红花酢浆草与三叶鬼针草的种内及种间密度制约机制研究
2023-11-01陈凤仙冯鑫鑫张宏伟乌俊杰
陈凤仙,冯鑫鑫,张宏伟,乌俊杰
大理大学农学与生物科学学院,云南大理 671003)
入侵植物能在自然或半自然的生态系统中繁殖、扩散,并对本地植物造成威胁,严重时甚至会改变本地的生物多样性。近年来,随着全球气候变暖和人类干扰活动的增加,植物的扩散变得更加频繁和广泛,这使得植物入侵的风险日益增加[1]。
云南省地处我国西南高原,地理位置优越,气候类型丰富,同时与多个国家接壤,人类活动频繁,对外交流和进出口贸易历史悠久,且具有多种生态系统类型,受自然生态系统和植被的干扰较大,这使得云南成为外来物种入侵的“重灾区”。外来入侵物种在破坏生态系统以及生物多样性的同时,还会造成生物污染和一定的经济损失[2]。该试验选择红花酢浆草(Oxaliscorymbosa)和三叶鬼针草(Bidenspilosa)进行试验,一方面红花酢浆草和三叶鬼针草对云南植物入侵情况较为严重[3],另一方面,2种植物在田间常见但易被忽视其入侵性。
红花酢浆草原产中南美洲,本作为一种观赏性植物引入,现逃逸为田间杂草。其适应性、抗逆性、侵占性和再生能力强,繁殖及扩散迅速,是典型的旱性、恶性杂草[4]。2007年我国将其列为检疫性杂草,目前在全国广泛分布,常被人忽略其入侵性。研究表明,红花酢浆草会对花草果蔬等多种作物的萌发及生长产生抑制作用[5-6]。三叶鬼针草原产中美洲,主要通过种子的倒刺黏附于人畜或货物进入我国,现已经广泛分布在我国华南、华中、华东、西南等地[7]。三叶鬼针草具有强大的入侵扩展机制,可通过高繁殖力和散布力、化感作用和改变土壤微生物结构等方面实现其快速入侵扩张的目的[8]。但目前,对三叶鬼针草的研究大多在重金属的富集作用以及药用价值上[8-9]。忽视红花酢浆草和三叶鬼针草的入侵性,可能导致其更大范围的扩散繁殖,从而进一步危害农林业和畜牧业。
种群密度是植物种群普遍存在的特征之一,而密度制约则是种群调节的一个重要机制[10]。植物密度变化受到个体间竞争的影响,竞争强度随植物发育阶段的变化而变化[11-12],且草本植物相比木本植物对密度压力更敏感[13]。在密度压力下,植株会显著改变自身的株型发育系统,并影响各器官间的异速生长,进而权衡器官生物量分配以完成生活史[14]。植物功能性状可以反映植物资源获取和适应环境变化的生存策略[15]。叶片对环境变化较敏感,可塑性大,植物可以通过改变叶片的各种特征来适应环境的变化[13]。通过观察和测量不同密度下叶片的功能性状,可以反映植物个体的生存策略是否会随着种群密度而发生变化。因此,探讨不同密度下植物的生长和叶片功能性状指标,对于揭示植物的环境适应性和生存策略具有重要意义。
入侵植物表现出的高度生长率、繁殖力和可塑性,多是针对本地种得出的。而不同入侵植物之间的比较研究对防治植物入侵也很重要[16],但这种研究现在仍不多见。2种入侵植物同时定居在同一空间时会对彼此产生什么影响,入侵植物之间由于彼此的化感作用是否会抑制彼此的生长和入侵。为此,笔者以红花酢浆草和三叶鬼针草为对象,进行不同密度的单一种植和混合种植,测量植物的生长和叶片功能性状指标,探索同种入侵植物间的密度制约机制以及入侵植物间的相互作用机制。
1 材料与方法
1.1 试验地概况试验样地设置在云南省大理白族自治州大理市大理大学实验基地(100°09′17″E、25°40′20″N),属于北亚热带高原季风气候类型。试验样地中的土壤理化性质均一,可以排除土壤异质性的影响。红花酢浆草和三叶鬼针草成熟采集后在室温下保存。
1.2 试验方法设置低密度(300粒/m2)、中密度(900粒/m2)和高密度(1 800粒/m2)3个播种密度,有2种种植方式(单一种植和1∶1混合种植),各3次重复。样方为边长1 m的正方形,共27块样方,样方间隔0.5 m,以防边缘效应。采用完全随机区组设计。于2021年7月15日播种,整个试验过程中去除样方中的其他植物。
2021年9月15日采集样品。每个样方内选择长势健康且能代表整块区大致长势的植株,单一种植的样地取5株,混合种植的样地红花酢浆草和三叶鬼针草各取5株,共计采集180株植株。先测量株高、叶绿素含量,然后采取全株挖掘法获取整株,将样品带回实验室。除去叶表面尘土,用水冲洗根系并快速擦干表面水分。每株选取健康完整的3片叶片,用叶绿素仪(SPAD-502 Plus)测量每片叶的叶绿素含量,均测量3次,以降低误差,3片叶的平均值代表该植株相应指标的数值。再用外径千分尺测量选取的这3片叶的厚度,在每片叶片的近叶尖处、叶片中间和近叶柄处分别进行3次测量,避开较大的叶脉,测量的平均值代表该叶片的厚度。用分析天平(精确度均为0.001 g)测量根重、单片叶重、地上鲜重。待以上数据收集完后,将所有采集植株放置于信封中,放入80 ℃烘箱内烘干72 h以上至恒重后称重。分别计算叶干物质含量(leaf dry matter content,LDMC)和根冠比,计算公式:
叶干物质含量=叶片干重(g)/叶片鲜重(g)
根冠比=根生物量(g)/地上部分生物量(g)
1.3 数据处理数据采用单因素方差分析(One-way ANOVA)和t检验;数据分析及图表均在Excel 2016和SPSS 24.0中完成。
2 结果与分析
2.1 密度对红花酢浆草的生长和叶功能性状的影响从表1可以看出,随着密度升高,红花酢浆草的各项指标均发生了显著变化(P<0.05)。红花酢浆草的萌发率在不同播种密度间有差异,在中密度下萌发率最低。在单一种植方式下,随着密度的升高,株高和叶绿素不断增加,叶片厚度降低,干物质含量(LDMC)无显著变化。在中密度(900粒/m2)下,红花酢浆草具有最低的根冠比和最高的总干重。在混合种植方式下,株高和叶绿素随着密度的升高而升高,叶片厚度、LDMC和总干重总体随着密度的升高而降低,根冠比在不同密度下无显著差异。综上所述,密度对红花酢浆草的生长产生了显著影响,随着密度增加,红花酢浆草的存活状态下降,表现出明显的负密度制约现象。
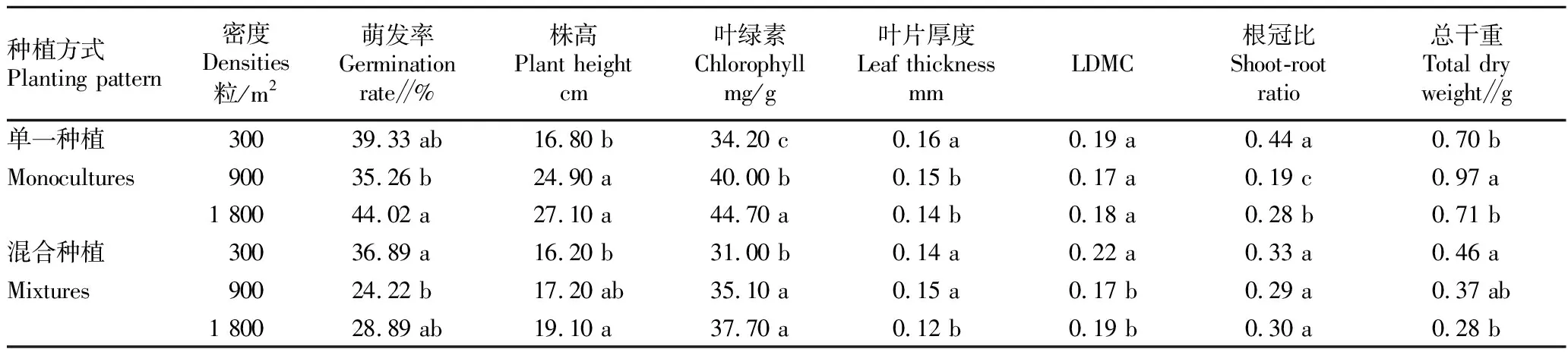
表1 3种密度下单一种植和混合种植的红花酢浆草的各项指标
2.2 种植方式对红花酢浆草的生长和叶功能性状指标的影响由图1可知,株高和叶绿素含量在中高密度下均表现出混合种植显著低于单一种植。叶片厚度在低密度和高密度下也表现出混合种植显著低于单一种植。LDMC在低密度和高密度下表现出混合种植显著高于单一种植。根冠比在低密度时为单一种植显著高于混合种植,在中密度时表现为混合种植显著高于单一种植。总干重在中高密度下均表现出混合种植显著低于单一种植。种植方式对红花酢浆草的影响主要表现在中高密度下。从株高、叶绿素、叶片厚度、LDMC和总干重方面,均表现为在混合种植下,红花酢浆草的生长受到了更强的抑制作用,表明三叶鬼针草对红花酢浆草的抑制作用大于红花酢浆草同种间的负密度制约作用。
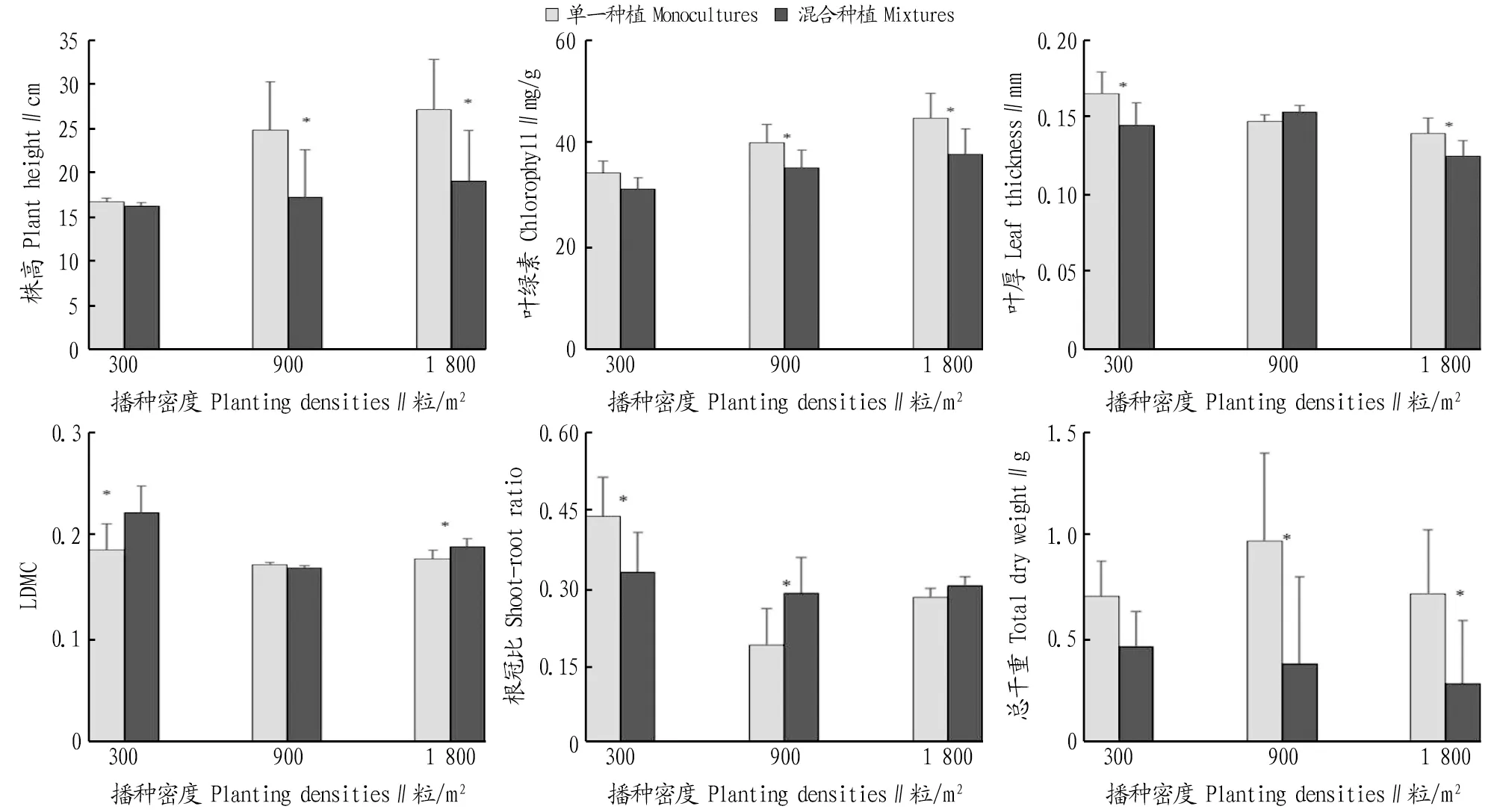
注:*表示不同种植方式间指标差异显著(P<0.05)。Note:* indicated significant difference between different planting patterns at 0.05 level.图1 单一种植和混合种植红花酢浆草的生长和叶功能性状指标的比较Fig.1 Comparison of indices of Oxalis corymbose under monoculture planting patterns and mixture planting patterns
2.3 密度对三叶鬼针草的生长和叶功能性状的影响从表2可以看出,随着密度升高,三叶鬼针草的各项指标之间均有显著差异(P<0.05)。三叶鬼针草的萌发率在不同播种密度间无显著差异。在单一种植方式下,随着密度升高,叶片厚度降低,叶绿素、根冠比和总干重无显著变化。在中密度(900粒/m2)下,株高和干物质含量(LDMC)最高。在混合种植方式下,株高、叶绿素、叶片厚度和总干重随着密度的升高而降低,根冠比总体随着密度的升高而升高,LDMC在不同密度之间无显著差异。由此可知,密度对三叶鬼针草的生长生存产生了显著影响。在中密度下,三叶鬼针草表现出良好的生长状态,随着密度增加,三叶鬼针草的存活状态下降,表现出明显的负密度制约现象。
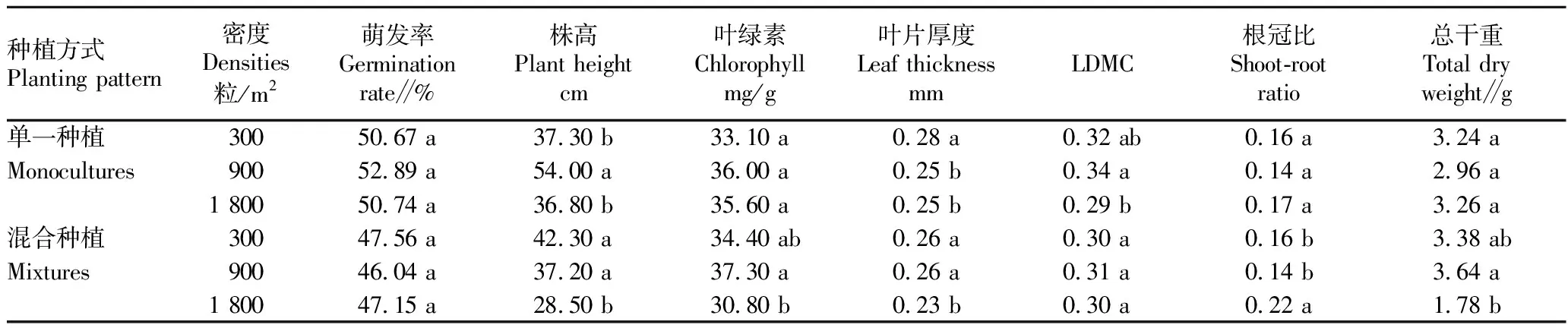
表2 3种密度下单一种植和混合种植的三叶鬼针草的各项指标
2.4 种植方式对三叶鬼针草的生长和叶功能性状的影响采用t检验的方法对同一密度下单一种植和混合种植2种种植方式间进行差异性分析。株高、叶绿素含量和LDMC在2种种植方式下均无显著差异。叶片厚度和总干重在高密度下也表现出混合种植显著低于单一种植。根冠比在高密度时表现为混合种植显著高于单一种植。由图2可知,种植方式对三叶鬼针草的影响主要在高密度时表现出来。在高密度下,三叶鬼针草在混合种植下受到了更强的抑制作用,表明红花酢浆草对三叶鬼针草的抑制作用大于三叶鬼针草同种间的负密度制约作用。
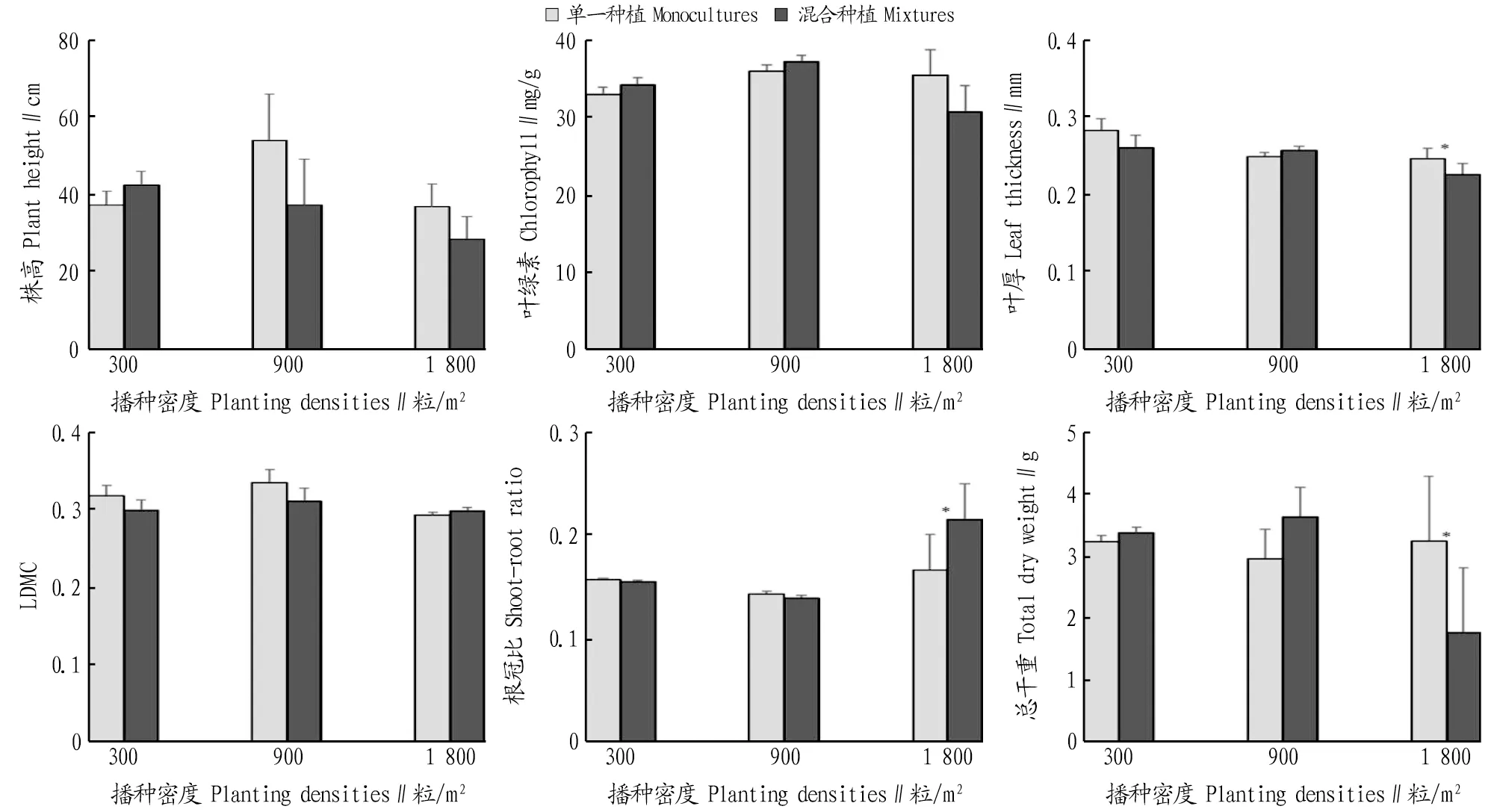
注:*表示不同种植方式间指标差异显著(P<0.05)。Note:* indicated significant difference between different planting patterns at 0.05 level.图2 单一种植和混合种植三叶鬼针草的生长和叶功能性状指标的比较Fig.2 Comparison of indices of Bidens pilosa under monoculture planting patterns and mixture planting patterns
3 讨论
3.1 负密度制约抑制入侵植物的生长发育种内竞争的实质是对光、水分、矿质营养和空间资源的竞争,在低密度下,种群内相邻植株间不存在竞争或竞争很弱,随着密度的升高,种内竞争会变得越来越激烈[17]。密度制约主要是通过影响相邻植物间器官对资源的竞争强度来调节资源分配,导致各器官的生物量分配比例发生变化[18-19]。
光是生境中重要的环境因子之一,种群密度引起的植株邻近体间遮阴效应将直接影响光照强度、光照时间和光质,从而促使植物调整资源利用和分配策略。红花酢浆草和三叶鬼针草的叶绿素含量和株高均随密度的升高而增加。这表明2种入侵植物应对高密度的策略是提高自身的高度以及提高叶绿素含量,这主要是为了竞争光资源,增加对光资源的获取能力。研究表明,四季竹(Oligostachyumlubricum)高度有随着密度增加而增高,以获得更多的光资源[20]。紫茎泽兰(Eupatoriumadenophorum)和兰花菊三七(Gynurasp.)随着光照强度的减弱,表现为增加高度以截获更多光能[21]。红花酢浆草和三叶鬼针草的叶片厚度均随密度的升高而降低。在混合种植方式下,红花酢浆草和三叶鬼针草的总干重也呈随密度升高而降低的趋势。北美车前(Plantagovirginica)也会随着密度的增加,单叶重和单株总生物量降低[19]。随着密度增加,加剧了个体之间的资源竞争,会导致植物体生物量下降[10]。种群密度变化导致的光资源差异及个体间密度制约效应,这可能是红花酢浆草和三叶鬼针草生物量及叶片功能性状发生变化的重要因素之一[22]。
综上所述,入侵植物红花酢浆草和三叶鬼针草的生长和功能性状指标随着种群密度的增加呈现规律性变化,表现出同种间的负密度制约效应。在高密度压力下即使是入侵植物,也会受制于密度带来的竞争压力,呈现出“竞争-密度效应”[10]。
3.2 红花酢浆草和三叶鬼针草之间存在相互制约的现象红花酢浆草和三叶鬼针草均是一年生草本植物,该试验发现混合种植相比于单一种植,2种植物均受到更强烈的抑制作用,说明2种入侵物种间直接存在相互制约作用,且负效应强度高于同种负密度制约的强度。
在900粒/m2(中密度)时,三叶鬼针草受红花酢浆草的负效应制约不明显,但红花酢浆草已明显受到三叶鬼针草的抑制作用,说明红花酢浆草对三叶鬼针草的抑制作用更敏感。三叶鬼针草的水浸提取液、叶片挥发物和根系分泌物会抑制其他植物种子的萌发、幼苗生长和光合作用[23-24]。只有在高密度下,三叶鬼针草在混合种植下才表现出叶片厚度和总干重呈大幅度下降的趋势,说明三叶鬼针草也同样受红花酢浆草的负作用。研究表明,红花酢浆草的水提液对三叶鬼针草具有较强的化感作用,会抑制三叶鬼针草的种子萌发和幼苗生长[25-26]。由此可知,红花酢浆草更容易受到三叶鬼针草的抑制作用,原因之一有可能是三叶鬼针草在中国为I级入侵等级,为恶性入侵种[27],具有较强的抗逆性和繁殖力[28],入侵性更强,对红花酢浆草有更为强烈的他感作用。
综上所述,红花酢浆草和三叶鬼针草之间具有相互抑制的现象。在一种入侵植物泛滥的生境中,人为传播在本地泛滥的其他入侵植物,是否会使入侵植物之间产生相互抑制的现象,从而为本地物种的生存提供条件,这是下一步需要研究的方面。另外,2种入侵植物之间的相互制约是否会影响其种子的发育繁殖,进而影响二代种子的萌发,这方面有待进一步研究。
4 结论
在全球变化背景下,云南省作为我国的边境省份长期受到植物入侵的影响。红花酢浆草和三叶鬼针草常见于农田和绿化带,严重危害花草果蔬等经济作物及农作物的生长,是易被人们所忽视的入侵植物。为了探究红花酢浆草和三叶鬼针草的入侵性问题,该试验采用完全随机区组设计对2种入侵植物进行不同密度的单一种植和混合种植。结果发现,红花酢浆草和三叶鬼针草同种个体间均具有负密度制约机制。2种入侵植物间具有强烈的负效应,且高于同种负密度制约。入侵植物间强烈的负作用有利于相互制约彼此的生长和入侵,此结果对防治入侵植物具有一定的指导意义。
