我国不同地区草地贪夜蛾内共生菌感染和耐寒能力差异研究
2023-11-09牛洪涛张志春赵冬晓郭慧芳
胡 慧,王 娜,牛洪涛,张志春,赵冬晓,郭慧芳
(江苏省农业科学院植物保护研究所,南京 210014)
草地贪夜蛾Spodopterafrugiperda是一种新入侵我国的重大害虫,自2019年1月首次在云南发现以来,迅速扩散至全国大部分地区,给玉米安全生产造成严重威胁。迁飞能力强、繁殖力高是草地贪夜蛾快速扩散的重要原因(郭井菲等,2019;齐国君等,2020)。
昆虫中普遍存在内共生菌,它们在调节宿主生殖和抗逆性等方面发挥着重要作用,是其进化的重要动力(Himleretal., 2011; van den Bosch and Welte, 2016; Lietal., 2020)。蚜虫、烟粉虱Bemisiatabaci等重要刺吸类害虫中的内共生菌已受到广泛关注,除原生共生菌外,发现它们还含有Regiella、Hamiltonella、Arsenophonus、Rickettsia、Serratia和Wolbachia等多种次生共生菌,共生菌的功能各不相同,主要包括提高宿主生殖能力以及抗杀虫微生物和寄生蜂能力等抗逆功能(Oliveretal., 2005; Himleretal., 2011;ukasiketal., 2013; Liu and Guo, 2019; Lietal., 2020)。此外,还发现共生菌也可削弱宿主的抗逆能力,如非洲粘虫中Wolbachia能降低宿主对昆虫核型多角体病毒的抵抗力(Grahametal., 2012),褐飞虱Nilaparvatalugens中共生菌Arsenophonus则降低了宿主对化学药剂的抵抗力(Pangetal., 2018)。较刺吸类害虫而言,鳞翅目害虫体内共生菌研究相对较少。在非洲粘虫和幻紫斑蛱蝶Hypolimnasbolina等中,发现了Wolbachia的感染(Dysonetal., 2002;Grahametal., 2012)。关于草地贪夜蛾体内微生物,仅有关于肠道微生物的研究,从幼虫中筛选分离出14种肠道细菌菌株,包括2种纤维素降解细菌(李宏伟等,2020)。该害虫中内共生菌感染情况如何?国内外尚未见报道。
昆虫的生长发育和繁殖有着适宜的温度,温度过高或者过低都会影响其正常的生命周期,严重时导致昆虫死亡(Broufasetal.,2009)。昆虫在长期进化过程中为适应恶劣的生境条件,表现耐寒和耐热能力多样性,如二化螟Chilosuppressalis不同地理种群耐寒能力有差异,在相同的低温下,北京种群的半致死时间较福州种群更长(张珺等,2005)。在15℃~30℃范围内,草地贪夜蛾在15℃下的死亡率最高,不能完成整个世代发育(刘小宇等,2022)。不同地区草地贪夜蛾耐寒能力是否存在差异?其是否与害虫休内生菌感染存在相关性,均未见研究报道。本文将比较不同地区采集草地贪夜蛾共生菌感染的差异以及耐寒能力差异,以为进一步分析共生菌功能奠定基础,也为我国草地贪夜蛾迁飞路线分析提供依据。
1 材料与方法
1.1 供试昆虫
2019年5-7月分别在我国不同地区玉米田采集草地贪夜蛾幼虫,每个地区采集35~128头(采集具体信息见表1)。草地贪夜蛾采回后在室内用人工饲料饲养,按地区分别进行群体饲养,饲养条件:温度27±1℃,相对湿度70%±5%,光周期L∶D=14∶10。饲养1~3代后,在各种群中随机选用3龄以上幼虫进行共生菌检测,待检测幼虫单头分装到离心管中,保存于-80℃冰箱。
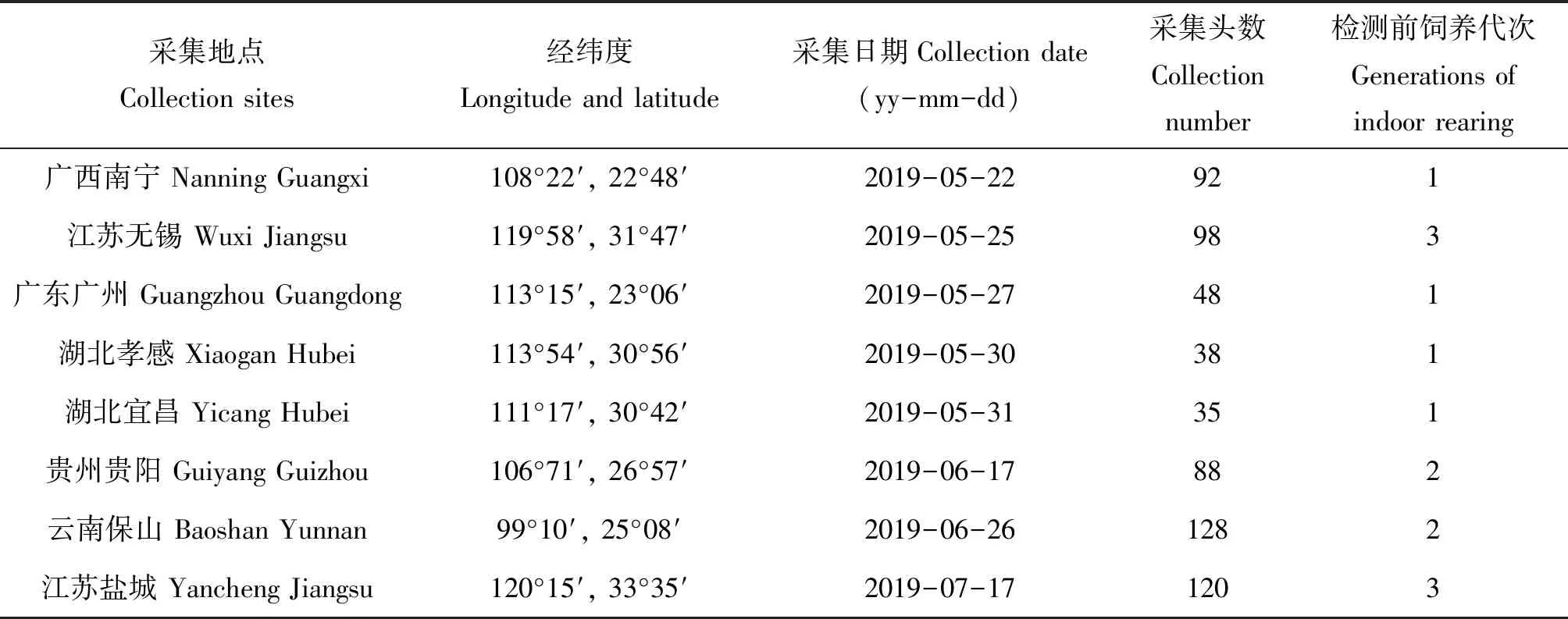
表1 草地贪夜蛾的采集信息
1.2 草地贪夜蛾基因组DNA提取
草地贪夜蛾单头幼虫放置于1.5 mL离心管中,先用75%酒精清洗一次,再用无菌水清洗一次,液氮中速冻后研磨至粉状后,立即加入350 μL PBS温和研磨30 s,用DNA提取试剂盒(Promega A1125)按说明书进行DNA提取,-20℃冰箱中保存备用。
1.3 草地贪夜蛾夜蛾体内共生菌检测
除广西南宁种群收集84头试虫外,其余各种群均收集60头试虫用于检测,共检测Portiera、Hamiltonella、Wolbachia、Cardinium、Arsenophonus、Rickettsia、Fritschea、Serratia和Acinetobacter9种共生菌,引物详细信息见表2(上海生工生物工程公司合成);PCR扩增反应体系为25 μL,包括 2×Master Mix 13 μL,上游引物和下游引物各1 μL,灭菌超纯水8 μL,DNA模板2 μL。扩增程序因共生菌不同而异,共9个程序,分别为:

表2 草地贪夜蛾不同地理种群共生菌感染率

表2 共生菌检测用引物序列
(A)95℃ 5 min;94℃ 30 s,56℃ 1 min ,72℃ 1 min,循环35次;72℃ 20 min。
(B) 95℃ 3 min;94℃ 30 s,58℃ 30 s,72℃ 45 s,循环35次;72℃ 10 min。
(C) 94℃ 2 min;94℃ 1 min,65℃→55℃,1 min,72℃ 2 min,循环10次;94℃ 1 min,55℃ 1 min,72℃ 2 min,循环25次;72℃ 6 min 。
(D) 95℃ 3 min;94℃ 1 min,58℃ 1 min,72℃ 90 s,循环35次;72℃ 5 min。
(E) 95℃ 5 min;95℃ 1 min,58℃ 1 min,72℃ 1 min,循环30次;72℃ 20 min。
(F) 95℃ 5 min;94℃ 1 min,64℃ 1 min,72℃ 90 s,循环35次;72℃ 5 min。
(G) 95℃ 3 min;94℃ 1 min,56℃ 1 min,72℃ 1 min,循环5次;95℃ 1 min,58℃ 1 min,72℃ 1 min,循环30次;72℃ 20 min。
(H) 95℃ 3 min;94℃ 1 min,56℃ 1 min,72℃ 90 s,循环35次;72℃ 10 min。
(I)95℃ 3 min;94℃ 1 min,56℃ 1 min,72℃ 90 s,循环35次;72℃ 10 min。
PCR产物的纯化、连接转化与序列测定:使用普通琼脂糖凝胶DNA回收试剂盒回收PCR产物(TIANgel Midi Purification Kit),具体步骤参照说明书。连接转化反应按照pEASY-T3 Cloning Kit试剂盒说明书进行。挑斑、摇菌后,将阳性克隆用于测序,所有测序均由南京金斯瑞生物科技有限公司完成,测序结果在NCBI网站中进行BLAST比对。
1.4 草地贪夜蛾夜蛾耐寒能力测定
通过测定-6℃低温下草地贪夜蛾的死亡率来比较害虫的耐寒能力。将经过室内连续20代人工饲养的各地区草地贪夜蛾2龄幼虫放入装有人工饲料的培养皿中(直径35 mm),每个培养皿20头作为一个重复,每个种群5个重复,放置于人工气候培养箱中。培养箱温度从27℃以0.25℃/min速度降至0℃,在0℃保持3 h后,再以0.25℃/min的速度降至-6℃,在-6℃保持5 h后,以0.25℃/min的速度升温至27℃,在27℃下保持48 h。每隔24 h加入新鲜人工饲料,观察记录各培养皿中死亡虫数,计算死亡率。
1.5 数据处理与分析
应用MEGA 6软件对草地贪夜蛾幼虫内共生菌进行同源性分析,邻接法(Neighbor-Joining(NJ))构建基于16S rDNA序列或23S rDNA序列的系统发育树。应用SPSS 19.0软件对不同地区草地贪夜蛾共生菌感染率差异进行χ2检验分析,对低温下害虫死亡率采用单因素方差分析和Tukey多重比较法进行不同地区间的差异显著性分析,对共生菌感染率与低温死亡率进行相关性分析。
2 结果与分析
2.1 我国草地贪夜蛾携带共生菌种类
对来自广西、广东、湖北、云南、贵州和江苏等8个地区草地贪夜蛾进行共生菌检测,共检测包括Portiera、Cardinium、Arsenophonus、Hamiltonella、Wolbachia、Rickettsia、Fritschea、Acinetobacter、Serratia等9种共生菌。发现各地草地贪夜蛾中只检测到Cardinium和Arsenophonus这两种共生菌,Cardinium16S rDNA序列号为MN864662.1,Arsenophonus23S rDNA序列号为MW282126.1。
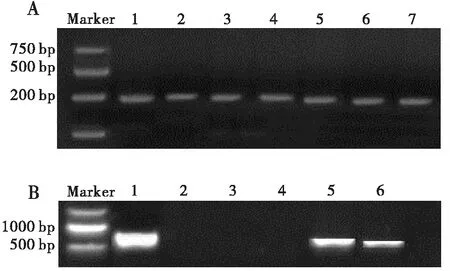
图1 草地贪夜蛾体内共生细菌扩增产物电泳图
2.2 草地贪夜蛾体内Arsenophonus和Cardinium的同源性分析
比较分析了草地贪夜蛾与其它昆虫体内共生菌Arsenophonus的同源性,基于23S rDNA序列通过NJ法构建的系统发育树显示(图2),草地贪夜蛾体内Arsenophonus与同属鳞翅目的二化螟中Arsenophonus亲缘关系最近,在同一分支上,与烟粉虱、丽蝇蛹集金小蜂Nasoniavitripennis、大豆蚜Aphisglycines中Arsenophonus的亲缘关系相对较远,在另一分支上,而与白蜡蚧Ericeruspela和褐飞虱的亲缘关系最远,后两者在另一独立分支上。
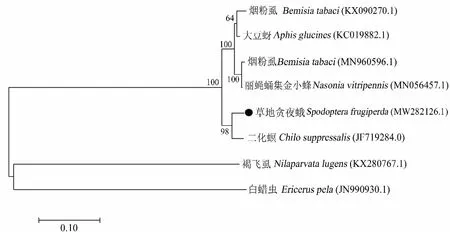
图2 基于23S rDNA序列构建的草地贪夜蛾共生菌Arsenophonus系统发育树
对草地贪夜蛾与其它昆虫体内共生菌Cardinium的同源性进行了分析,基于16S rDNA序列分析构建系统发育树表明(图3),草地贪夜蛾体内共生菌Cardinium与果蝇Drosophilabifurca、螫斑鼓翅蝇Sepsispunctum、恩蚜小蜂Encarsiainaron最近,在同一分支上;与烟粉虱体内Cardinium较为接近,而与Hamaliasirokata和白背飞虱Sogatellafurcifera中Cardinium亲缘关系相对较远,与螨Microzetorchestesemeryi中Cardinium的亲缘关系最远。

图3 基于16S rDNA 序列构建的草地贪夜蛾共生菌Cardinium系统发育树
2.3 不同地区草地贪夜蛾共生菌感染率差异
对采自江苏、广东、广西、贵州、云南和湖北等6个省份8个地区草地贪夜蛾共生菌感染率进行了比较分析,结果发现,Arsenophonus和Cardinium这两种共生菌感染率在不同地区间均存在显著差异(表2)。从Arsenophonus的感染来看,各地理种群中总体较低,以云南保山地区草地贪夜蛾Arsenophonus的感染率最高,为20.0%,其次是广西南宁地区,感染率为15.5%,贵州贵阳和江苏盐城的感染率均为1.7%,而广东广州、江苏无锡以及湖北宜昌和孝感这4个地理种群中均未发现Arsenophonus的感染(χ2=78.658, df=7,P<0.001)。共生菌Cardinium的感染总体较高,感染率最高的为江苏无锡种群,高达96.7%,其次为云南保山种群,感染率为93.3%,广东广州种群感染率为86.7%,贵州贵阳种群为53.3%,湖北孝感和宜昌种群感染率则分别为30.0%和18.3%,江苏盐城种群感染率也为18.3%,而广西南宁种群中未发现有Cardinium感染(χ2=296.727, df=7,P<0.001)。
2.4 不同地区草地贪夜蛾耐寒能力比较
为明确不同地区草地贪夜蛾耐寒能力差异,比较了广西南宁等8个地区草地贪夜蛾在-6℃低温胁迫下的死亡率。结果发现,不同地区草地贪夜蛾耐寒能力存在显著性差异,湖北宜昌地区草地贪夜蛾耐寒能力最强,低温胁迫后死亡率最低,为35%,显著低于江苏盐城和湖北孝感地区死亡率(76%和69%),其它6个地区害虫低温下死亡率也较低,在46%~49%之间,同样显著低于盐城和孝感地区,但6个地区间无显著差异(F7, 39=4.653,P=0.0011;图4)。
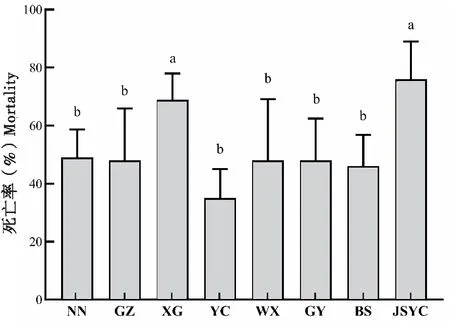
图4 不同地区草地贪夜蛾-6℃胁迫下的死亡率比较
2.5 草地贪夜蛾共生菌感染与耐寒能力相关性分析
对各地区草地贪夜蛾共生菌感染率和低温下害虫死亡率相关性分析表明,Cardinium感染率与-6℃低温胁迫下的死亡率没有显著相关性(线性回归相关系数=0.3971,P>0.05),Arsenophonus感染率与-6℃低温胁迫下的死亡率也没有显著相关性(线性回归相关系数=0.5023,P>0.05),说明草地贪夜蛾中Cardinium和Arsenophonus感染与宿主耐寒能力均无直接相关性。
3 结论与讨论
本研究首次发现了草地贪夜蛾体内有内共生细菌Cardinium和Arsenophonus的感染,两种共生菌的感染在我国2019年采集的不同地理种群中存在差异。Cardinium在我国江苏、云南以及广东各地草地贪夜蛾中感染率较高,而在广西南宁种群中未被发现,而Arsenophonus在被检测种群中有一半未被发现,已发现地区感染率也不高。作为一种新入侵我国的害虫,草地贪夜蛾一入侵后就表现出了共生菌感染在不同地区间的差异,说明草地贪夜蛾在入侵我国的初期尚未受到本地其它害虫共生菌的显著影响,可能与迁入虫源相关。
关于草地贪夜蛾在我国的迁飞路径,Li等(2020)利用轨迹分析方法预测了草地贪夜蛾在我国有东线和西线两条由南向北的迁飞路线,其中东线源于泰国、老挝、越南,进入我国华南地区后,逐步向北迁入长江流域、黄淮以及黄河以北地区;西线则源于缅甸,进入我国云南省后,经贵州、四川后,迁入山西、陕西、甘肃等地。从我国各地草地贪夜蛾中共生菌Cardinium感染情况比较来看,我国广东广州、云南保山和江苏无锡地区害虫体内Cardinium感染率均很高,而广西南宁地区未见Cardinium感染,说明江苏地区草地贪夜蛾来源于广东的可能性很大,广西和广东的境外虫源可能存在一定的差异。在草地贪夜蛾中发现的另一种共生菌Arsenophonus在云南种群中有一定的感染率,而在广东以及长江中下游的湖北和江苏地区均接近零感染。草地贪夜蛾在各地共生菌的差异为用轨迹分析方法预测出来的害虫东西线两条迁飞路径初步提供了一定的现实依据,但同一地区或者模拟同一路径上害虫共生菌感染的差异性,也反映出迁飞性害虫迁飞路径的复杂性,要进一步通过不同地区草地贪夜蛾共生菌的差异分析来明确害虫的详细迁飞路径,今后还需要对更多地区害虫共生菌进行具体分析。再从各地区草地贪夜蛾抗寒能力差异来看,尚未表现出与害虫东西迁飞路线的相关性。
关于共生菌Cardinium在昆虫中的功能,已报道它主要与宿主生殖调控相关。Cardinium和Wolbachia共感染能诱导烟粉虱雄性后代减少和细胞质不亲和,在寄生蜂Encarsiasuzannee中Cardinium也引起宿主昆虫的细胞质不亲和(Lvetal.,2020;Pilgrimetal., 2021)。而就Arsenophnus的功能,已发现其具有调控生殖和影响化学药剂和杀虫微生物敏感性等多种功能。已报道Arsenophnus在丽蝇蛹集金小蜂中可杀死雄性胚胎,导致其雌性化(Hugeretal., 1985),在褐飞虱中帮助宿主抗杀虫真菌绿僵菌(朱欢欢等,2017),还降低了褐飞虱对吡虫啉的敏感性(Pangetal., 2018)。为发掘共生菌新功能,本研究在比较发现我国不同地区草地贪夜蛾耐寒能力存在显著差异基础上,初步分析了Arsenophnus和Cardinium与宿主耐寒能力的相关性,但未发现其存在关联。Arsenophnus和Cardinium这两种共生菌在草地贪夜蛾中是否具有抗杀虫微生物以及调控生殖的功能?除此外,有无其它功能?均有待进一步的研究。
