滇池藻胆蛋白时空分布特征及其影响因素分析
2023-11-30刘平魏薇杨俊晖李建文左李美孟睿段平洲焦立新
刘平,魏薇,杨俊晖,李建文,左李美,孟睿,段平洲*,焦立新
1.环境基准与风险评估国家重点实验室,中国环境科学研究院
2.昆明市城市排水监测站
藻胆蛋白是一类具有生物活性的天然色素,众多研究表明,藻胆蛋白不仅可以作为食品和化妆品工业的光敏剂或色素添加剂,还具有抗癌、抗氧化、降血压血脂等诸多药用价值,因此受到了广泛的关注[1-3]。藻胆蛋白主要存在于蓝藻、红藻、隐藻和少数甲藻中,起到吸收光照、促进光合作用的效果[4-5]。已知的藻胆蛋白主要分为4 类[6-7]:藻红蛋白(phycoerythrin,PE)、藻红蓝蛋白(phcoerycyanin,PEC)、藻蓝蛋白(phycocyanin,PC)和别藻蓝蛋白(allophycocyanin,AP)。
滇池是云南省面积最大的高原淡水湖泊,由于地处昆明市城区,承接了大量生活污水和工农业废水,导致其富营养化问题突出[8-10]。文献报道滇池北部区域藻密度为1 909.00×104~51 009.66×104个/L,南部区域为1 342.67×104~25 130.97×104个/L,以蓝藻和绿藻为绝对优势种群[11-12]。2010—2018 年,平均每年从滇池打捞蓝藻藻泥约6 万t(鲜质量)。有研究表明滇池蓝藻的藻胆蛋白含量丰富,其含量比养殖的螺旋藻高出近1 倍[13-14]。因此通过回收利用蓝藻藻泥提取藻胆蛋白,既能达到治理环境污染目的,又可以减轻固废处理负担,产生经济效益[15-16]。
然而现阶段对滇池藻胆蛋白的分布特征尚没有全面的研究,这影响了对蓝藻的治理和综合利用。目前工业界提取分离藻胆蛋白的主要原材料是藻红蛋白和藻蓝蛋白,因此笔者基于2022 年4 次藻类采样和藻胆蛋白的检测结果,探究了滇池藻红蛋白和藻蓝蛋白的时空分布规律,以及其与水体理化和生物因子的相关性,阐述藻胆蛋白积累的环境驱动因子,以期为滇池微藻的开发利用提供理论依据。
1 研究区与研究方法
1.1 研究区概况
滇 池 流 域 位 于 云 贵 高 原 中 部(102°29′E~103°01′E,24°29′N~25°28′N),地处云南省昆明市南部。滇池流域面积为2 920 km2,湖体面积为309.5 km2,湖体平均水深为5.3 m,湖体库容为15.6 亿m3。滇池分为草海和外海,二者相互连接却又几乎不可互换,其中外海为滇池的主体。滇池多年平均入湖水资源量为9.7 亿m3,2013 年实施了牛栏江—滇池补水工程,年均补水量为5.6 亿m3。常年汇入滇池的河流有35 条,其中,面积大于100 km2的有盘龙江、洛龙河、宝象河(新宝象河)等10 条;面积介于50~100 km2的有新运粮河、东白沙河(海河)、马料河、南冲河、淤泥河5 条;面积介于10~50 km2的有老运粮河、采莲河、大清河、枧槽河、广普大沟、金汁河、中河(护城河)、古城河8 条;面积小于10 km2的河流有乌龙河、大观河、西坝河等12 条。
1.2 采样与分析方法
1.2.1 藻体采样
在滇池湖体设置32 个点位〔图1〕,其中滇池南、断桥、白鱼口、海口西、罗家营、灰湾中、草海中心、观音山西、观音山中、观音山东为国控点位,将滇池分为草海、外海北部、外海中部和外海南部4 个湖区。分别于2022 年2 月(冬季)、5 月(春季)、8 月(夏季)和11 月(秋季)进行藻体采样。采样时使用GPS 系统定位,用2.5 m 长的PVC 管采集整个水柱,混合均匀,每个采样点设3 个平行。每个样品采集至少1.2 L,所有样品于4 ℃状态下及时送至实验室。每个水样使用低速大容量多管离心机在6 000 r/min 下离心20 min,收集鲜藻。取1 L 水样过0.45µm 的GF/C 玻璃纤维滤膜,将过滤出的藻体和GF/C 滤膜冷冻干燥。
1.2.2 藻胆蛋白浓度测定
1.2.2 .1 藻胆蛋白的提取
藻体用磷酸盐缓冲液(PBS)溶液,于-20 ℃反复冻融3 次,然后放入离心管置冰浴中,使用细胞破碎机分别以120 W 的功率破碎藻体,以超声10 s、间歇10 s 的方式进行处理,进行40 次。离心后取上清液,向得到的藻胆素提取液中加入适量甲醇和浓盐酸(体积比为95∶5),记录稀释倍数,待测[17]。
1.2.2 .2 吸收光谱和标准曲线
配制0.5 mg/mL 的藻红蛋白、藻蓝蛋白标准溶液,在250~700 nm 波长内扫描测定溶液的吸光度(A),得到藻红蛋白、藻蓝蛋白溶液的吸收光谱。准确配制0.01、0.02、0.03、0.04、0.05、 0.10、0.15、0.20 mg/mL 的藻红蛋白和藻蓝蛋白标准系列溶液,每个浓度作3 个平行样,分别测定标准样品在560 和620 nm 的吸光度。以测定的A为纵坐标,以浓度为横坐标,绘制标准曲线[18]。
1.2.2 .3 藻胆蛋白定量
藻红蛋白、藻蓝蛋白浓度计算参考Beer 等[19]的方法测定。藻胆蛋白的吸光度均使用紫外可见分光光度计(岛津UV-2501)测定。提取的藻红蛋白、藻蓝蛋白在阳光下可显现出荧光特性,但光照可促使其分解,因此试验过程应在避光、低温条件下进行。同时,藻胆蛋白作为一种营养物质,细菌可使其快速降解,因此本试验在无菌条件下进行。
1.2.2 .4 稳定性和重现性试验
准确配制0.1 mg/mL 藻红蛋白、藻蓝蛋白样品溶液10 份,按上述藻胆蛋白定量方法,分别测定样品溶液在560 和620 nm 的吸光度[20]。
1.2.2 .5 回收率试验
取10 mL 具塞试管7 支,分别准确加入样品溶液1.0 mL,其中1 号试管不加标准溶液,2~4 号试管中各加0.1 mg/mL 藻红蛋白、藻蓝蛋白标准溶液1 mL,5~7 号试管中各加入0.1 mg/mL 藻红蛋白、藻蓝蛋白标准溶液2 mL,加空白液补充至4 mL,按上述藻胆蛋白定量方法,分别测定藻红蛋白、藻蓝蛋白溶液在560 和620nm 的吸光度,计算回收率[21]。
1.3 数据分析
利用中国科学院资源环境科学数据中心(http://www.resdc.cn/)2015 年中国土地利用遥感监测数据提取滇池水体边界,分辨率为30 m×30 m。借助ENVI5.1、ArcGIS10.2 软件进行空间裁剪、掩模、重分类,将采集监测的藻胆蛋白数据导入影像,建立藻胆蛋白浓度时间和空间序列栅格数据集,揭示滇池藻胆蛋白的时空分布规律。
测定各采样点平行样,取平均值。使用SPSS22.0 软件进行数据统计分析,使用Origin2018软件绘图。采用方差(ANOVA)分析和最小显著性差异(LSD)法测验环境因子与藻胆蛋白浓度的时空差异,并通过Pearson 相关性分析研究藻胆蛋白浓度与环境因子的关系(P>0.05 时相关性不显著,P<0.05 时相关性显著,P<0.01 时相关性极显著)。
2 结果与分析
2.1 藻胆蛋白的检测方法
由藻红蛋白、藻蓝蛋白标准溶液在250~700 nm 波长的吸光光谱〔图2(a)〕可知,藻红蛋白在可见光范围内有3 个主要的吸收峰,分别在460、520 和560 nm 处,其中在560 nm 处的吸收峰最强,因此采用该波长作为藻红蛋白的测定标准[22]。藻蓝蛋白只有1 个明显的吸收峰,位于620 nm,因此选定此峰为藻蓝蛋白的定量标准。以不同浓度的藻红蛋白、藻蓝蛋白标准溶液分别在560 和620 nm 下测定吸光度,均得到了良好的线性规律〔图2(b)〕。经过多个样品重复测试,得到藻红蛋白质量浓度为93.17%±2.34%,藻蓝蛋白质量浓度为91.48%±1.77%,说明该方法的重现性良好。同时,藻蓝蛋白和藻红蛋白的回收率分别达到97.63%和95.47%,说明方法的回收率较高,可用于藻胆蛋白的定量测试和分析。
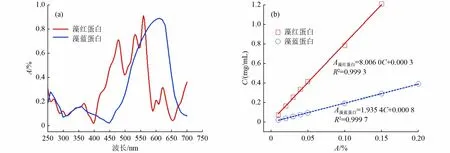
图2 藻胆蛋白的吸光波谱和标准曲线Fig.2 Absorption spectrum and standard curve of phycobiliprotein
2.2 滇池藻胆蛋白时空分布规律
2.2.1 滇池藻红蛋白时空分布规律
滇池湖体32 个点位的4 次藻红蛋白分析结果如图3 所示。在时间尺度上,整体上夏季和秋季的藻红蛋白浓度明显高于春季和冬季;从平均值来看,表现为夏季〔(6.89±2.99)µg/L〕>秋季〔(5.24±3.61)µg/L〕>春季〔(3.85±1.93)µg/L〕>冬季〔(3.83±2.43)µg/L〕。虽然冬季的藻红蛋白平均浓度最低,但是在局部区域却依然出现较高的藻红蛋白浓度,导致其标准偏差较大,原因可能是滇池在11 月气温升高,藻类在适宜条件下恢复生长和生物活性。
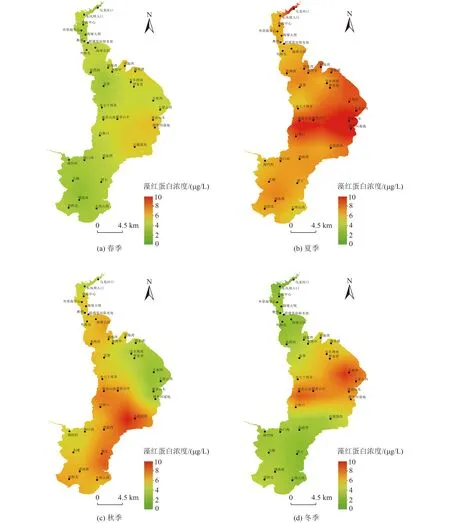
图3 滇池藻红蛋白浓度时空分布Fig.3 Spatial and temporal distribution of phycoerythrin concentration in Dianchi Lake
从空间分布上,外海北部的藻红蛋白浓度显著高于其他区域,具体表现为外海北部〔(6.25±2.81)µg/L〕>外 海 中 部〔(4.99±2.16)µg/L〕>外 海 南 部〔(4.66±2.09)µg/L〕>草海〔(3.94±2.67)µg/L〕。可以看出藻红蛋白的分布呈现明显的南北低、中间高特征:春季时虽然整体浓度较低,但在外海中东部区域(观音山东、捞鱼河湿地、古滇国西)呈现出了较高的浓度分布;夏季滇池藻红蛋白的浓度较高,局部点位达到了14.13 和15.76 µg/L,浓度最高的区域依然集中在外海北部和中部,其中东部沿岸区域浓度高、分布广,而西部区域集中在观音山西、白鱼口等点位检测出了较高浓度的藻红蛋白;秋季多个区域由于藻类死亡和活性抑制,藻红蛋白浓度快速降低,但在西南部(古滇国西)和中西部(观音山西)还具有较高的浓度;冬季时,外海北部和南部区域已经呈现极低的藻红蛋白浓度,但在外海中部区域浓度依然较高。
2.2.2 滇池藻蓝蛋白时空分布规律
滇池湖体藻蓝蛋白的浓度分析结果如图4 所示。可以看出藻蓝蛋白和藻红蛋白的时空分布规律类似,但藻蓝蛋白的浓度明显更高,这归因于滇池湖体较高的蓝藻种群密度[23-24]。与藻红蛋白相比,夏秋季藻蓝蛋白的浓度与春冬季的差距更大。从时间上分析,表现为夏季〔(55.10±22.37)µg/L〕>秋季〔(38.83±25.72)µg/L〕>春季〔(30.37±13.66)µg/L〕>冬季〔(16.54±11.30)µg/L〕,其中夏季最高浓度可以达到106.42µg/L(观音山东)和107.67 µg/L(捞鱼河湿地),而春冬季的浓度均在50 µg/L 以下,说明藻蓝蛋白浓度主要依靠蓝藻的生物量和生物活性,在夏秋季达到高峰[25]。藻胆蛋白是藻类的捕光色素蛋白,其浓度高低与其光合作用密切相关,滇池在夏秋季光照强度较高,这也是藻胆蛋白浓度集中升高的重要原因。从空间上分析,藻蓝蛋白的产出依然以外海中部为主,其四季平均浓度为外海中部〔(44.38±28.65)µg/L〕>外海北部〔(34.66±21.75)µg/L〕>外海南部〔(33.64±19.30)µg/L〕>草海〔(27.87±15.85)µg/L〕,同样呈现南北低、中间高的规律,因此外海中部区域可以作为藻胆蛋白提取和回收的重点区域。
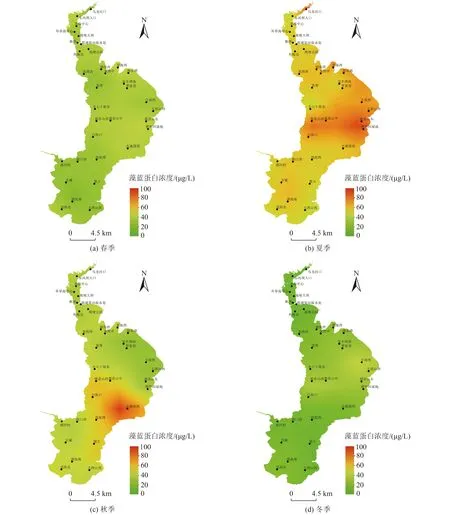
图4 滇池藻蓝蛋白浓度时空分布Fig.4 Spatial and temporal distribution of phycocyanin concentration in Dianchi Lake
2.3 滇池藻胆蛋白的影响因素分析
2.3.1 藻胆蛋白与水环境因子的相关性
基于藻胆蛋白时空差异性分析结果, 对滇池10 个国控点位的藻红蛋白、藻蓝蛋白浓度分布和环境因素进行了Pearson 相关性分析,结果见图5。全湖的藻密度和叶绿素与总磷(TP)浓度、pH 及五日生化需氧量(BOD5)存在很强的线性相关性(P<0.01),与溶解氧(DO)浓度和透明度存在较强的线性相关性(P<0.05),说明滇池藻类繁殖增长主要受到TP 浓度和BOD5限制[26-27],藻密度的增加反过来影响水体的pH、DO 浓度和透明度[28]。藻蓝蛋白浓度与TP浓度(相关系数为0.731 8,P<0.01)、BOD5(相关系数为0.594 1,P<0.05)存在很强的线性相关性,而藻红蛋白浓度与湖水理化指标(营养盐浓度、DO 浓度、pH、透明度)的相关性都较弱,说明滇池藻体主要以产藻蓝蛋白的蓝藻为主,红藻、硅藻等藻红蛋白生产者的种群密度很低[11-12]。同时,藻蓝蛋白、藻红蛋白的浓度比(藻蓝/藻红)与湖水的理化指标相关性也很低,说明二者的波动趋势存在较大差别,不同藻体的种群密度间没有显著的相关性。
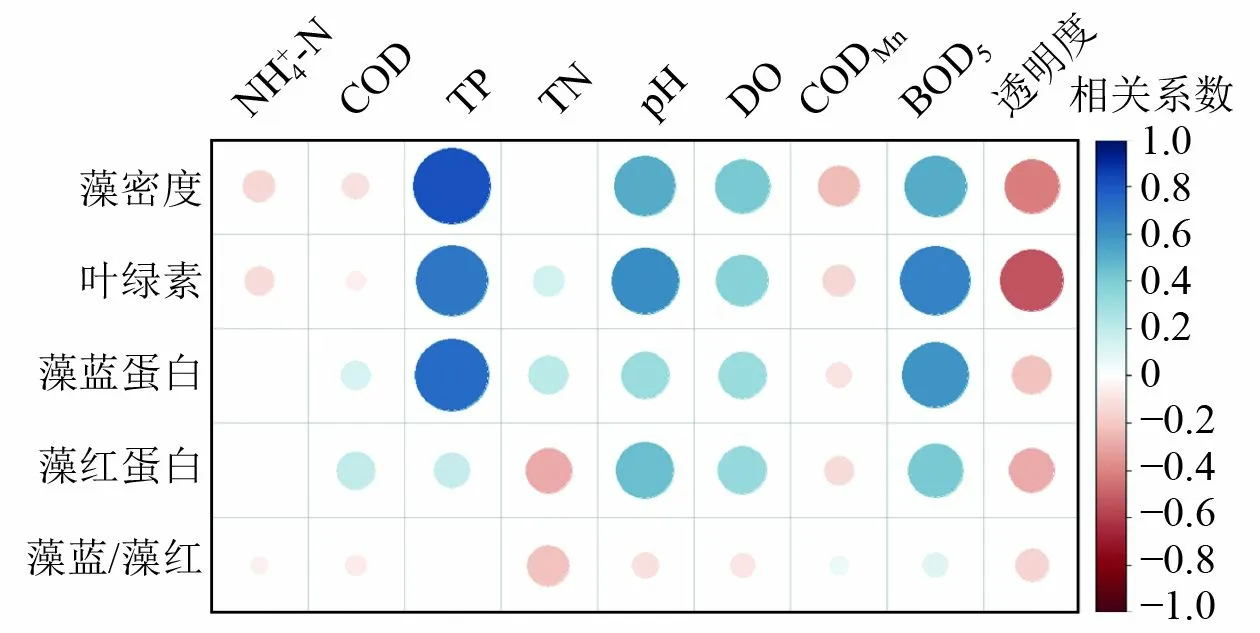
图5 滇池藻类组分与环境因子相关性分析Fig.5 Correlation analysis of algae components and environmental factors in Dianchi Lake
2.3.2 不同湖区藻胆蛋白的相关性
针对滇池藻蓝蛋白、藻红蛋白波动趋势的差异,分别分析滇池TN、TP 浓度的时空变化与藻胆蛋白时空分布关系。由2022 年滇池水体TN、TP 浓度和N/P 月度变化(图6)可知,草海的TN 浓度均高于3.5 mg/L,且TP 浓度在8 月达到0.10 mg/L 以上,TN、TP 浓度高于滇池其他湖区,且存在与其他湖区不一致的波动规律。这是由于草海上游区域的昆明主城区排放的污染负荷约占流域污染负荷总量的80%[11,29],对草海水体污染贡献显著,导致其氮、磷浓度高于外海且富营养化程度较高[30]。外海的TN、TP 浓度低于草海,TN 浓度在夏季最低,冬季较高;TP 浓度趋势相反,为春夏季较高,冬季较低[27]。外海TN 的主要来源为昆明主城区的污水和面源排放,夏季由于生态流量增加而稀释了TN 浓度[31]。TP 浓度在不同区域的变化规律差别较大,在外海北部是春季较高(5—6 月),外海中部的最高值出现在夏季(7—8 月),外海南部的最高值出现在秋季(9 月)。可见,滇池不同区域的水体N/P 差异显著,而每种藻类成为优势藻种都有其特定合适的营养盐浓度范围和所需的N/P,因此藻胆蛋白在滇池不同区域的浓度变化趋势也会有所差别[32-33]。
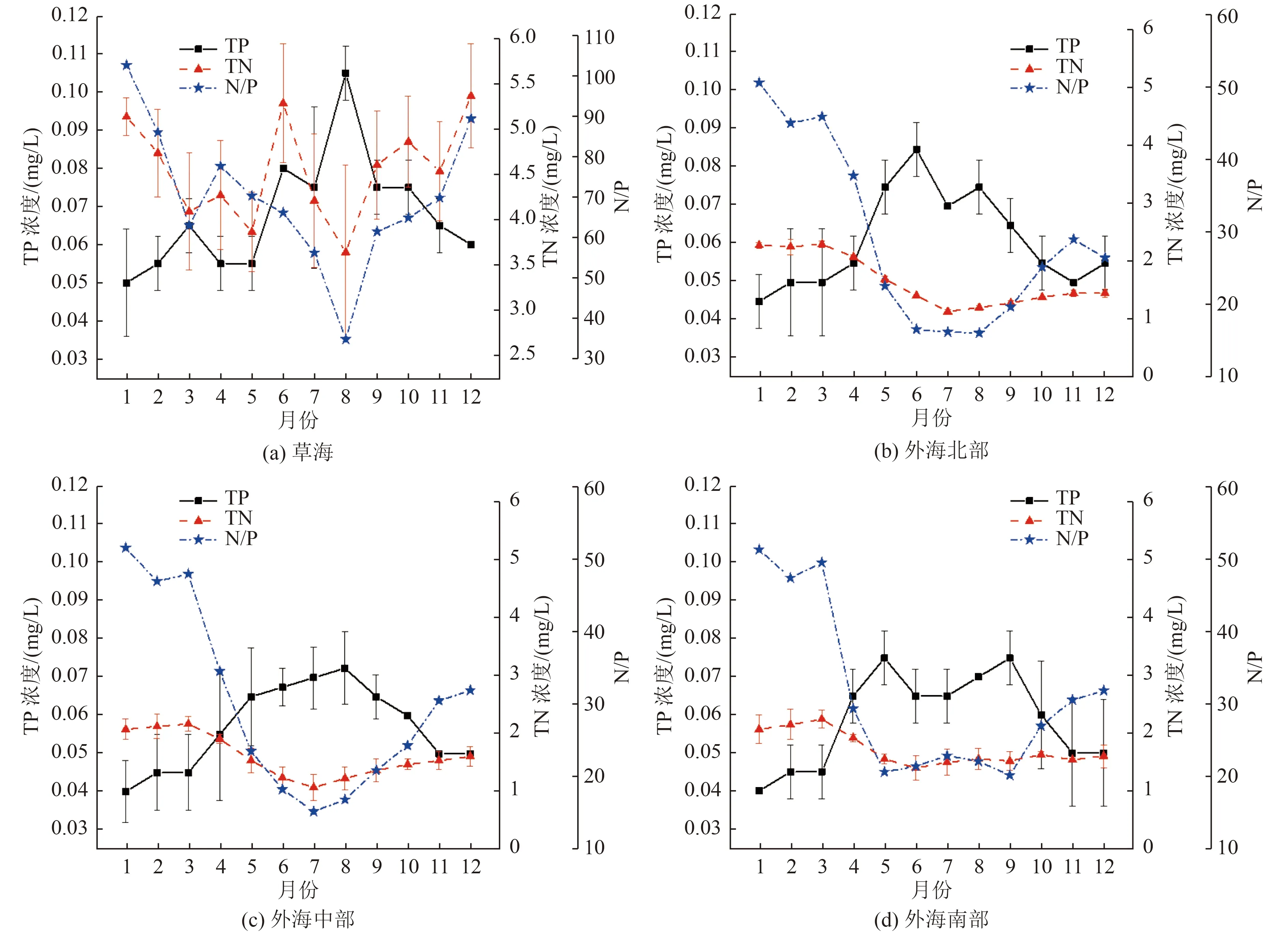
图6 2022 年滇池水体TN、TP 浓度和N/P 月度变化Fig.6 Monthly variation of TN, TP concentrations and N/P in Dianchi Lake in 2022
进一步分析滇池4 个湖区TN、TP 浓度与藻胆蛋白浓度的相关性,结果如表1 所示。由表1 可知,草海TN、TP 浓度之间不存在相关性,而外海中部和南部的TN、TP 浓度则表现为较显著的负线性关系。草海的藻红蛋白、藻蓝蛋白浓度与TN、TP浓度间的线性相关性较弱;外海北部和外海中部,藻蓝蛋白浓度与TP 浓度存在极显著的正线性相关关系(P<0.01),外海南部二者也存在显著的线性关系(P<0.05);外海北部和外海中部,藻蓝蛋白和藻红蛋白浓度与TP 浓度和N/P 均存在显著的负线性相关关系,尤其是藻蓝蛋白浓度与N/P 的线性相关性非常显著(P<0.01),外海南部藻胆蛋白浓度与N/P 的线性相关性降低。藻蓝蛋白与藻红蛋白的相关性方面,二者存在较为显著的线性相关性,说明产藻蓝蛋白的蓝藻种群和产藻红蛋白的红藻、硅藻种群具有正线性关系。除水体的营养盐因素外,藻密度的变化趋势还与气象、水文因素有关[34],下面结合已有文献对藻胆蛋白的控制因子进行讨论。

表1 滇池不同湖区水体TN、TP 浓度与藻胆蛋白浓度皮尔逊相关系数Table 1 Pearson correlation coefficient between TN, TP and phycobiliprotein concentrations in different lakes of Dianchi Lake
3 讨论
通过分析可以得出,藻胆蛋白主要集中在滇池湖体的中部区域,南北区域浓度均较低。滇池水华在草海和外海北部尤其是东北部较为严重,在外海南部较轻[35],这与藻胆蛋白的变化趋势有所不同。研究发现,藻胆蛋白的产生量与藻密度和种群类别相关[36],而滇池的水动力条件和营养盐浓度共同驱使了藻密度和藻种群的时空分布[37]。因此,藻胆蛋白的时空差异性可以从以下3 个方面解释:1)水动力条件。研究表明,水力停留时间越长,即流速越慢,越有利于水中藻类的繁殖[38]。滇池水力停留时间呈现草海<外海北部<外海南部<外海中部的规律,因此外海中部地区的水力条件最有利于藻类繁殖[39]。但调研显示,草海又是水华高发区,具有最高的藻密度,其原因可能是滇池的主导风向为西南风,使草海成为藻类漂移的聚集区域[40]。2013 年之后,牛栏江—滇池引水工程使外海北部15 条入湖河流水量占总入湖水量的85%以上,外海南部湖区的宽度变窄与出水河流的汇聚,导致南北水体的流动性强,东西水体的流动性差[41]。2)营养盐条件。滇池的藻类优势种为铜绿微囊藻(属于蓝藻门),当湖体的TN 和TP 浓 度 低 于2 和0.2 mg/L 的 阈 值 时[42],TN 和TP 浓度的增加可以促进藻类的繁殖,但当浓度超过该阈值时对其生长几乎无促进作用[43]。在研究阶段滇池大多数时间的TN 和TP 浓度均未超过阈值,因此营养盐是滇池水体水华暴发的重要限制因子。滇池不同区域的水体均呈现TP 浓度春夏季较高、秋冬季较低,TN 浓度春冬季较高、夏秋季较低的变化规律,N/P 呈现出冬季高、秋季低的趋势,且草海N/P 显著高于外海。相关研究表明,在高N/P 环境下,绿藻比蓝藻具有更强的利用优势,因此草海的藻类群落结构以绿藻为主,且春季由于N/P 较高,绿藻占比高,藻胆蛋白浓度低于秋季[44-45]。水体的盐类和金属离子含量在较高水平下,均会显著抑制藻胆蛋白,外海北部湖区承接了城区来水,污染成分复杂,这可能是造成藻胆蛋白产率低的又一重要原因[46-47]。3)藻种群。相关研究表明,滇池主要以蓝藻和绿藻为绝对优势种群,其中绿藻在南北部区域为绝对优势种,而蓝藻虽然相对红藻产量较低,但依然是藻红蛋白主要生产者[11-12]。从全湖均值来看,蓝藻在所有藻类中占绝对优势,绿藻仅次于蓝藻,其他藻类种群占比较小,对藻胆蛋白的贡献有限。已有研究表明,富营养化较高的水环境中,绿藻比蓝藻更具有生长优势[13],绿藻在春季的藻密度和种群占比较高,而蓝藻在春季的藻密度和种群占比较低,在冬季则较高。综上所述,水动力条件、营养盐浓度和藻种群密度三者共同影响了藻胆蛋白的产量分布,造成了其南北低、中部高,夏秋高、春冬低的时空分布特征。
对于藻胆蛋白的收集不能单纯根据藻密度来判断,例如草海虽然具有最高的藻密度,但其藻胆蛋白的产量却最低。藻蓝蛋白的产量远高于藻红蛋白,但其季节性差异却很强,主要集中在夏季和秋季;藻红蛋白的产量虽然较低,但其在夏秋冬三季都存在高产区。因此对于藻胆蛋白的收集和利用,需要根据实际情况安排打捞时机和打捞位置[48]。
4 结论
(1)滇池藻胆蛋白的浓度具有明显的季节变化,呈现夏季>秋季>春季>冬季的趋势。滇池藻胆蛋白的空间分布总体上呈现外海北部>外海中部>外海南部>草海的规律,且以藻蓝蛋白为主要成分,藻红蛋白的浓度较低。
(2)滇池的藻蓝蛋白浓度与TP 浓度和BOD5存在显著的正线性相关性(P<0.01),且外海藻蓝蛋白浓度与TP 浓度和BOD5的相关性强于草海,而藻红蛋白与TN、TP 浓度和BOD5的线性相关性均较低,说明BOD5和TP 浓度是控制藻蓝蛋白产生的重要因子,而藻红蛋白则受营养盐的影响较小。
(3)滇池藻胆蛋白的产生量受到水动力条件、营养盐浓度和藻种群密度三者共同影响,滇池外海北部和外海中部由于具有合适的N/P 和水力条件,因此在夏秋季表现出了较高的藻胆蛋白浓度。
