低温驯化马占相思对橡胶树白粉病的抗性机制初探
2023-11-11黄曦泽冯德宇梁晓宇
王 燕,黄曦泽,冯德宇,王 萌,梁晓宇*,张 宇*
1. 海南大学植物保护学院,海南海口 570228;2. 海南大学三亚南繁研究院,海南三亚 572025
由白粉菌(Erysiphe quercicola)引起的白粉病是近年来影响橡胶树产胶量的主要因素之一,在环境条件适宜情况下极易暴发成灾,造成橡胶树反复落叶,严重推迟割胶时间。橡胶树白粉病暴发大多起始于每年的冬春交替之际,此时林段中橡胶树大多处于落叶期,可为白粉菌提供寄生的叶片较少。因此寻找橡胶树白粉菌的其他寄主植物,控制其初侵染的菌源,可在一定程度上降低白粉病的危害规模。目前已报道的橡胶树白粉菌寄主主要有马占相思(Acacia mangium)、飞扬草(Euphorbia hirtaL.)、麻风树(Jatropha curcasL.)和刺头婆(Urena lobatasp.)等[1]。其中马占相思是豆科(Leguminosae)含羞草亚科(Mimosoideae)金合欢属(Vachellia)的一种热带常绿高大乔木。作为重要的胶园防护林树种,马占相思有降低风速、贮水防旱、保护水土、降温防寒、改良土壤等生态效能,可为橡胶树的生长和产胶提供良好的条件,还可生产上等薪炭材料,创造丰厚的经济效益[2-4]。研究马占相思与橡胶树白粉菌的生物互作,可进一步了解白粉菌在寄主上的行为反应,为橡胶树白粉病的高效防治提供理论基础。
低温驯化是植物感受和传递低温信号引起低温适应性反应,提高抗冻性的过程[5]。低温驯化过程中植物体内发生许多生理与生化变化,如渗透调节物质大量积累、抗氧化酶活性增强、细胞膜脂质组成发生变化等[6]。前期研究发现,在低温环境中马占相思的新抽嫩叶不再感染橡胶树白粉菌,获得了显著的抗病性,推测是低温驯化提高了马占相思对橡胶树白粉病的抗性。为了探究其中的抗性诱导机制,本研究以23、19、15 ℃温度条件下培养的马占相思植株叶片为试验材料,验证这一抗性现象,利用扫描电镜和GC-MS/MS等仪器分析低温驯化对马占相思嫩叶蜡质的晶体结构和组成的影响,并测定接种橡胶树白粉菌的植株氧化酶活性,初步解析其抗病诱导机制。
1 材料与方法
1.1 材料
1.1.1 植物材料 供试品种为马占相思1年生幼苗,该品种购自广州市瑞景园林景观设计有限公司,在自然条件下继续培养待用;橡胶树品种为热研73397橡胶树芽接苗,购自中国热带农业科学院橡胶研究所,在自然条件下培养抽古铜叶片待接种。
1.1.2 菌株 橡胶树白粉菌OH-3菌株由本实验室分离纯化,接种至热研73397橡胶树芽接苗上扩繁培养。
1.2 方法
1.2.1 低温驯化 马占相思的原产地属于热带湿润气候,年均温度不低于23 ℃[7]。故本研究以23 ℃作为马占相思的自然生长温度,19 ℃和15 ℃作为马占相思的低温驯化温度,将长势一致且健康的马占相思幼苗分别转移至上述3个温度的人工气候箱(光照时间12 h/d,光照强度10 000 lx,空气相对湿度85%)中培养30 d。
1.2.2 橡胶树白粉菌接种 用软毛笔刷将橡胶树芽接苗叶片上白粉菌的孢子粉轻轻扫到烧杯中,加入无菌水涡旋振荡摇匀,用血球计数板计数,制备浓度为105个/mL的孢子悬浮液。用喷壶将孢子悬浮液均匀地喷雾至马占相思幼苗叶片表面,后将幼苗置于培养条件为19 ℃、光照时间12 h/d,光照强度10 000 lx,湿度85%~90%的人工气候室中培养并观察病程。每个驯化温度下接种6株苗,实验重复3次。
1.2.3 扫描电镜观察 将马占相思叶片剪成1 cm2大小的片段,用2.5%戊二醛浸泡后,4 ℃下固定12 h;0.1 mol/L磷酸缓冲液冲洗3次,每次20 min;梯度酒精脱水(30%、50%、70%、90%、100%)各20 min,醋酸异戊酯置换酒精清洗1次;冷冻干燥12 h。将样品粘于导电胶并固定在托盘上,抽真空后喷金,利用扫描电镜(Phenom G6 pure)对叶片表面进行蜡质晶体形态的观察[8-9]。每个驯化温度下处理3片叶,实验重复3次。
1.2.4 蜡质提取 蜡质提取参照赵帅等[10]的方法稍加改进。将马占相思叶片至于装有20 mL三氯甲烷的烧杯中,轻轻摇晃烧杯加速蜡质溶解,后将液体转移至旋蒸瓶中,待三氯甲烷在40 ℃下完全蒸干,分别加入30 μL吡啶和30 μL双(三甲基硅烷基)三氟乙酰胺,密封后放入70 ℃水浴锅中衍生反应1 h,反应完全后再次40 ℃旋蒸干,分次加入1 mL三氯甲烷溶解,转移至进样瓶。每个驯化温度下处理9片叶,实验重复2次。
1.2.5 GC-MS/MS分析 使用Trace1300-TSQ 9000气相质谱联用仪(美国赛默飞)对上述蜡质提取液进行成分的定性定量分析。GC-MS/MS条件:载气为氦气,进样量1 μL,进样口温度280 ℃,检测器温度320 ℃;检测程序:50 ℃保持2 min,以20 ℃/min升温至240 ℃保持2 min,再以1.5 ℃/min升温至320 ℃保持15 min。参考计算机质谱数据库,依据蜡质成分中酸、醇、醛和烷烃均有其明显的M-15特殊离子峰判定各蜡质组分种类以及碳原子数,检索后判定化合物名称[11]。蜡质中各组分相对含量根据其离子峰面积占总离子峰面积的比值进行计算。
1.2.6 氧化酶活性测定 称取0.2 g马占相思叶片,在预冷研钵中研磨成粉后加入4 mL磷酸缓冲液(0.05 mol/L, pH 7.8),4 ℃条件下10 000 r/min离心20 min,转移上清液至新离心管中用于抗氧化酶活性测定。超氧化物歧化酶(SOD)活性采用氮蓝四唑法NBT法测定[12];过氧化氢酶(CAT)活性采用过氧化氢法测定;过氧化物酶(POD)活性采用愈创木酚法测定[13]。每个驯化温度下处理6片叶子,实验重复3次。酶活性单位均以单位蛋白含量的酶活性表示,U/g。
1.3 数据处理
采用Excel软件进行数据整理,采用DPS和Origin软件进行差异显著性分析和制图。
2 结果与分析
2.1 低温驯化增强马占相思对橡胶树白粉病的抗性
将马占相思幼苗分别置于23、19、15 ℃温度条件下培养抽出嫩叶,后转移至橡胶树白粉菌最适温度19 ℃条件下接种(图1)。接种12 d后发现,23 ℃培养的植株嫩叶发病严重,上下表面布满含有大量菌丝的病斑;23 ℃培养的植株成熟叶均不发病;19 ℃培养的嫩叶发病较轻,上下表面有少量且菌丝稀少的病斑,19 ℃培养的成熟叶均不发病;15 ℃培养的嫩叶和成熟叶均不发病,将低温驯化后不发病植株重新置于23 ℃下培养至抽出嫩叶,新抽叶片接种白粉菌后发病严重。由此可见,低温驯化显著增强了马占相思对橡胶树白粉病的抗性。

图1 不同培养温度下马占相思叶片接种橡胶树白粉菌的病症Fig. 1 Lesion of A. mangium leaves inoculated with E. quercicola under different culture temperature conditions
2.2 低温驯化对马占相思嫩叶蜡质晶体结构的影响
通过观察不同温度条件培养的马占相思叶片表面形态(图2),发现23 ℃培养的嫩叶表面不平整,有明显的凹凸,19 ℃和15 ℃培养的嫩叶表面趋于平滑,有轻微的凹凸,叶片形态更趋近于23 ℃培养的成熟叶。利用扫描电镜观察了马占相思叶片表面的蜡质层结构,发现23 ℃嫩叶蜡质层凸起、排列不规则,晶体结构相互叠加缠绕;19 ℃嫩叶蜡质层凸起,结构排列有序;23 ℃成熟叶和15 ℃嫩叶蜡质层平滑,有明显的晶体状蜡质。综上所述,低温驯化可促进马占相思嫩叶蜡质晶体结构的成熟化。

图2 不同培养温度条件下马占相思叶片的上表面形态Fig. 2 Morphology of the upper surface of A. mangium leaves under different culture temperature conditions
2.3 低温驯化对马占相思嫩叶蜡质组分的影响
提取不同温度条件下培养的马占相思叶片蜡质,利用GC-MS/MS检测体系对蜡质组分进行定性定量分析,不同叶片的蜡质检测总离子峰图如图3所示。与23 ℃嫩叶的蜡质检测离子峰相比,15 ℃嫩叶、19 ℃嫩叶和23 ℃成熟叶的检测离子峰更多,峰图更趋近一致,说明15 ℃嫩叶、19 ℃嫩叶和23 ℃成熟叶的蜡质组成更复杂。进一步筛选到10个响应强度高的离子峰,经检索质谱数据库,推测出这些化合物的种类。10个蜡质成分主要归为5类,其中烷烃类化合物有5种,分别为C25烷、C27烷、C31烷、C36烷、C44烷;酚类化合物有2种,分别为C14酚和C23酚;酮类、醇类和酯类化合物各1种,分别为C30酮、C30醇和C32酯。
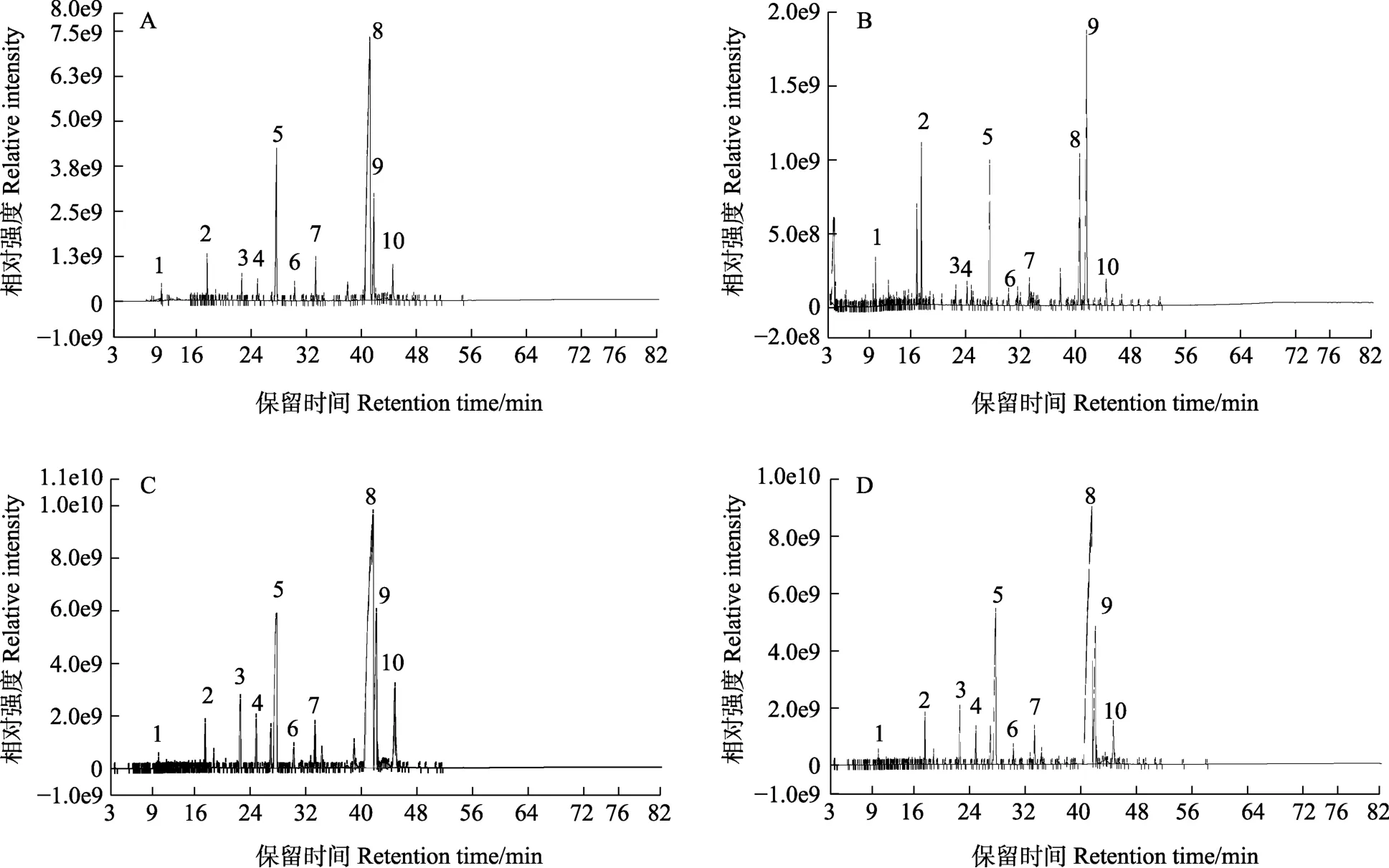
图3 不同培养温度条件下马占相思叶片的蜡质成分检测总离子峰Fig. 3 The total ion peak of A. mangium leaf wax under different culture temperature conditions
以检测总离子峰为100%分析比较以上化合物在不同温度条件下培养的马占相思叶片蜡质中的相对含量(表1)。结果表明,马占相思叶片蜡质中C30酮的相对含量最高,不同培养温度的叶片C30酮相对含量差异显著,由低到高排列依次为:15 ℃嫩叶<23 ℃成熟叶<19 ℃嫩叶<23 ℃嫩叶。另外,在23 ℃嫩叶中相对含量较高的有C36烷、C32酯和C30醇,而15 ℃嫩叶和23 ℃成熟叶中的相对含量均显著降低。由此可见,不同温度对马占相思叶片蜡质成分的影响较大,低温驯化使叶片蜡质组成更复杂,一定程度上使嫩叶的主要蜡质成分趋近于成熟叶。

表1 不同培养温度条件下马占相思叶片蜡质主要成分的相对含量Tab. 1 Relative contents of the main components of wax in A. mangium leaves under different culture temperature conditions
2.4 低温驯化对马占相思嫩叶氧化酶活性的影响
通过比较23 ℃和15 ℃培养的马占相思嫩叶接种橡胶树白粉菌后SOD、CAT和POD氧化酶活性发现。随着侵染时间延长,2种温度培养的嫩叶的3种氧化酶活性均呈先升高后降低的趋势,且在24 h时达到最高值。与23 ℃嫩叶相比,15 ℃嫩叶的SOD、CAT和POD酶活性在部分侵染时间内显著增强,其中SOD酶表现最为明显(图4)。综上表明,低温驯化可以在一定程度上提高马占相思嫩叶在接种白粉菌时的氧化酶活性。
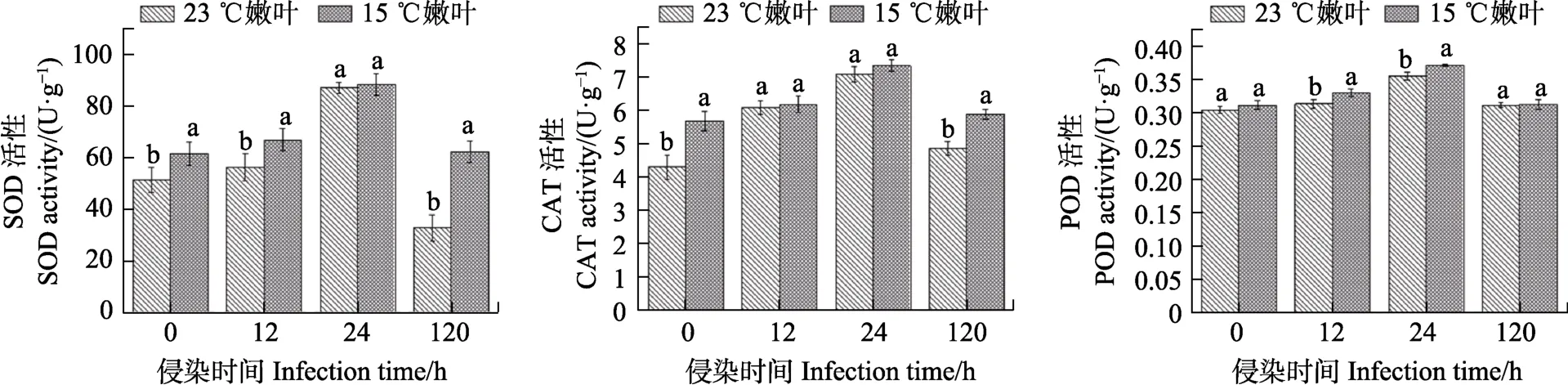
图4 23 ℃和15 ℃培养的马占相思嫩叶的氧化酶活性Fig. 4 Oxidase activity of young leaves of A. mangium incubated at 23 ℃ and 15 ℃
3 讨论
温度是影响植物生理过程的重要非生物因素[14]。本研究利用不同温度驯化的马占相思幼苗嫩叶接种橡胶树白粉菌后发现,15 ℃温度条件下抽出的嫩叶不发病,说明低温驯化可显著增强马占相思嫩叶对橡胶树白粉病的抗性。已有研究表明,植物可以通过改变根、茎、叶的形态,解剖结构和超微结构来适应低温环境[15]。植物还可以通过积累细胞内渗透调节物质[16],增加代谢相关蛋白、分子伴侣、抗冻蛋白的合成来耐受或抵抗低温胁迫[17]。推测驯化过程中的低温胁迫引起马占相思的嫩叶形态结构或组成物质的变化,从而产生了对橡胶树白粉病的抗性。
本研究观察了不同驯化温度条件下马占相思叶片的表面形态,发现19 ℃和15 ℃培养的嫩叶表面相对平滑,形态更接近23 ℃培养的成熟叶。扫描电镜结果表明15 ℃嫩叶和23 ℃成熟叶蜡质层结构趋近一致,表面平滑且有明显的晶体状蜡质,这进一步说明低温驯化可促进马占相思嫩叶蜡质晶体结构的成熟化。植物表皮蜡质是植物抵御外界生物胁迫的第一屏障,通过提高表皮的机械强度和疏水性,能有效防止真菌等病原体的侵害[18-19]。蜡质晶体结构容易受温度诱导产生重组现象,当植物处于低温环境时,蜡质晶体结构容易形成棒状、管状等结构,而处于高温环境时容易形成板状、片状结构[20]。本研究中,马占相思的嫩叶易受橡胶树白粉菌侵染,而成熟叶片对橡胶树白粉病具有天然抗性,因此,推测低温驯化可能是通过促进马占相思嫩叶蜡质晶体结构的成熟化,形成了更具防御性的物理屏障,通过阻碍白粉菌的侵染发挥抗病作用。
环境温度不仅能够影响植物蜡质的晶体结构,还会影响烷烃、烯烃、醇、酮、酯和酚等蜡质组分的分布和比例[21]。本研究利用GC-MS/MS检测技术对不同温度驯化的马占相思叶片蜡质组分进行了定性定量分析,发现其主要含有烷烃、醇、酮、酯、酚等5类有机化合物,C30酮相对含量最高。总离子峰图和相对含量结果进一步表明,低温培养使叶片蜡质组成更复杂,15 ℃嫩叶和23 ℃成熟叶的蜡质组成较一致,说明低温驯化在一定程度促进嫩叶的主要蜡质成分比例趋近于成熟叶。已有研究表明,一些植物蜡质组分可以抑制或者刺激真菌孢子的萌发。水稻叶片蜡质组分十六烷二醇可以诱导稻瘟菌附着胞的分化形成,并促进灰霉菌孢子萌发和角质酶的表达[22]。因此,推测低温驯化也可能促进或抑制了马占相思嫩叶的某种蜡质成分,从而调控病原菌的侵染和增殖分化来实现蜡质的化学防御功能。
另外,一些植物蜡质成分被真菌角质酶降解的产物,也可作为植物损伤相关分子模式的诱导子被植物模式识别受体所识别,通过诱导植物活性氧的积累,继而启动下游包括超敏反应在内的一系列防御反应[23-24]。如,外施无抑菌活性的蜡质单体可增加大麦对白粉菌的抗性[25]。本研究测定了23 ℃和15 ℃培养的马占相思嫩叶接种橡胶树白粉菌后的SOD、CAT和POD氧化酶活性,发现低温驯化可以在一定程度上提高马占相思嫩叶在接种白粉菌时的氧化酶活性。因此,推测低温驯化也可能促进马占相思嫩叶生成可作为诱导子的蜡质组分,当被白粉菌侵染时激发了下游的免疫反应,从而产生抗病性。当然本研究还有许多值得探讨的问题,如,马占相思叶片中哪种蜡质成分发挥了对橡胶树白粉病的抗病作用?低温驯化是否还会通过影响蜡质之外的物质来提高马占相思的抗病性?抗病相关机制需要代谢组和转录组等高通量分析手段和分子生物学实验进行进一步验证。
4 结论
本研究对不同温度培养的马占相思接种橡胶树白粉菌,发现低温驯化显著增强了马占相思对橡胶树白粉菌的抗性;通过比较不同驯化温度的嫩叶蜡质晶体结构和组分差异,发现低温驯化可促进马占相思嫩叶蜡质晶体结构和主要成分的成熟化;进一步比较不同温度培养的叶片氧化酶活性发现,低温驯化可以一定程度提高马占相思接种橡胶树白粉菌后的氧化酶活性。本研究初步探索了橡胶树白粉病寄主的抗病机制,为橡胶树的抗病育种提供了理论依据。
