华北落叶松人工林天然更新幼苗的空间分布特征
2023-10-30赵伟文孙雁君高玉凤
赵伟文,孙雁君,高玉凤
(山西农业大学 水土保持科学研究所,山西 太原 030013)
林木群落的空间格局能够揭示林分历史、种群动态和竞争状况,同时能够反映树木在水平上的相对位置,是立地微环境、种子扩散方式、非生物因素干扰共同作用的空间结构。研究林木空间结构能够定量分析种群的水平空间格局,还可以揭示空间格局形成因素,阐明种群及林木的空间动态变化,并进一步了解种群形成的生态学过程[1]。目前,针对华北落叶松的研究多集中在土壤养分、物种多样性等方面,对华北落叶松大面积天然更新幼苗分布区空间分布格局的研究较少,研究华北落叶松人工林天然更新幼苗的空间分布特征有助于揭示林木天然更新机制。1977年,Ripley等[2]提出了Ripley’ K函数,由此空间点格局分析法应运而生,这是一套用于分析离散点的空间分布的工具,它是在森林生态学中使用最广泛的描述空间格局的方法之一[2],能最大程度地利用植物二维坐标系特征信息,在不同尺度下分析种群的空间格局,该方法已在植物生态学研究中被广泛应用[3-4]。随着研究的深入,学者们将基于网格的方法扩展到点格局分析中,以处理有限大小和不规则形状的对象,其本质思想是用单元大小小于植物大小的分类格栅地图来表示研究区内的植物,根据植物大小和形状在地图上由一个或几个相邻的网格单元进行表示,对有限大小和不规则形状的对象有了明确的考虑[5]。点格局分析法以每株树木的空间坐标为基本数据,所有个体构成空间二维点图,进行空间拟合后,可以最大限度地解释森林天然更新动态变化过程。空间点格局函数g(r)函数由K函数演变而来[6],g(r)消除了取样面积大小的制约,以物种个体在空间点位置的坐标为基准,以个体为中心做圆,通过统计落在圆形面积区域内点的个数来进行分析,该方法经过修正后可以实现在任意尺度下反映种群的聚集程度[7]。拓锋等[8]研究发现,云杉在小尺度研究上的分布格局主要受自身生物学特性的影响,在大尺度研究上的分布格局主要受生境异质性影响。池森等[9]通过对次生林幼树空间分布格局的研究表明,郁闭度、树种组成、林隙大小和林龄是影响幼树空间分布变化的主要因素。郝珉辉等[10]通过对吉林蛟河针阔叶混交林的研究表明,林分水平上的大树位置分布会对其与幼树生长的空间关联性产生影响。本文以关帝山华北落叶松人工林为研究对象,分析了华北落叶松天然更新幼苗的空间分布特征,并通过空间点格局方法探讨了华北落叶松大树和天然更新幼苗的分布特征以及两者在空间上是否存在一定的关联性,以期为后期人工林天然可持续更新提供理论依据。
1 研究区概括与方法
1.1 研究区概况
本试验研究区位于山西省吕梁市关帝山庞泉沟国家级自然保护区(37°46′~37°55′N,111° 22′~111°33′E),地处吕梁山脉中段,娄烦县、交城县和方山县三县的交界处[11]。海拔介于1300~2830 m。区域内四季分明,属暖温带季风大陆性气候,年平均气温为4.1 ℃,年平均相对湿度为71%,年平均降水量为822 mm,年平均蒸发量为1300 mm,无霜期为180 d,年平均水汽压为936.6 hPa。区域内土壤类型主要是山地褐土、褐土和棕壤土[12],平均土壤层厚度为75 cm,腐殖质层的厚度为9~10 cm[13]。
乔木种类有辽东栎(Quercus mongolica)、油松(Pinus tabuliformis)、华北落叶松(Larix principisrupprechtii)、青扦云杉(Picea wilsonii)和白扦云杉(Picea meyeri)等;灌木种类主要有美蔷薇(Rosa multiflora)、茶条槭(Acer tataricum)、铁线莲(Clematis florida)、黄芦木(Berberis amurensis)、黄刺玫(Rosa xanthina)、金露梅(Potentilla fruticosa)和银露梅(Potentilla glabra)等;草本种类主要有东方草莓(Fragaria orientalis)、老鹳草(Geranium wilfordii)、青蒿(Artemisia caruifolia)、毛茛(Ranunculus japonicus)和歪头菜(Vicia unijuga)等。
1.2 研究方法
1.2.1 样地设置 在关帝山庞泉沟国家级自然保护区柴逯沟华北落叶松人工林更新幼苗典型分布区设置12块面积为400 m2(20 m×20 m)的样地,样地内只分布有华北落叶松大树及更新幼苗。采用罗盘仪和皮尺对样地进行布设。布设完成后用手持GPS(G120,中国)记录各个样地经纬度、海拔,用水平仪测量样地坡度,用罗盘仪测量样地坡向(表1)。
1.2.2 数据采集
1.2.2.1 华北落叶松大树基础数据采集 对每块样地内的华北落叶松大树进行每木检尺,记录胸径、树高、冠幅和枝下高,标记每株大树的相对位置(即X/Y坐标)[14],并记录和计算各样地的郁闭度。在样地内随机选取3株长势良好的大树,利用生长锥(CO250,瑞士)对树芯进行采集[15],并记录树龄。
1.2.2.2 华北落叶松更新幼苗基础数据采集 在本研究中,根据实际情况将华北落叶松高度小于1.3 cm的苗木定义为更新幼苗。对样地内华北落叶松更新幼苗进行每木检尺,用游标卡尺测量地径,用皮尺测量树高和距离华北落叶松大树的相对位置(即X/Y坐标),样地大树与更新幼苗的空间分布如图1所示。
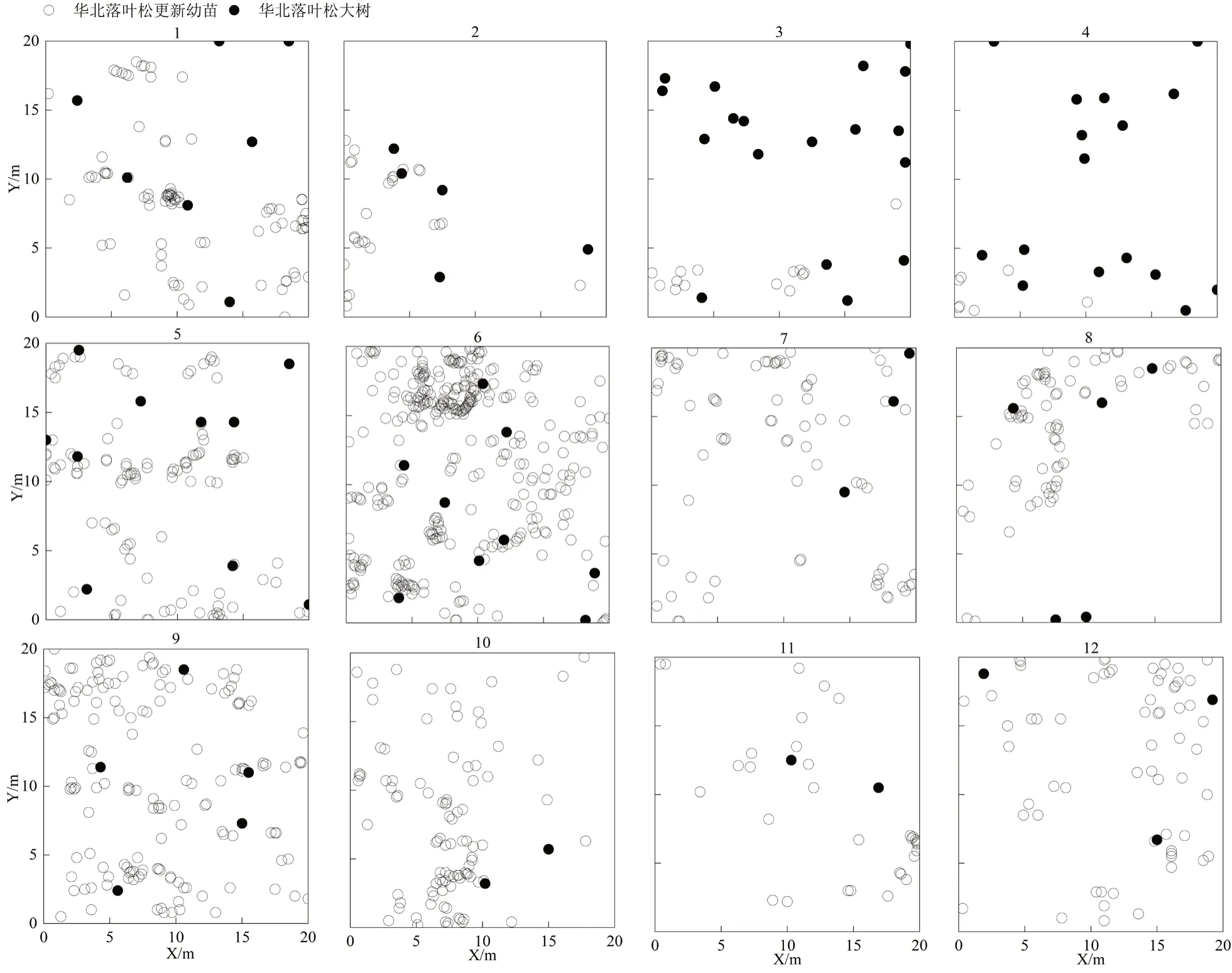
图 1 研究区样地内华北落叶松大树和更新幼苗个体的空间分布图
1.2.2.3 林下植被及枯落物调查 在样地内采用“X”型采样法设置5个5 m×5 m的样方,记录灌木种类和盖度。在灌木采集样点附近设置1 m×1 m的草本调查样方,记录草本种类及盖度。在草本样点附近设置30 cm×30 cm的小样方,记录枯落物层的厚度。
1.2.3 数据分析
1.2.3.1 种群空间分布格局 以Ripley在1977年提出的Ripley’s K函数K(r)为基础[2],采用单变量成对相关函数g(r),以样地中每个植株的空间坐标为基础,分析华北落叶松个体在不同尺度下的分布格局。其中,K(r)函数的计算公式为[2,16]:
式(1)中,a为样地面积大小;N为样地内林木树株数;dij为个体i到个体j之间的距离;r为空间尺度,以目标树为圆心的取样圆半径;Wij为边缘效应矫正系数,以点i为圆心、dij为半径的圆落在样地面积a中的弧长与自身圆周长的比值[8];Ir(dij)为指示函数,当dij>r时,Ir(dij)=0;当dij≤r时,Ir(dij)=1。
g(r)函数由K(r)函数演化而来,与K(r)函数相比,g(r)函数用圆环代替K(r)函数中的圆,分析圆环内所有树木的多度,能够有效消除小尺度上的累积效应,通过计算点在某一距离出现的频数来分析空间格局,使得分析结果更为可靠,因而g(r)函数在评估聚集程度方面得到了充分应用[17]。g(r)函数的表达式为:
式(2)中,若g(r)=1,则种群在r尺度上呈随机分布;若g(r)<1,则种群在r尺度上呈均匀分布;若g(r)>1,则种群在r尺度上呈聚集分布[18]。
1.2.3.2 种群空间关联分析 2个不同径级个体之间的空间关联性分析,实际上是不同径级个体之间的多变量点格局分析,也被称为多元点格局分析[19]。采用双变量成对相关函数g12(r)来分析不同物种间的空间关联性。计算公式为:
式(3)中:当g12(r)=1时,则2个不同径级个体之间在r尺度上表现为无关联性;g12(r)>1时,2个不同径级个体之间在r尺度上表现为正相关性;g12(r)<1时,则2个不同径级个体之间在r尺度上表现为负相关性。在本研究中,2个不同径级个体被定义为华北落叶松大树和天然更新幼苗。
分析过程采用完全空间随机模型(CSR)和异质泊松模型(HP)[20],前者假设种群的空间分布不受任何因素的影响,用来检验生境异质性是否影响种群的空间分布;后者考虑到生境异质性对种群的影响,用来分析物种与生境之间的关系[21]。统计过程采用1 m的步长来计算20 m尺度上物种的空间分布及关联,采用蒙特卡洛随机模拟199次,计算置信区间为99%的上下包迹线,以检验物种分布及相关性是否显著[4]。在双变量分析中,g12(r)在上包迹线以上,表明不同径级物种间呈正相关;g12(r)在下包迹线以下,表明不同径级物种间呈负相关;g12(r)在上、下包迹线之间,表明不同径级物种间无空间相关性。
1.2.4 数据处理 空间结构分析及绘图利用R 4.2.5软件的“spatsat”包[22]完成,使用皮尔逊相关法分析更新幼苗状况与生境因子之间的关系,并利用SPSS 23.0软件进行分析。
2 结果与分析
2.1 华北落叶松种群的空间分布机制
2.1.1 更新幼苗的空间分布格局 基于g(r)函数对12块样地内华北落叶松更新幼苗的分析结果表明(图2),1号样地内华北落叶松更新幼苗在0~2 m尺度上表现为聚集分布,在2~10 m尺度上表现为随机分布; 2号样地内华北落叶松更新幼苗在0~7 m尺度上表现为聚集分布,在7~10 m尺度上表现为随机分布;3号样地内华北落叶松天然更新幼苗在0~2.5和7~10 m尺度上表现为聚集分布,在4.0~6.5 m尺度上表现为均匀分布,在其余尺度上表现为随机分布;4号样地内华北落叶松更新幼苗在0~5和8.5~10.0 m尺度上表现为聚集分布,在其余尺度上表现为随机分布;5号样地内华北落叶松天然更新幼苗在0~2.5 m尺度上表现为聚集分布,在4.5~7.0和8~10 m尺度上表现为均匀分布,在其余尺度上表现为随机分布;6号样地内华北落叶松天然更新幼苗在0~5 m尺度上表现为聚集分布,在5.0~5.5 m尺度上表现为随机分布,在5.5~10.0 m尺度上表现为均匀分布;7号样地内更新幼苗在0~5和6~9 m尺度上表现为聚集分布,在其余尺度上表现为随机分布;8号样地内更新幼苗在0~6.5 m尺度上表现为聚集分布,在6.5~9.0 m尺度上表现为随机分布,在9~10 m尺度上表现为均匀分布;9号样地内更新幼苗在0~2.5、4.5~5.0和7~8 m尺度上表现为聚集分布,在8.5~9.0 m尺度上表现为均匀分布,在其余尺度上表现为随机分布;10号样地内更新幼苗在0~4 m尺度上表现为聚集分布,在6~10 m尺度上表现为均匀分布,在4~6 m尺度上表现为随机分布;11号样地内更新幼苗在0~3 m尺度上表现为聚集分布,在6.5~8.0 m尺度上表现为均匀分布,在其余尺度上表现为随机分布;12号样地内更新幼苗在0~3和5.0~6.5 m尺度上表现为聚集分布,在其余尺度上表现为随机分布。
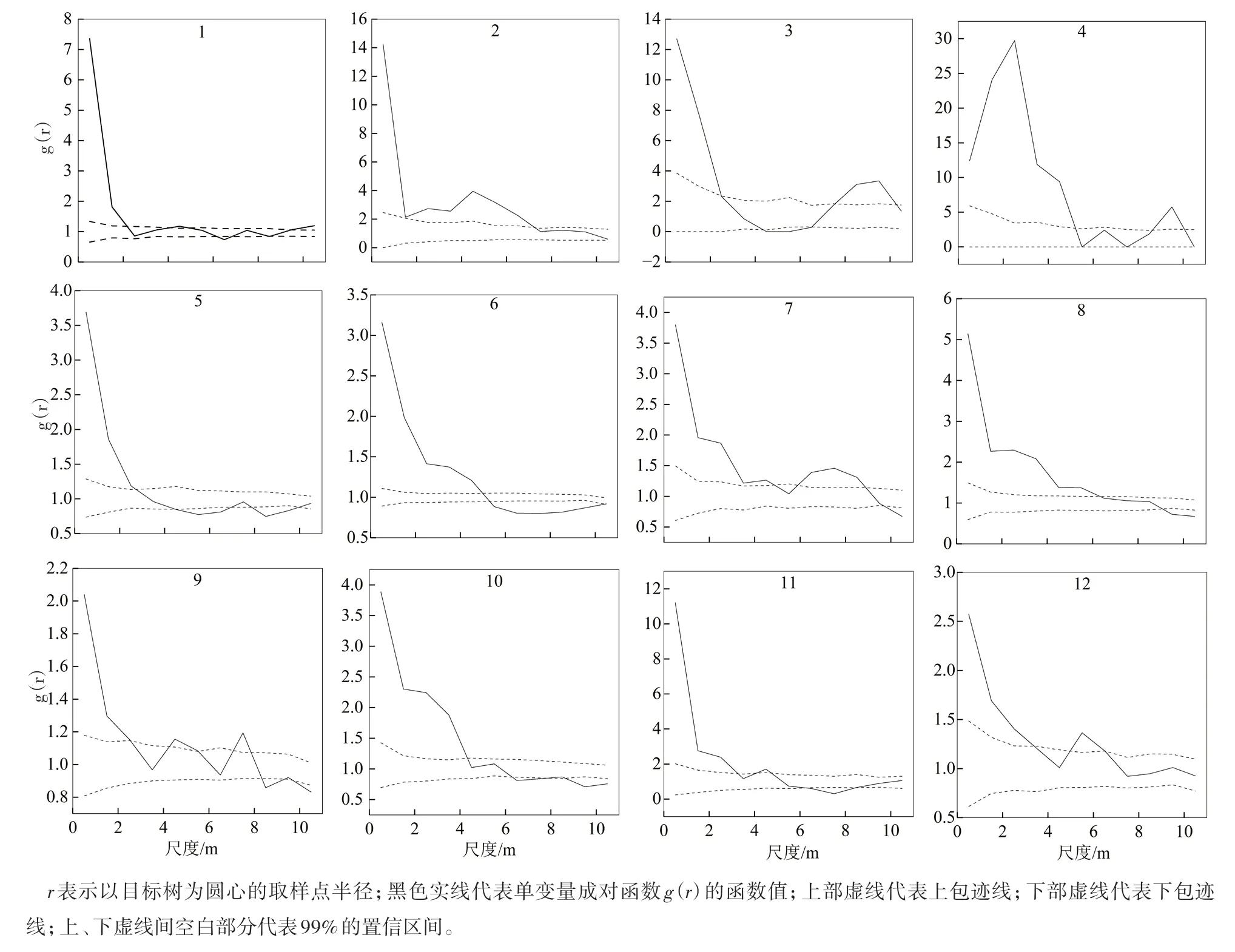
图2 研究区样地内华北落叶松更新幼苗个体间的空间格局
综上,12块样地内华北落叶松天然更新幼苗在小尺度0~2 m范围内表现为聚集分布,在较大尺度(2~10 m)上表现了不同程度的随机和均匀分布。
2.1.2 更新幼苗与大树的空间关联性 对样地内华北落叶松大树和天然更新幼苗之间的空间关联性分析研究结果如图3所示。由图3可知,其1、6、9、10号样地内华北落叶松大树和更新幼苗在整个尺度上无显著空间关联性;2号样地内华北落叶松大树与更新幼苗在0~1、2~4和7~8 m尺度上呈正相关,在其余尺度上无相关性;3号样地在5~7和8~9 m尺度上呈负相关,在其余尺度上无相关性;4号样地在4~6 m尺度上为正相关性,在7.5~9.0 m尺度上呈负关联性,在其余尺度上无相关性;5号样地内大树与更新幼苗在2.5~3.0 m尺度上呈正关联,在其余尺度上表现为无关联性;7号样地内大树与更新幼苗在8~9 m尺度上呈显著正相关,在0~8和9~10 m尺度上两者之间无相互作用;8号样地内大树与更新幼苗在0~1、3~4和5~6 m尺度上表现为正相关性,在1~3、4~5和6~10 m尺度上无相关性;11号样地内大树与天然更新幼苗在4.5~5.0 m尺度上表现为正关联性,在其余尺度上表现为无关联性;12号样地内大树与更新幼苗在2~3 m尺度上表现为正相关性,在其余尺度上无相关性。
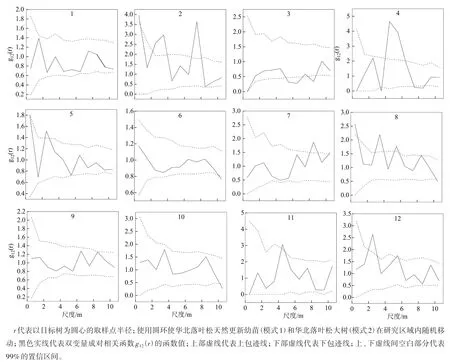
图3 研究区样地内华北落叶松更新幼苗与大树个体间的空间关联性
综上,在大部分的样地内,华北落叶松大树对更新幼苗在研究尺度的小尺度范围(0~4 m)内表现为促进作用,在一定程度上表明,两者的存在有利于对有限资源的充分利用,进而维持稳定共生状况。但在其余样地内,华北落叶松大树和更新幼苗之间无相互促进作用,也无相互抑制作用,即两者生长互不影响,这有利于种群的稳定发展。但在部分样地内,华北落叶松大树和更新幼苗在大尺度上表现为促进作用,表明两者在样地大尺度上能够相互促进生长。
2.2 幼苗生长状况与生境因子的相关性
由表2可知,更新幼苗密度与草本盖度和灌木覆盖度呈负相关,与林龄、郁闭度和枯落物厚度呈正相关;更新幼苗高度与郁闭度呈显著负相关,与林龄、灌木覆盖度和草本盖度呈正相关,与枯落物厚度呈负相关;更新幼苗地径与郁闭度和枯落物厚度呈负相关,与林龄、灌木覆盖度和草本盖度呈正相关。

表2 更新幼苗密度、高度和地径与生境因子的相关性
3 讨论
幼苗期是森林天然更新过程中最关键的时期,对相同种群内物种分布格局及空间关联性的研究有利于深入了解内部的生态学特性及发展进程。在空间分布格局研究中,物种在小尺度上的分布格局主要由苗木自身补给决定,而在大尺度上的分布格局主要由生境异质性决定。
在本研究中,研究样地内均分布有华北落叶松天然更新的幼苗, 在少数样地内华北落叶松天然更新幼苗数量较少。华北落叶松天然更新幼苗在0~2 m空间尺度上表现为聚集分布;随着研究尺度的增加,更新幼苗的聚集性分布格局逐渐变为不同程度的随机或均匀分布,这与Zhao等[18]对落叶松空间分布的研究结果一致。在大尺度和小尺度下的分布格局出现差异性的原因主要是小尺度的聚集分布有利于华北落叶松天然更新幼苗之间的相互遮蔽,以抵御森林中的恶劣环境,缓减土壤养分及水分等的胁迫作用,进而提高成活率;大尺度的随机和均匀分布使华北落叶松天然更新幼苗避免了由生境异质性引起的种内竞争(如对光、养分、水分及其他资源的竞争)[23-24]。通常认为,小尺度的空间分布格局受相邻个体的竞争影响,而大尺度上除了受生境异质性影响外,还由斑块性决定[21]。另一方面,华北落叶松大树数量较少的情况可以为更新幼苗的大量繁殖和生长创造良好的条件,大树受空间环境资源的制约,密度效应导致个体出现死亡,天然更新幼苗由聚集分布向随机或均匀分布转变。
成年大树对其更新幼苗的影响不可忽视,2个物种在不同尺度上表现出不同的关联性,这是由于不同生态机制对物种空间分布的影响有差异[25]。先前的研究证实,成年大树的空间分布特征会影响幼苗和幼树生长的空间相关性[26-27]。当母树呈聚集分布时,其对幼苗的生长表现出明显的抑制作用,并且抑制强度会随着聚集强度的增加而增加[10]。在本研究样地内,华北落叶松大树和天然更新幼苗之间在较小的研究尺度上表现出较强的正相关关系,但在一部分样地内华北落叶松大树与天然更新幼苗之间在小尺度上未表现为正相关或负相关关系。这可能由于华北落叶松大树对光照、水分、土壤养分及其他资源的需求较低,不会与幼苗发生竞争。结合样地物种分布特征可以看出,天然更新幼苗大多分布在华北落叶松大树周围,但在一些样地内,华北落叶松大树周围出现天然更新幼苗的频数较少,这可能是由于大树阻碍了种子扩散,因华北落叶松的种子具有种翅,其传播主要靠风,大多数分散的种子只能在小范围内形成幼苗。华北落叶松个体间在较大尺度上未表现出空间关联性,可能的原因是物种在大尺度上出现了分离现象,很难使2个个体产生相互作用。Pacala等[28]提出的空间分离假说表明,种群内部的聚集最终会导致种间在大尺度上的分离,也就是说在较大尺度范围上不会出现任何关联性。这与Wang等[29]和Zhang等[30]的研究结果一致。生境因子与天然更新幼苗生长状况的相关关系不明显,此问题有待后期深入探究。
4 结论
(1)基于g(r)函数对分布在样地内的华北落叶松天然更新幼苗个体间的空间分布格局显示分析,华北落叶松天然更新幼苗在0~2 m尺度上表现为聚集分布,在2~10 m尺度上表现为随机分布或均匀分布,这种分布有利于天然更新幼苗充分利用环境资源。
(2)基于双变量g12(r)函数对华北落叶松大树与更新幼苗进行空间关联性分析表明,华北落叶松大树对天然更新幼苗在小尺度上表现为正关联性,在较大尺度上表现为无关联性或负关联性,这种关联性有利于华北落叶松的天然更新进程向好的方向发展。
