部分蝽类昆虫翅表微纳结构与浸润性研究
2023-11-09孙明霞梁爱萍
朱 琳,孙明霞*,梁爱萍,2*
(1. 天津师范大学生命科学学院,天津市动物多样性保护与利用重点实验室,天津 300387; 2. 中国科学院动物学研究所,北京 100101)
自然界中种类繁多的生物经过亿万年的优胜劣汰、生存斗争,为了适应不断变化的环境,逐渐具有了与其生境高度适应的功能特性(任露泉和梁云虹,2010)。自然界中诸多动植物体表的多尺度形貌结构赋予其表面特殊的浸润性,自然界中“荷叶效应”就是最典型的现象,德国植物学家Barthlott和Neinhuist(1997)发现荷叶表面具有独特的微米级乳突结构,荷叶上的水珠可以自由滚动,带走叶面上的污垢从而达到自我清洁的效果。在水稻叶表面也发现液滴单向运输的现象,水稻叶片表面具有与荷叶类似的微纳复合突起结构,这些突起结构排列不均匀是其表面水滴各向异性滚动的主要原因(Fengetal., 2002)。除了荷叶、水稻、玫瑰(Fengetal., 2008; Fengetal., 2011)等植物上可以看到这种浸润现象外,昆虫翅表的浸润性也备受关注。昆虫是唯一一种有翅会飞行的无脊椎动物,昆虫翅膀经过数百万年的进化,已经成为自然界中最复杂的三维周期性介质结构材料之一,具有荧光、超疏水、抗反射和快速导热性等优良特性(Liuetal.,2017;Shahalietal., 2019)以及出色的飞行技能等多种不同的功能。1996年,Wagner等人(1996)对97种昆虫翅膀的浸润性进行了研究,浸润性与“SM指数”(SM Index,翅膀表面积与身体质量的商)之间存在相关性。高“SM指数”的昆虫,例如具有“大型翅膀”的蜻蜓目(弯艳玲等,2008)、脉翅目(Watsonetal., 2010, 2011)和许多鳞翅目(房岩等,2007;Zhengetal., 2007)都具有超疏水性的翅膀,并随着水滴的滚动来去除翅表的污物。而低“SM指数”的具有“小型翅膀”的昆虫,如膜翅目(韩宜伟,2017)和鞘翅目(Sunetal., 2012)则往往呈现出更多的亲水特性。浸润性表面在实际应用中,如自清洁材料、防雾化或防冰玻璃、防水织物和仿生物粘合剂表面的制备,需要对其表面性能如浸润性等进行精确的控制和工程设计(Wangetal., 2017; Chenetal., 2019; Patiletal., 2022)。具有微纳复合尺度形貌的生物表面结构已经通过多种技术成功地应用于仿生功能性表面的制造当中(Lietal., 2021; Huetal., 2022; Quetal., 2023; Wangetal., 2023)。而这些文章大多是针对同质的翅膀(整个翅膀均为膜质或均为鞘翅)进行的研究,而对异质的翅膀(整个翅膀既有膜质又有革质)的浸润性研究还尚未被报道。作为典型半翅目昆虫之一的蝽类,其翅表浸润性研究报道较少。蝽类昆虫属于六足亚门、昆虫纲、有翅亚纲、半翅目、蝽科,其前翅基半部革质,端半部膜质,为半鞘翅。多为植食性,刺吸植物茎叶或果实的液汁,常栖息于山地和平原,见于林缘、河谷、灌丛和草丘中、也常出没在石洞、树洞或倒木下。本文选择了20种不同生境的蝽类昆虫,对其前翅的革质、膜质区浸润性、微观结构、化学成分进行研究,研究结果可为不同机械性能材料浸润性研究提供参考。
1 材料与方法
1.1 实验材料
蝽类昆虫(如图1-A~F)标本于2021年7月采自重庆市(南川区)、贵州省(赤水市、习水县)和山东省(烟台市),隶属9科17属20种,共300头成虫标本。标本鉴定由昆虫分类学家帮助完成,采用系统分类法进行分类鉴定。

图1 蝽类成虫
1.2 实验仪器和方法
实验前,用流动的去离子水把20种蝽类昆虫前翅革质和膜质区(图1-G)清洗干净,用手术剪截取2 mm×2 mm尺寸的样品。将样品粘贴在载玻片上,应用KRUSS公司生产的DSA30S型全自动接触角测量仪采用坐滴法测量接触角,液滴大小为2 μL,温度为25±1℃。平行样本10个,计算其平均值及标准偏差。使用IBM SPSS Statistics 26软件,对20种蝽类昆虫翅表革质、膜质区接触角进行独立样本T检验,并对两组数据进行person相关性分析,采用OriginPro 2021制作柱形图。
将样品粘于扫描电镜样品柱上,在24 mA电流下,使用BAL-TEC SCD005型离子溅射仪喷金300秒处理,然后在FEI公司生产的Quanta 200型扫描电子显微镜下进行观测。并使用Image J软件测量微观结构参数,重复10次,计算其平均值及标准偏差。
3种蝽类昆虫翅表的化学物质成分应用Thermo Scientific公司生产的傅里叶红外光谱仪(FT-IR is50)进行分析,使用Origin 2021软件对红外光谱做后续处理。
2 结果与分析
2.1 接触角及数据分析
为了研究蝽类昆虫翅表浸润性,分别测量了20种蝽类翅表革质、膜质区的静态接触角,实验发现,蝽类昆虫翅膀表面接触角范围为40.66°~138.37°(见表1),按照蝽类昆虫翅表革质、膜质区的静态接触角的大小将其分为3类(图2)。其中紫蓝曼蝽Menidaviolacea、硕蝽Eurostusvalidas、茶翅蝽Halyomorphahalys、麻皮蝽Erthesiafullo、金绿宽盾蝽Poecilocorislewisi和小斑红蝽Physopeltacincticollis疏水性较强,革质区接触角分别为97.13°、102.56°、105.63°、111.52°和138.57°,膜质区为85.88°、97.57°、102.71°、95.63°、91.81°和126.82°,革质区比膜质区疏水性强。稻棘缘蝽Mictistenebrosa、泛刺同蝽Acanthosomanigrodorsum、曲胫侎缘蝽Cletuspunctiger和广腹同缘蝽Homoeocerusdilatatus呈现亲水性及弱疏水性,革质区接触角分别为40.66°、62.57°、68.88°和93.71°,膜质区为79.93°、74.76°、78.59°和104.66°,膜质区比革质区疏水性强。赤条蝽Graphosomarubrolineata、稻绿蝽Nezaraviridula、环斑猛猎蝽Sphedanolestessinicus、菜蝽Eurydemadominulus、荔蝽Tessaratomapapillosa、横带红长蝽Lygaeusequestris、珀蝽Plautiafimbricius、负子蝽Sphaerodemarustica、中稻缘蝽Leptocorisachinensis和斑须蝽Dolycorisbaccarum呈现亲水性及弱疏水性,革质区接触角分别为61.07°、64.68°、81.12°、81.91°、82.34°、83.80°、83.83°、91.16°、94.72°和100.95°,膜质区为61.32°、71.24°、77.24°、77.77°、80.65°、83.81°、81.45°、90.55°、93.00°和103.58°,革质区与膜质区疏水性趋近。

表1 20种蝽类昆虫翅表革质、膜质区的静态接触角

图2 三类翅表革质区和膜质区接触角图
采用SPSS软件对蝽类昆虫革质和膜质区接触角进行独立样本T检验,结果显示曲胫侎缘蝽、稻棘缘蝽、泛刺同蝽、紫蓝曼蝽、麻皮蝽、金绿宽盾蝽和小斑红蝽的革质和膜质区接触角具有极显著性差异(P<0.001),硕蝽和广腹同缘蝽的革质和膜质区接触角差异较显著(P<0.01),茶翅蝽的革质和膜质区接触角具有显著性差异(P<0.05),而另外的10种蝽类昆虫的革质和膜质区接触角无显著性差异(P≥0.05)。因此,根据静态接触角值和独立样本T检验结果分为革质区疏水性弱于、强于和趋于膜质区3类进行研究(图3)。

图3 20种蝽类昆虫翅表革质与膜质区接触角差异分析柱形图
2.2 翅表微观结构
经过扫描电子显微镜观测,20种蝽类昆虫翅表微观表面结构具有多样性。6种蝽类昆虫翅表革质区比膜质区的疏水性强,电镜观测发现紫蓝曼蝽和硕蝽革质区分布着长短不一的刚毛(图4-A,C),但密度较为稀疏,呈现较弱疏水性,膜质区仅有少量凸起(图4-B,D)。茶翅蝽革质区分布着不规则裂纹及少量分泌物,膜质区则布满皱纹(图4-E,F)。麻皮蝽革质区分布着大量无规则排列的凸起和分泌物,膜质区具有低密度的凸起(图4-G,H)。金绿宽盾蝽革质区具有蜂窝状六边形乳突结构,里面布满大小不一的圆柱形乳突,每个六边形结构由约29~40个乳突组成(高度0.903±0.286 μm,直径0.676±0.138 μm,间距0.995±0.141 μm)。膜质区则分布着大量不规则凸起,革质区的乳突结构使其疏水性明显强于膜质区(图4-I,J)。小斑红蝽的翅翼表面结构为圆柱形乳突,顶端近似球形,结构基部明显增大,革质区具有高约1.516±0.293 μm的乳突(直径0.702±0.170 μm,间距1.191±0.132 μm),膜质区具有高约1.227±0.122 μm的乳突(直径0.491±0.061 μm,间距0.655±0.158 μm),革质区比膜质区的乳突结构具有相对较大的高度、直径和间距,使革质区翅表疏水性更强(图4-K,L)。这些粗糙结构能有效减少与水的接触面积,有助于增大接触角,有效使蝽类昆虫翅表革质区具有更高的疏水性。4种蝽类昆虫翅表的膜质区比革质区更为疏水,翅表膜质区具有椭圆形突起和分泌物,且比革质区的相对密度更高。扫描电镜图像显示广腹同缘蝽和曲胫侎缘蝽的革质区和膜质区都分布着大小不一的椭圆形突起和分泌物,但革质区的突起密度较低,这些结构的低密度,将不足以抵抗水滴停留在结构之间的渗透,使水易于滑动到翅表上,疏水性较弱(图5-A,B,E,F)。稻棘缘蝽膜质具有凹凸不平的褶皱,而革质区较为平坦光滑(图5-C,D)。泛刺同蝽膜质区分布有不规则的椭圆形突起,疏水性较强,而革质区仅分布着不规则图案,较为平坦(图5-G,H)。

图4 6种蝽类昆虫翅表革质区和膜质区电镜图

图5 4种蝽类昆虫翅表革质区和膜质区电镜图
而在10种蝽类昆虫中,翅表革质、膜质区疏水性没有显著差异,如图6中的赤条蝽、荔蝽、珀蝽、负子蝽、中稻缘蝽和斑须蝽疏水性较弱,扫描电镜显示革质区和膜质区的翅面结构相似,仅具有大小不一的隆起和分泌物,表面较为平坦,无明显粗糙结构。

图6 6种蝽类昆虫翅表革质区和膜质区电镜图
2.3 化学成分分析
采用傅立叶红外光谱仪对3种蝽类昆虫(小斑红蝽、荔蝽、曲胫侎缘蝽)前翅革质和膜质区表面成分进行了定性分析,这3种蝽类昆虫翅表革质区疏水性分别强于(小斑红蝽)、弱于(曲胫侎缘蝽)和趋于(荔蝽)膜质区(见表1),探究其表面化学成分对不同翅面疏水性的影响。分析结果显示红外光谱特征峰相近,整体结构相似。在3 280、2 920、2 850、1 621、1 540、1 400、1 240、1 160、1 060 cm-1处有吸收峰(图7)。3 280 cm-1处的吸收带由酚类、醇类或羧酸的O-H、N-H基伸缩振动引起;2 920 cm-1、2 850 cm-1处的吸收带显示翅表面含有脂肪烃结构;1 621、1 540 cm-1处的吸收带显示翅表面含有苯环结构;1 240、1 160、1 060 cm-1处的吸收带是由酸酐、酰氯、酯、醛、酮、羧酸、酰胺官能团的C-O基伸缩振动引起。蜡质层是长链烃类、脂肪酸、醇,以及酯(由长链醇和长链脂肪酸形成)等组成的混合物,所以其红外光谱在此处有明显的吸收峰存在,表明蝽类昆虫翅表分布着长链烃类、脂肪酸酯和脂肪酸醇构成的蜡质层。图中红色实、虚线分别表示小斑红蝽革、膜质区;绿色实、虚线分别表示荔蝽革、膜质区;蓝色实、虚线分别表示曲胫侎缘蝽革、膜质区。

图7 小斑红蝽、荔蝽和曲胫侎缘蝽翅表革质区和膜质区红外光谱图
2.4 疏水机理分析
接触角(Contact angle)是指在固、液、气三相交界处,自固-液界面经过液体内部到气-液界面之间的夹角,即从液固气三相态相交点作气-液界面的切线,该切线与液滴及平面之间的夹角,通常用θ表示。利用接触角能直观地衡量液体对固体的浸润程度。
1805年,Young提出了液滴与光滑固体表面的接触模型,如图8-a所示。当液滴在光滑固体表面达到平衡时,固-气、固-液、液-气界面张力与接触角之间的关系方程,即Young’s方程(Young, 1805),它是研究所有浸润现象的定量理论基础,见式(1)。式中γSG为固-气的界面张力、γSL为固-液的界面张力、γLG为液-气的界面张力。然而,实验发现该模型的最大接触角只能达到120°左右,这与超疏水表面接触角大于150°的事实不相符,因此Young’s模型只适合光滑均匀的理想模型。
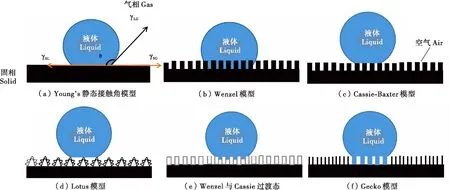
图8 6种固液接触型(Wang &Jiang,2007)
(1)
现实中大多数固体表面是非光滑的,布满各种形状的粗糙结构,例如蛾翅表的鳞片结构(张建军,2008),蝉翅表面的乳突结构(Jessicaetal., 2020)。现阶段除了Young’s 模型外,还有另外5种模型来描述水滴与材料表面为形貌结构的接触状态:Wenzel模型、Cassie-Baxter模型、Lotus模型、Wenzel与Cassie的过渡态模型以及Gecko模型(图8)。
Wenzel(1936,1949)认为由于表面粗糙结构的存在,液体始终会将表面上的凹槽结构填满,增加了固体表面真实的表面积,使得实际固液接触面积大于表观几何接触面积(图8-b),导致疏水性的增强,建立了Wenzel方程,见式(2)。
cosθ=γcosθe
(2)
式中:θ代表液体在粗糙表面的表观接触角;θe代表液体在理想的光滑平面上的真实接触角(本征接触角),由固体表面的化学组成决定。γ定义为粗糙度因子,也就是实际固液接触面面积与本征固液接触面面积的比值。
Wenzel方程不适用于解释由不同种类的化学物质组成的固体表面(即异质表面)。Cassie和Baxter提出粗糙的表面可以被视为不均匀的固体表面,当固体表面高度疏水时,假设液滴无法填满非光滑表面的凹槽,表面的沟槽中会滞留一部分空气,使得凹槽处的液体无法与固体表面直接接触,这导致液滴与固体表面的接触变成一部分液体与固体表面直接接触,还有一部分的液体会与凹槽内的空气接触。液滴在这种微纳米级粗糙结构表面上的接触是一种固-气-液复合接触,即复合润湿状态。这种微纳米级的表面可以吸附空气,并在其表面形成一层空气膜。空气膜的存在使液滴不能填满粗糙表面上的缝隙和凹槽,从而导致在水滴的下面截流有空气,表观上的固液接触实际上由固-液、固-气共同组成(图8-c),并提出了一种液滴在固-液-气复合界面计算接触角的方程(Wangetal., 2009; Barthlottetal., 2017),见式(3)。
cosθ=φs(1+cosθe)-1
(3)
式中:φs代表固体在复合接触面中所占面积的百分比值,0<φs<1。
江雷等人(2007)根据疏水表面不同的接触角滞后,认为疏水表面状态还包括Lotus状态、Wenzel与Cassie的过渡态以及Gecko状态等3种(图8-d~f)。Lotus状态指的是指具有大接触角和极小滚动角的Cassie状态的一种特例,这种具有类似荷叶表面结构的自清洁效应(图8-d)。此外,还存在一种Wenzel与Cassie的过渡态,液滴在外力作用下会改变其浸润状态,变为如图8-e所示的浸润状态,即产生一种叫做过渡态的浸润状态的转换。当过渡态发生时,接触角减小,表面对水滴的黏附力增加,水滴可以在表面以一定倾斜角度滑动,因此当前超疏水表面理论研究的重点是如何设计最优的表面结构,避免产生过渡态。Gecko状态是指接触角较高、粘附性较高的一种状态(图8-f)。
本研究的蝽类昆虫中小斑红蝽的疏水性较强,如图9所示的小斑红蝽翅膀上非光滑疏水表面的模型,其特征参数R,h,d分别表示圆柱半径、圆柱高度、圆柱间距,则φs=πR2/d2,表面粗糙度因子γ=(d2+2πRh)/d2。

图9 小斑红蝽翅表革质部分柱状乳突结构模型
小斑红蝽翅表革质和膜质区接触角分别为138.37°和126.82°,具有较高疏水性,革质区具有高约1.516±0.293 μm的乳突(直径0.702±0.170 μm,间距1.191±0.132 μm),膜质区具有高约1.227±0.122 μm的乳突(直径0.491±0.061 μm,间距0.655±0.158 μm),蝽类昆虫翅表主要由蜡质晶体(Waxcrystals)构成,水滴在其光滑表面的本征接触角约为105°(Wan, 2009)左右,θe取105°时,由翅表面柱状乳突结构参数可知R和d值,计算出φs和γ,利用Wenzel模型(式(2))、Cassie模型(式(3))计算出翅表革质区理论接触角分别为67.04°和142.94°,与实际所测相对误差分别为51.55%和3.30%。翅表膜质区理论接触角分别为93.44°和132.29°,与实际所测相对误差分别为 26.32%和4.31%。因此认为小斑红蝽翅表微观结构浸润模型满足Cassie-Baxter模型,翅表面微米级柱状突起的间隙被空气占据,柱体与水滴之间稳定地吸附一层气膜,水滴无法进入凹槽,不能与翅表面充分接触(只与乳突顶端接触)形成复合接触,因此水滴接触时可产生Cassie-Baxter状态,进而产生相对最大的接触角。
生活在不同栖息地的昆虫其翅表浸润性有所差异(Sun, 2012; Junhoetal., 2017),本文研究的蝽类昆虫生活在不同的生境中,小斑红蝽、金绿宽盾蝽、茶翅蝽、麻皮蝽、硕蝽、紫蓝曼蝽、斑须蝽和泛刺同蝽主要寄生于植物中上层,例如果树(苹果、梨、桃、葡萄、柑橘和山楂树等)和林木(柳、松、白栎、梧桐树等)。活动于植物中上层的这些蝽类昆虫翅表疏水性较强,接触角为91.81°~138.67°,翅表具有乳突、不规则凸起和皱纹等粗糙结构从而导致了高接触角。这些蝽类昆虫翅表呈现较高疏水性使其可以寄生于植物中上层,雨天不至于从高处跌落,并能带走翅表颗粒污染物,丰富了蝽类昆虫的食物来源,从而增加蝽类昆虫物种丰富度。菜蝽、珀蝽、赤条蝽、稻棘缘蝽、广腹同缘蝽、曲胫侎缘蝽、中稻缘蝽、横带红长蝽、稻绿蝽、环斑猛猎蝽和稻棘缘蝽主要寄生于植物中下层,例如水稻、小麦、花生、玉米、白菜、大豆和萝卜等。活动于植物中下层的这些蝽类昆虫翅表呈现亲水性及弱疏水性,接触角为40.66°~94.72°,翅表具有大小不一隆起和分泌物结构,但这些结构的低密度性使其呈现亲水性及弱疏水性。雨天时翅表常被雨水打湿,但仍可以掉落于杂灌木等植物下层,可在草丛爬行活动。负子蝽常悬浮池塘或湖泊静水中,接触角为90.55°,电镜下呈现相对光滑的表面,它们翅表光滑的表面可以归因于它们生活在水生环境中。
研究指出,形貌结构、化学成分和生境是决定材料表面呈现疏水特性的关键因素(Blossey, 2003),小斑红蝽翅表因表面覆盖的蜡质层使液滴与其表面接触由原来的几丁质(θe=95°)变为蜡质晶体(θe=105°),疏水性提高10.53%。且其寄生于毛竹、油桐和柑桔等植物中上层。此外,表面微形貌结构及其产生的粗糙度能够增强材料的疏水性(Neelesh, 2004; Kochetal., 2009; Kochetal., 2013),乳突结构的存在,形成了液滴与翅膀表面的复合式接触(接触角为138.37°),满足Cassie模型。且栖息于植物中上层的蝽类昆虫疏水性较强。因此蝽类昆虫翅表面的高疏水性是微米级粗糙结构与蜡质晶体及生境协同作用的结果。
3 结论与讨论
本文以20种蝽类昆虫为研究对象,通过研究其前翅的革质、膜质区域浸润性、微观结构和化学成分,分析了部分疏水性强的蝽类昆虫疏水机理,结果显示:(1)蝽类昆虫翅表浸润性存在明显差异,接触角为40.66°~138.37°。(2)6种蝽类昆虫翅表革质区比膜质区的疏水性强,具有乳突、不规则凸起和皱纹等粗糙结构。特别是柱状乳突结构(直径0.702±0.170 μm,高度1.516±0.293 μm,间距1.191±0.132 μm)有效地使革质区表面更疏水。相反,4种蝽类昆虫翅表的膜质区比革质区更疏水,椭圆形突起和分泌物密度更高。而在10种蝽类昆虫中,疏水性没有显著差异,并且革质和膜质区的翅面结构相似,仅具有不规则的图案和分泌物。(3)蝽类昆虫翅表分布着长链烃类、脂肪酸酯和脂肪酸醇构成的蜡质层。(4)理论分析结果显示,部分疏水性强的蝽类昆虫例如小斑红蝽翅表因具乳突结构而能够截留空气,使水滴较易产生Cassie-Baxter接触状态,从而呈现相对最大的接触角。且栖息于植物中上层的蝽类昆虫疏水性较强。蝽类昆虫不同硬度的翅表的浸润性归因于微米级微观结构和表面蜡质层的联合效应及生境的协同作用。这一机理为不同机械性能材料浸润性研究提供参考。
致谢:感谢南开大学生命科学学院穆怡然老师、天津工业大学分析测试中心武春瑞老师、天津师范大学生命科学学院张骁博士在本研究中给予的支持与帮助。
