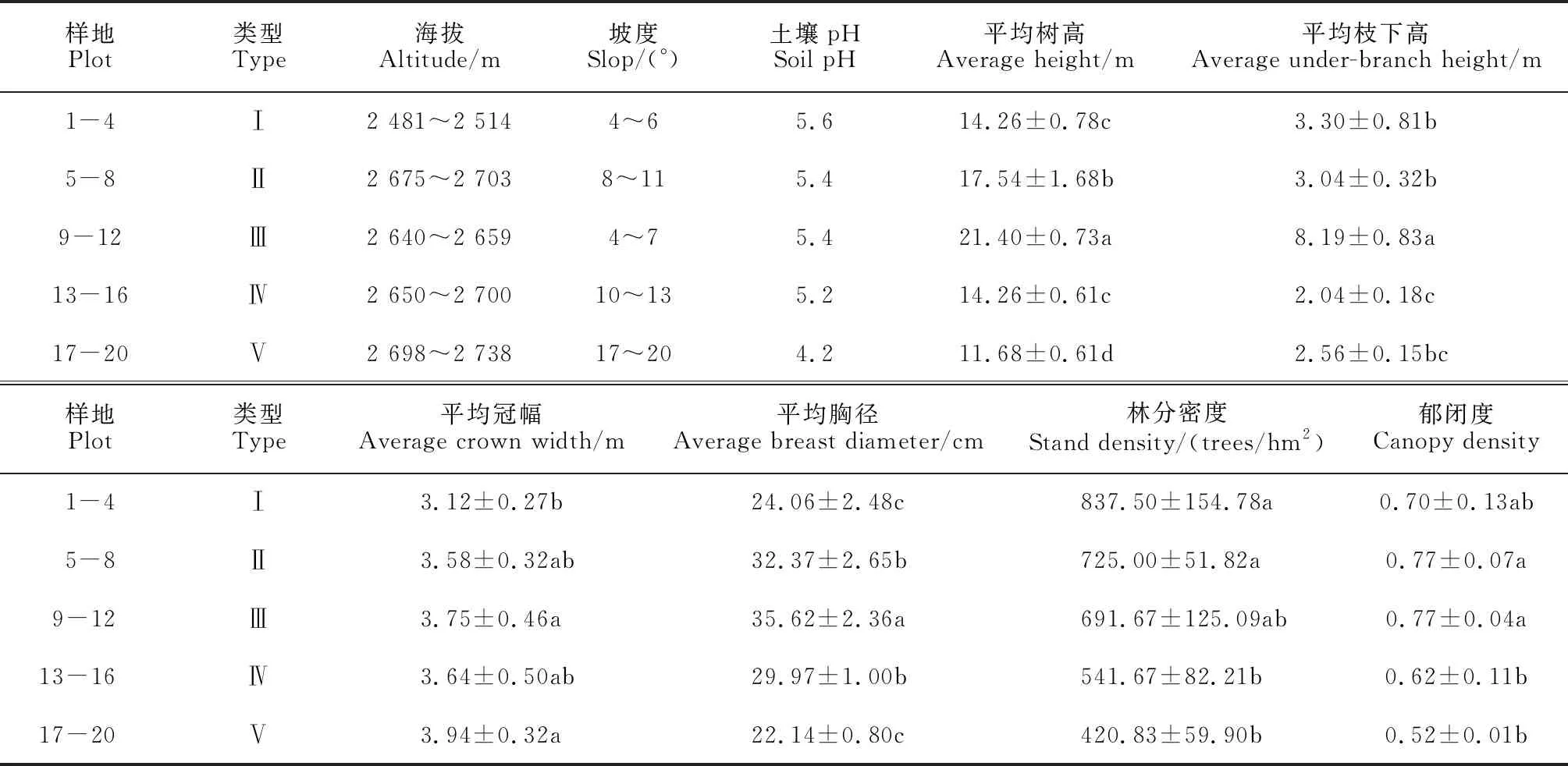
表1 样地概况
1.3 数据处理
采用以下公式综合测度和分析群落物种多样性特征。
重要值=(相对密度+相对盖度+相对频度)/3
(1)
丰富度指数(D):D=S
在对于企业信用认定方面,由于区块链网络具有去中性化的特点。相对于传统的信任机制,其信任模式并非建立在传统第三方信用机构而是通过技术进行。区块链技术的“去信任”机制能使原本由政府和机构审查提升为全社会乃至全世界共同治理,其中去中心化和不可篡改的特点在保证数据源客观准确的同时降低了跨境监管成本,增强了参与者的信心,为跨境电商中各交易主体在跨境交易中提供可靠的信任基础。
(2)
Simpson指数(优势度指数,H′):
(3)
Shannon-Wiener多样性指数(H):
(4)
Pielou均匀度指数(Jsw):
(5)
式中:Pi=ni/n,ni为第i种的个体数;n为所有种的个体总数;i=1,2,3,…,S;S为物种数。
所有数据采用Excel 2019进行整理,用SPSS 27.0统计软件对灌草层物种多样性指数和林分因子进行单因素方差分析(one-way ANOVA),用最小显著差异法(LSD)检验差异显著性(P=0.05);用Canoco 5.0对物种多样性和林分因子进行冗余分析(RDA),其余数据均用Origin 2021作图。
2 结果与分析
2.1 不同森林群落林下物种组成及重要值
2.1.1 不同森林群落林下物种组成

F. 科;G. 属;S. 种。图1 不同森林群落灌草层物种组成F. Family; G. Gensus; S. Species.Fig.1 Species of shrub and herb layers in different forest communities
2.1.2 不同森林群落林下物种重要值
对草本层与灌木层各物种的重要值进行统计,结果(表2)显示:不同森林群落草本层与灌木层的优势种及重要值前5的物种组成不同。群落Ⅲ、群落Ⅳ的草本层以蔷薇科(Rosaceae)和禾本科(Poaceae)植物为主,群落Ⅴ草本层主要以蕨类和蔷薇科的草本植物为主。

表2 不同森林群落灌草层优势种重要值
蛇莓在每个群落中的重要值都较高,为整个调查区草本层的优势种。群落Ⅰ、Ⅱ、Ⅲ灌木层的优势种均为峨眉蔷薇与粉叶小檗。群落Ⅲ灌木层中峨眉蔷薇占据绝对优势。群落Ⅳ与群落Ⅴ灌木层优势种均为银叶杜鹃。杜鹃花科(Ericaceae)植物在群落Ⅳ和群落Ⅴ的灌木层中占据绝对优势。
2.2 不同森林群落物种多样性比较
单因素方差分析(图2)表明:群落Ⅰ与群落Ⅱ灌木层的H值、H′值之间无显著差异(P>0.05),但显著(P<0.05)高于其他3个群落。群落Ⅰ、群落Ⅳ灌木层的D值较高,显著高于其他3个群落。群落Ⅱ灌木层的JSW值最高,显著高于群落Ⅳ和群落Ⅴ。

图2 不同森林群落灌木层与草本层物种多样性指数Fig.2 Species diversity indexes of shrub layer and herb layer in different forest communities
由此可见,云杉林群落灌木层的物种多样性最高,丽江云杉林群落次之。群落Ⅱ草本层的4个多样性指数值均最大,H值显著高于其他4个群落,H′值显著高于群落Ⅳ,D值显著高于群落Ⅳ与群落Ⅴ,JSW值显著高于群落Ⅰ与群落Ⅳ。由此可见,丽江云杉林群落草本层的物种多样性最高。总的来看,云杉林群落与丽江云杉林群落林下植物多样性较高,针阔混交林群落与阔叶林群落林下植物多样性均较差。
2.3 林分因子与物种多样性的冗余分析
以草本层和灌木层的4个多样性指数作为响应变量,以6个林分因子作为解释变量进行RDA排序(表3),结果表明:灌木层和草本层的第1轴和所有轴的蒙特卡洛置换检验结果P值均小于0.05,RDA排序结果可靠。RDA在第一轴和第二轴分别解释了灌木层多样性特征数据总方差的74.70%和6.12%,解释了草本层多样性特征数据总方差的58.40%和1.15%。由此可见,前两轴能较好地反映灌木层和草本层物种多样性与林分因子之间的关系,且主要由第一轴决定。

表3 林分因子与灌草层物种多样性的冗余分析结果及蒙特卡洛置换检验结果
灌木层的RDA双序图(图3,A)表明,RDA 1轴主要反映了平均枝下高、平均冠幅、平均胸径、平均树高的变化,RDA 2轴主要反映了林分密度、郁闭度的变化。蒙特卡洛置换检验(表3)表明,平均枝下高、林分密度均显著影响灌木层物种多样性,对变异的解释率分别为71.5%、15.4%。平均枝下高与林分密度是影响灌木层物种多样性的主要林分因子,平均枝下高与灌木层的D值、H值、H′值呈负相关关系,林分密度与灌木层4个多样性指数均呈正相关。草本层的RDA双序图(图3,B)表明,RDA 1轴主要反映了林分密度、郁闭度、平均冠幅的变化,RDA 2轴主要反映了平均枝下高的变化。蒙特卡洛置换检验(表3)表明,仅平均枝下高显著影响草本层物种多样性,对变异的解释率为75%。平均枝下高是影响草本层物种多样性的主要林分因子,平均枝下高与草本层H值、H′值、JSW值呈正相关。
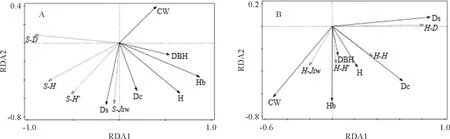
S-.灌木层;H-.草本层。H. Shannon-Wiener多样性指数;H′. Simpson优势度指数;D.物种丰富度指数;Jsw. Pielou均匀度指数。图3 林分因子与灌草层物种多样性的RDA排序图S-. Indexes of shrub layer; H-. Indexes of herb layer. H. Shannon-Wiener diversity index;H′. Simpson dominance index; D. Species richness index; JSW. Pielou evenness index.Fig.3 RDA ordination diagram of species diversity and forest factors
3 讨 论
3.1 不同森林群落对林下植物物种组成的影响
林下植物物种组成是森林群落的重要特征之一[16]。本研究中不同森林群落林下植物物种组成均不同。蛇莓在每个群落中的重要值都较高,为整个调查区草本层的优势种。3个针叶林群落灌木层的优势种均为峨眉蔷薇与粉叶小檗,但峨眉蔷薇在红杉林群落中占据绝对优势。不同乔木树种的生物学特性可能会导致林下光环境的差异,从而影响林下植物物种组成[3]。云杉、丽江云杉均为耐阴树种,其自然整枝能力差,导致林分平均枝下高较低,林下透光少,更适合阴生灌木的生长,使得云杉林群落和丽江云杉林群落灌木层植物科属种组成较丰富;红杉为喜光树种,其自然整枝能力强,林分平均枝下高较高,林下透光条件好,因此在红杉林群落中,阴生灌木(如粉叶小檗、木帚栒子等)的生长受到限制,使得峨眉蔷薇可以占据较大的生态位,在红杉林群落灌木层中占据绝对优势[24],从而导致红杉林群落灌木层植物科属种组成较为简单。杜鹃花科植物在针阔混交林群落与阔叶林群落的灌木层中占据绝对优势,可能是因为这2个群落的土壤酸性较高,对其他灌木的生长有抑制作用[25],而杜鹃花科植物喜酸性土[26],更占优势。
3.2 不同森林群落对林下植物多样性的影响
群落类型对森林群落的植物多样性有着重要影响[5],在本研究中,不同森林群落林下植物多样性之间有显著差异,但物种多样性较高的群落为云杉林与丽江云杉林,均为针叶纯林,而针阔混交林群落、阔叶林群落林下植物多样性均较低,此结果符合Barbier等[11]针叶纯林林下植物多样性大于针阔混交林的观点,但与范川等[10]的研究结果不同。阔叶林群落林下植物多样性较低可能是因为其林分发育尚未成熟,林分密度小、郁闭度低,林下土壤酸性较高,不利于林下植物的生长[4,25]。针阔混交林群落林下植物多样性低则可能是因为其平均枝下高过低,加上混交冷杉斜上伸展的侧枝的遮挡,导致林下光照不足、林下空间狭小,不能满足林下植物生长所需[24,27-28]。各群落草本层物种多样性均高于灌木层,表明草本层对于森林物种多样性的贡献更大,与李婷婷等[16]的观点相同。因此应更加注重草本层物种多样性的提升。
3.3 林分因子对林下植物多样性的影响
本研究中,林分因子对灌木层和草本层的物种多样性均有显著影响,与前人研究结果[20-22]相同,但分别影响灌木层和草本层的主要林分因子与前人研究结果不尽相同。冗余分析结果表明:平均枝下高和林分密度是影响林下植物多样性的主要因子。其中平均枝下高对灌木层和草本层的物种多样性均有显著影响,林分密度仅对灌木层物种多样性有显著影响。平均枝下高与灌木层物种多样性呈负相关关系,此结果与苏天成等[21]的观点相同。平均枝下高会影响林下透光率,林下的光照条件对林下植物组成[3]和植物物种多样性[15,27]均有重要影响,当光照较充足时,林下喜光植物生长旺盛,而阴生植物的生长会受到限制。由于本研究调查区海拔较高,日照较强,平均枝下高最高的红杉林群落,其林下充足的光照反而抑制了阴生灌木(如粉叶小檗、木帚栒子等)的生长[29],而峨眉蔷薇对光照的需求更高,生长不受抑制,占据较大的生态位,在灌木层占据绝对优势,导致其灌木层物种多样性低[24]。平均枝下高与草本层物种多样性呈正相关,此结果与陈聪琳等[22]的研究结果相同,与李婷婷等[16]、朱媛君等[17]的研究结果则相反。可能是因为李婷婷等[16]的研究中林分郁闭度较大,其草本层多为喜阴植物,较高的平均枝下高会使草本层受到的光照更多,反而不利于其物种多样性的增加[15]。而本研究区中草本层多为喜阳或半喜阳植物,且平均枝下高可能还会通过影响灌木层物种多样性从而间接影响草本层物种多样性。平均枝下高越高,灌木层物种多样性越低,灌木层植物生长不佳,却为草本层植物的生长提供了较大的空间,同时增加了灌木层之下的透光率,使得草本层植物能得到充足的光照,从而有利于草本层物种多样性的增加[27-28]。因此红杉林群落虽灌木层物种多样性差,但草本层物种多样性相对较高。
林分密度可通过影响林下光照条件或土壤理化性质来影响林下植物多样性[30]。适当增加林分密度有利于林下植物多样性的增加,过高的林分密度则会限制林下植物多样性的发展[31]。本研究中,林分密度对灌木层的物种多样性有显著影响,但与草本层的物种多样性没有显著相关关系,与张林等[19]的研究结果不同。林分密度对草本层的物种多样性没有显著影响,可能是因为本研究区内草本层物种多样性整体较高,对林分密度差异的响应不明显[22]。林分密度与灌木层物种多样性呈正相关,与金锁等[31]的研究结果不同,表明神木垒各群落的林分密度均未达到限制林下植物多样性的阈值,对于林分密度较小、灌木层物种多样性较低的针阔混交林群落与阔叶林群落,适当增大其林分密度可能有利于灌木层物种多样性的增加。
4 结 论
(1)神木垒5种主要森林群落林下植物多样性存在差异,林下植物多样性较好的群落为云杉林群落与丽江云杉林群落,灌木层多样性最高的群落为云杉林群落,草本层多样性最高的群落为丽江云杉林群落。
(2)林分因子对林下植物多样性有显著影响,灌木层物种多样性主要受平均枝下高与林分密度的影响,草本层物种多样性主要受平均枝下高的影响。红杉林群落较高的平均枝下高可能是导致其灌木层物种多样性较低的重要因素;针阔混交林群落与阔叶林群落林分密度较小、平均枝下高较低,可能是导致其林下植物多样性较差的重要原因。
(3)针阔混交林群落、阔叶林群落林下植物多样性较差,可考虑适当增大林分密度以提高物种多样性。由于阔叶林群落林分发育尚未成熟,其林下植物多样性水平可能还未达到群落最佳水平,因此阔叶林群落林下植物多样性较差的结果具有一定局限性,还需通过后续调查和研究进一步探索。
