Lewis酸性MOF催化葡萄糖异构和差向异构反应机制
2023-11-21焦莹莹应制洲孙亮亮张元宝罗群兴
刘 苗,何 颖,焦莹莹,应制洲,杨 洋,孙亮亮,张元宝,罗群兴*
(1.西北大学化工学院,陕西 西安 710069; 2.陕西省合成气转化重点实验室,陕西师范大学化学化工学院,陕西 西安 710119; 3.厦门大学化学化工学院,福建 厦门 361005)
葡萄糖是合成生物质燃料和化学品的原料之一,也是人体细胞新陈代谢的主要能量来源,其可通过异构和差向异构分别转化为果糖和具有医药价值的甘露糖及其他稀有单糖[1-2],如阿洛酮糖、阿洛糖、塔格糖和艾杜糖等(如图1)。研究发现,果糖不仅是调节甜食中糖和热量水平的健康糖源,同时也是生物质转化为5-羟甲基糠醛(HMF)平台化合物的重要中间体[3];甘露糖能够抑制肿瘤细胞对葡萄糖的吸收,延缓癌细胞的生长和繁殖[4];阿洛酮糖和塔格糖热量低,能有效抑制人体血糖升高和体脂积累,是Ⅱ-型糖尿病和肥胖症人群的新型甜味剂[5]。因此,以丰富廉价的天然葡萄糖为原料,调控异构和差向异构反应路径,实现单糖分子间的定向催化转化,不仅有助于推动生物质资源的高值利用,而且对于改善人类膳食、医疗、保健和公共健康等重要民生问题具有重要意义,契合我国清洁能源和健康中国的发展战略需求。
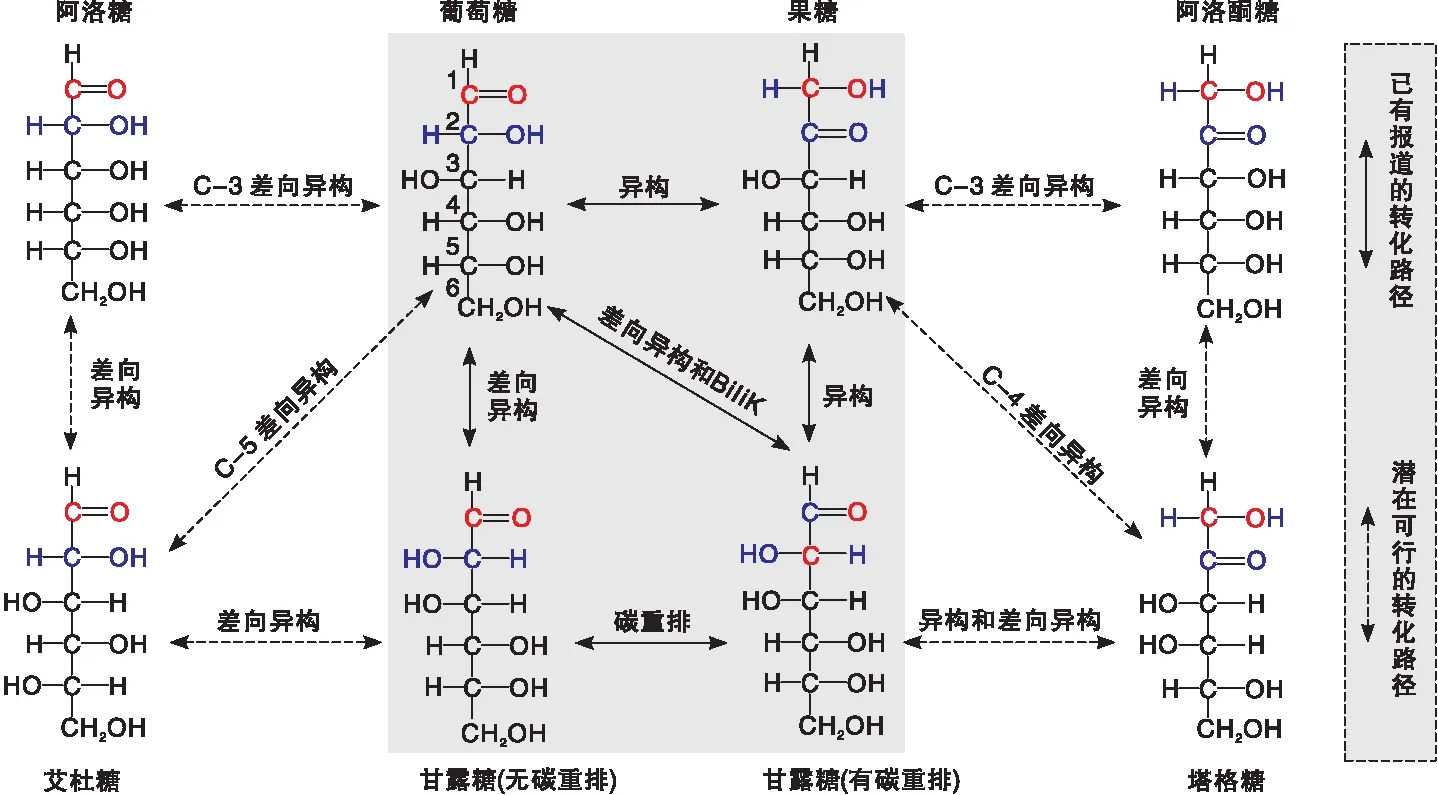
图1 单糖异构和差向异构的反应路径Figure 1 Schematic reaction paths of monosaccharide isomerization and epimerization
反应机理表明,碱性或Lewis酸性中心均可催化异构和差向异构反应,前者是通过烯醇中间物种(碱催化)或氢负离子转移(L酸催化)分别将醛糖转化为酮糖[6],后者是指葡萄糖C-2位置上羟基和氢原子在活性中心诱导作用下发生手性转换[4-7]。文献[8-9]报道,在纯硅Beta分子筛骨架中掺杂过渡金属Sn(Ⅳ)或Ti(Ⅳ)可构筑一种具有耐水性的Lewis酸性位点,该活性中心与醇羟基和羰基发生配位作用,通过氢负离子转移诱导羰基基团还原。在此基础上,Davis等将Sn-Beta用于水相催化葡萄糖的醛-酮异构反应中,首次提出位于疏水孔道内Lewis酸性位点优先与葡萄糖分子的O-1和O-2配位形成过渡态中间物种,再通过负氢离子转移生成果糖,且在短时间内达到热力学平衡[6,10-13]。此外,研究还发现,骨架内的Sn物种在水相中催化葡萄糖→果糖异构反应,在甲醇溶剂中却催化葡萄糖→甘露糖差向异构,而骨架外Sn物种在水相和甲醇中均只催化葡萄糖异构反应[12-13]。另一方面,Christianson J R等[14]添加硼酸盐作为助催化剂,也可实现Sn-Beta催化剂上异构向差向异构反应路径的转变。理论计算表明,硼酸盐与葡萄糖首先形成配合物,抑制异构反应发生,然后分子筛骨架上硅羟基(B酸位点)和硼酸盐-葡萄糖配合物协同催化C-1和C-2化学键断裂、C-1和C-3新键形成,通过碳骨架重排发生差向异构(Bìlik机理,图1阴影部分)[15]。
近年来,以金属有机骨架材料(MOF)为催化剂,利用其开放骨架中Lewis酸性金属位点和疏水骨架实现了葡萄糖异构和差向异构反应[16-20]。Luo Q X等[21]利用同位素示踪法和NMR光谱报道了Cr-MIL-101和Zr-UiO-66催化剂中Lewis酸性金属中心对葡萄糖异构和差向异构反应路径的差异,发现Cr(Ⅲ)在水或甲醇中只催化葡萄糖异构反应,但在高温时产生无骨架重排的差向异构产物;Zr(Ⅳ)中心可以同时催化葡萄糖异构和差向异构反应,且甘露糖产物发生分子内碳骨架重排。但是,关于不同金属Lewis酸性中心催化葡萄糖异构反应的机理和对异构反应路径影响机制的本质原因尚不明确。本文采用自旋极化的密度泛函理论,构建五配位Cr(Ⅲ)团簇和六配位Zr(Ⅳ)团簇为模型结构单元,研究两种催化剂本征活性位上葡萄糖→果糖异构和葡萄糖→甘露糖差向异构过程中的过渡态及能垒,阐明两种催化剂体系中葡萄糖转化反应路径的本质差异和机制。
1 理论计算
1.1 计算方法
采用自旋极化的密度泛函理论(SP-DFT)方法,包括广义梯度近似(GGA)、PBE泛函以及双精度极化数值基组DNP[22-25]。计算过程中轨道截断半径0.46 nm,结构优化中每个原子总能量和受力的收敛精度分别设为10-5Ha(1 Ha= 27.2114 eV)和0.1×10-3Ha·nm-1。其中,van der Waals弱相互作用通过DFT-D色散(Density Functional Theory Dispersion)方案予以矫正[26],而过渡态采用LST/QST或Dimer方法搜索[27]。此外,在计算过渡态过程中以第一步物理吸附后的稳定构型作为能量基点。
1.2 催化剂模型构建
金属有机骨架材料Cr-MIL-101由八面体构型金属Cr(Ⅲ)簇(六配位)和对苯二甲酸配位组装成的纳米笼状结构材料,通过加热脱出水分子或氟离子,每个金属Cr原子可以暴露出一个不饱和配位中心(CUSs);Zr-UiO-66材料是由八面体构型金属Zr(Ⅳ)簇(八配位)和对苯二甲酸配位组装成的三维多孔材料,通过脱除羧基可使每个金属Zr(Ⅳ)原子暴露出2个不饱和配位中心。为了简化计算,Zr-UiO-66和Cr-MIL-101结构分别采用最小的Zr与Cr的团簇模型来进行计算。其中,Zr采用六配位单金属中心,而Cr则采用五配位三金属团簇(如图2)。为降低远端不饱和氧对反应的干扰,这些氧原子采用H饱和形成OH端,并将这些远端OH固定,进而避免其对反应物中H剥离的影响。

图2 Cr-MIL-101(a)和Zr-UiO-66(b)的金属团簇结构单元模型Figure 2 Structural unit models of Cr-MIL-101 (a) and Zr-UiO-66 (b) metal clusters
2 结果与讨论
2.1 Zr-UiO-66模型体系理论计算

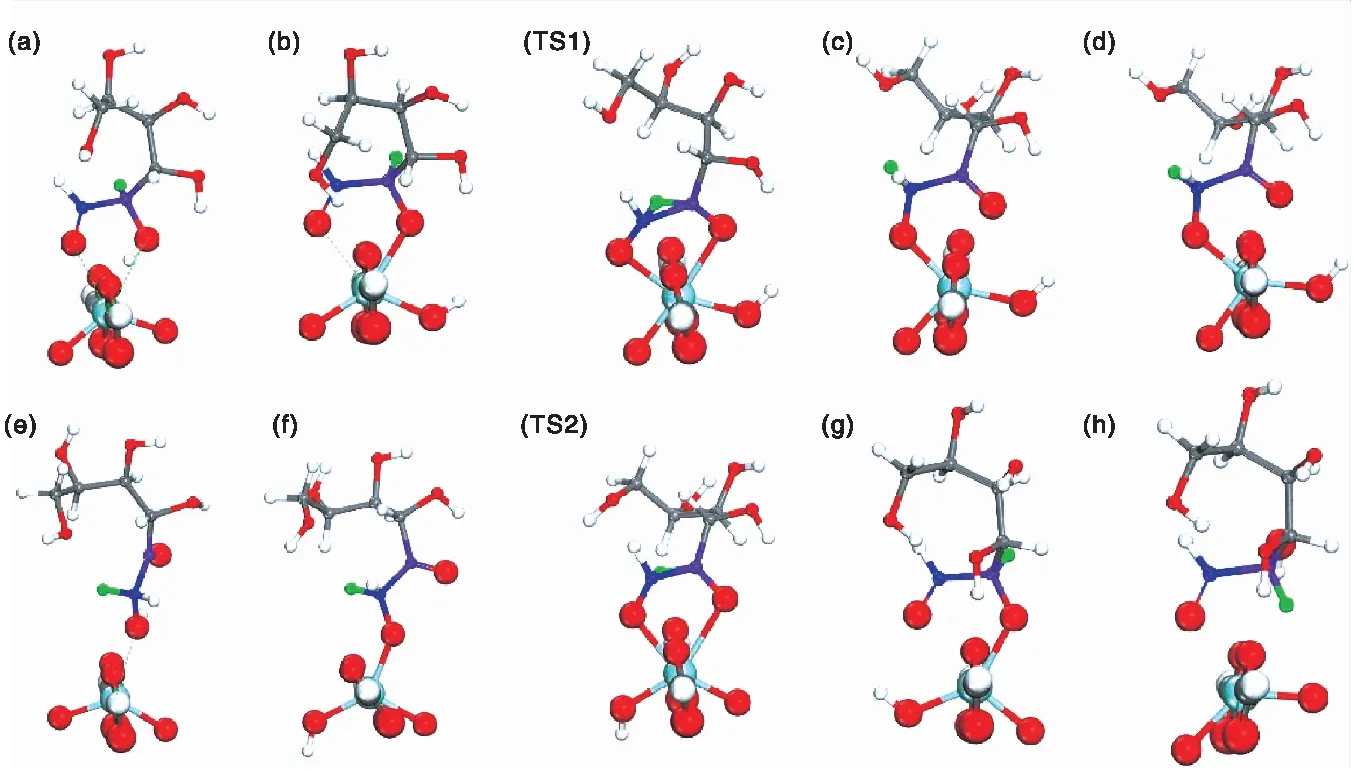
图3 Zr-UiO-66催化葡萄糖转化过程中涉及的中间体和过渡态Figure 3 Intermediates and transition states involved in glucose transformation over Zr-UiO-66C、H、O、Zr分别为灰色、白色、红色、青色;C1和C2原子分别为蓝色和紫色;负氢离子转移原子为绿色
图4为Zr-UiO-66催化葡萄糖转化基元反应步骤中间体及过渡态能垒。
从图4可以看出,葡萄糖C-1和C-2相连的羰基和羟基与金属Zr(Ⅳ)发生配位作用后,体系能量降低。负氢离子转移形成中间过渡态(TS1)过程中体系能量增加,该步骤(负氢离子转移)的活化能为62.12 kcal·mol-1,为整个反应过程的速控步骤。同理,当果糖分子与团簇再次发生物理吸附,体系能量降低,但随着金属配位活化和反负氢离子转移形成中间过渡态(TS2),体系能量再次升高,该步骤所需的活化能为47.69 kcal·mol-1。上述结果表明,果糖→甘露糖异构过程的反负氢离子转移活化能低于葡萄糖→果糖异构转化过程负氢离子转移活化能,导致Zr-UiO-66可以同时催化葡萄糖→果糖异构和葡萄糖→甘露糖差向异构,与前期实验结果一致[21]。
2.2 Cr-MIL-101模型体系理论计算

图5 Cr-MIL-101催化葡萄糖转化过程中涉及的中间体和过渡态Figure 5 Intermediates and transition states involved in glucose transformation over Cr-MIL-101C、H、O、Cr分别为灰色、白色、红色和浅蓝色,C1和C2分别为蓝色和紫色,负氢离子转移原子为绿色
图6为Cr-MIL-101催化葡萄糖转化基元反应步骤中间体及过渡态能垒。
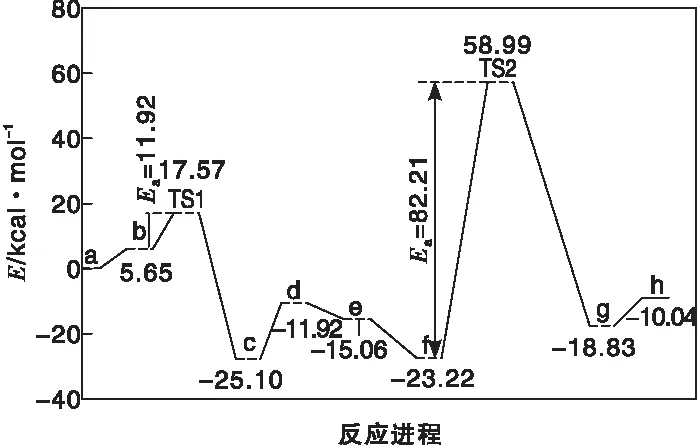
图6 Cr-MIL-101催化葡萄糖转化基元反应步骤中间体及过渡态能垒Figure 6 The energy profile of the intermediates and transition states involved in elementary steps of glucose transformation over Cr-MIL-101
3 结 论
(1) Zr-UiO-66和Cr-MIL-101催化剂对葡萄糖分子的配位活化机制不同。Zr-UiO-66模型结构单元中一个Zr(Ⅳ)原子分别与葡萄糖的醛基和羟基配位,Cr-MIL-101模型结构单元中两个Cr原子分别与葡萄糖的醛基和羟基配位。
(2) Zr-UiO-66催化体系中葡萄糖→果糖异构转化过程负氢离子转移活化能(62.12 kcal·mol-1)高于果糖→甘露糖异构过程的反负氢离子转移活化能(47.69 kcal·mol-1);Cr-MIL-101催化体系中葡萄糖→果糖异构过程的负氢离子转移活化能(11.92 kcal·mol-1)远低于果糖→甘露糖异构过程的反负氢离子转移活化能(82.21 kcal·mol-1)。
