有机物料与化肥长期配施对小麦玉米轮作潮土细菌群落和酶活性的影响
2023-10-25张灵菲马垒李玉东郑福丽魏建林谭德水崔秀敏李燕
张灵菲,马垒,李玉东,郑福丽,魏建林,谭德水,崔秀敏,李燕
有机物料与化肥长期配施对小麦玉米轮作潮土细菌群落和酶活性的影响

1山东农业大学资源与环境学院,山东泰安 271018;2山东省农业科学院农业资源与环境研究所,济南 250100
【目的】研究有机物料与化肥长期配施对土壤细菌群落和酶活性的影响,揭示土壤养分、胞外酶活性与细菌群落之间关系,为制定潮土区小麦玉米轮作制度下长期且合理的施肥策略提供理论依据。【方法】连续10年的定位试验,设置不施肥 (NF)、单施化肥(NPK)、化肥配合秸秆还田(NPKS)、50%的化肥配施6 000 kg·hm-2猪粪(NPKP)和50%的化肥配施6 000 kg·hm-2牛粪(NPKC)等5个处理。【结果】(1)有机物料与化肥长期配施(NPKS、NPKP和NPKC)可显著提高土壤肥力和胞外酶活性,其中NPKC处理提升效果最为显著,土壤有机质、全氮、碱解氮、有效磷含量和碱性磷酸酶活性较NPK处理提升幅度为13.8%—15.4%、9.7%—15.5%、7.2%—15.9%、13.6%—38.5%和2.5%—13.1%。(2)有机物料与化肥长期配施显著改变了细菌群落结构和组成。与NPK处理相比,小麦季NPKS处理显著上调物种为和,NPKP处理显著提高、和丰度,NPKC处理仅丰度显著升高。玉米季NPKS处理显著上调的物种为和,NPKP处理显著提高丰度;NPKC处理和丰度显著升高。(3)功能预测结果表明,长期有机物料与化肥配施较单施化肥对土壤碳、氮循环功能有促进作用,尤其NPKC处理对硝化作用、尿素溶解、芳香族化合物的降解、木聚糖分解、纤维素降解作用均有较强的影响。(4)蒙特尔分析表明,土壤pH是潮土细菌群落结构和生态功能的主要调控因素。【结论】有机物料与化肥长期配施(尤其是化肥配施牛粪)可提高土壤肥力和胞外酶活性,增加有益菌群丰度,显著改变细菌群落结构和组成,有利于碳、氮、磷循环,从而促进潮土形成适合作物和细菌生长的环境。
小麦玉米轮作;有机物料;化肥;长期施肥;细菌群落;酶活性;土壤养分;潮土
0 引言
【研究意义】土壤细菌可通过调节碳、氮、磷等胞外酶活性,参与有机质降解和养分运转,对维持农田生态系统功能具有重要作用[1]。然而土壤细菌对其生存环境变化极为敏感[2],受土壤耕作措施的深刻影响。有机无机肥配施可以提升土壤养分,促进团聚体结构形成,是一项绿色可持续农业发展措施。研究有机无机肥配施对细菌群落的影响,揭示细菌与土壤理化性质关系,为黄淮海潮土区土壤地力提升和有机废弃物的综合利用提供理论依据。【前人研究进展】有机无机肥配施对土壤细菌群落影响已有大量研究,大多数研究发现配施有机肥可显著提高细菌群落多样性并改善细菌群落结构。SCHMID 等[3]发现长期有机无机肥配施提高细菌多样性。LI等[4]通过25年定位试验发现,有机无机肥配施可显著提高变形菌门和放线菌门等R-策略型物种(偏好高养分生态位,具有较快的生长速率)丰度。LIU等[5]研究表明,有机无机肥配施通过增加土壤不稳定碳含量的方式,刺激地杆菌和厌氧菌介导土壤氮循环。但也有研究表明,有机无机肥配施增加了假单胞菌属和梭状芽孢杆菌属等病原菌的相对丰度,并且引入了新的病原菌链球菌属[6]。有机无机肥配施对微生物的影响主要是通过改变土壤酸碱度[7]、增加有机物质库和养分的可用性[8],还可以促进有机颗粒与矿物结合形成大团聚体[9],为微生物生长繁殖提供稳定的生存空间。有机无机肥配施对细菌的影响与土壤类型密切相关[10]:敖金成等[11]发现pH是黄壤细菌多样性的主要决定因素;LI等[8]发现潮土中有机无机肥配施主要通过增加有机碳和全氮改变细菌群落组成;而CUI等[12]则发现有机无机配施后,土壤有机碳和碱解氮是影响水稻土形成细菌群落的关键驱动因素。此外,不同有机物种类对细菌群落的影响也存在较大差异。MEI等[13]长达40年对黑钙土研究可知,化肥配施粪肥较配施秸秆显著提高细菌多样性。SUN等[7]则发现畜禽粪便(猪粪、牛粪)较秸秆更有利于细菌群落的稳定性。FAN等[14]则发现相较于猪粪和秸秆,化肥配施牛粪是降低潜在病原菌风险最有效的措施。综上所述,有机无机肥配施对细菌的影响因素多种多样,应结合当地环境条件、有机物料特性选择最佳施肥模式。【本研究切入点】添加有机物料是改善土壤质量,解决秸秆和畜禽粪便等有机废弃物资源化利用问题的有效措施,然而不同有机物料施用下潮土细菌群落的变化情况,及其与土壤养分和酶活性的互作机理尚不明确。【拟解决的关键问题】本研究基于连续10年有机无机肥配施长期定位试验,采用高通量测序技术,测定细菌群落结构和组成,研究其与土壤生化指标互作关系,研究结果将为黄淮海潮土区有机废弃物的资源化利用,制定绿色高效的施肥模式提供理论依据。
1 材料与方法
1.1 长期试验概况
田间试验始于2010年10月,位于山东省德州市德城区哨马营村(116°17′44 E,37°30′34 N)。该地属温带大陆性气候,年均气温13.1 ℃,年均降水556.2 mm,年平均日照时数2 592 h,日照率为60%,无霜期208 d,供试土壤为潮土。试验前土壤养分含量为:pH 8.3,有机质(SOM)20.3 g·kg-1,全氮(TN)1.2 g·kg-1,全磷(TP)0.83 g·kg-1,全钾20.8 g·kg-1,有效磷(AP)37.4 mg·kg-1,速效钾(AK)282 mg·kg-1。冬小麦-夏玉米轮作,小麦品种为济麦22、玉米品种为郑单958。
1.2 试验设计
试验采用随机区组设计,4次重复;每个小区长10 m,宽5 m,总面积为50 m2。设置5个处理:①不施肥(NF);②单施化肥(NPK);③化肥配合秸秆还田(NPKS);④50%化肥配施6 000 kg·hm-2猪粪(NPKP);⑤50%化肥配施6 000 kg·hm-2牛粪(NPKC)。各处理养分投入如表1所示,NPK处理小麦季施用氮肥(N)300 kg·hm-2,磷肥(P2O5)120 kg·hm-2,钾肥(K2O)100 kg·hm-2;玉米季施用氮肥(N)250 kg·hm-2,磷肥(P2O5)45 kg·hm-2,钾肥(K2O)45 kg·hm-2。NPKS处理小麦和玉米秸秆均粉碎后还田,其他处理小麦和玉米秸秆全部移除。NPKP处理小麦季和玉米季均施入6 000 kg·hm-2猪粪(养分含量:N 1.63%、P2O52.51%、K2O 1.01%)、NPKC处理小麦季和玉米季均施入6 000 kg·hm-2牛粪(养分含量N:1.64%、P2O52.58%、K2O 0.96%)。粪肥含水率为30%。小麦季和玉米季所有处理的氮肥基追比均为5﹕5,磷肥、钾肥和有机肥作为底肥在小麦和玉米播种整地前一次性施用。

表1 各处理养分投入情况
除NF和NPKS处理外,其余处理氮磷钾总投入量相同
Except for the NF and NPKS treatments, the total input of nitrogen, phosphorus and potassium in the rest of the treatments is the same
1.3 样品采集与测定
土壤样品采集时间为2021年小麦灌浆期和玉米灌浆期。采用“S”形采样法,每个小区取0—20 cm土层土样7钻,充分混匀。将土样置于含有冰袋的保温箱运回实验室。立即过2 mm筛,一部分置于4 ℃冰箱保存测土壤酶,另一部分置于-80 ℃冰箱保存测微生物,剩余土样风干后测定土壤pH及养分含量。
土壤养分参照《土壤农化分析》[15]测定:pH按水土比5﹕1用pH计测定;SOM采用重铬酸钾容量法(外加热法)测定;土壤TN采用全自动凯氏定氮仪测定;土壤TP采用酸溶-钼锑抗比色法测定;碱解氮(AN)采用碱解扩散法测定;AP采用0.5 mol·L-1碳酸氢钠浸提钼锑抗比色法测定;AK采用醋酸铵浸提火焰光度法测定。土壤酶活性采用北京索莱宝科技有限公司试剂盒测定,β-葡萄糖苷酶(β-GC)采用对硝基酚比色法;脱氢酶(DHA)采用氯化三苯基四氮唑还原法;脲酶(UE)采用靛酚蓝比色法;碱性磷酸酶(ALP)采用磷酸苯二钠比色法。
每个土壤样品称0.50 g鲜土,根据Fast DNA Spin Kit for Soil 试剂盒(MP Biomedicals,Santa Ana,CA,USA)说明书提取DNA。对土壤细菌16SrRNA V4-V5区进行PCR扩增。扩增引物为515F(GTGYCAGC MGCCGCGGTAA)、907R(CCGYCAATTYMTTT RAGTTT);扩增条件为94 ℃预变性5 min,30 个循环(90 ℃变性60 s,55 ℃退火60 s,72 ℃延伸75 s),72 ℃延伸10 min。利用QIA quick PCR Purification Kit(Qiagen)纯化2%琼脂糖凝胶回收的PCR产物。根据荧光定量结果和样品的测序量,将PCR扩增产物按比例混合,采用Illumina MiSeq 进行细菌测序。
1.4 数据处理与分析
土壤细菌高通量测序数据处理步骤:检验序列质量,合并双端序列;筛选高质量序列(质量分数>20),并切除扩增引物;去冗余,聚类(UNOISE算法聚类生成ASV);剔除嵌合体;利用RDP数据库(http://rdp.cme.msu.edu/)对序列进行物种分类注释。
使用R 4.2.0中的“vegan”包进行计算Bray-curtis距离进行主坐标分析(PCoA);使用STAMP 2.1.3 软件进行ASV水平差异物种分析,组间差异显示95%的置信区间,并只显示Welch’s t-test经FDR校正后q<0.05的部分;使用 FAPROTAX 数据库预测土壤细菌生态功能;使用 Mantel 检验计算细菌β-多样性、功能结构与土壤生化指标之间的相关关系;使用Microsoft Excel 2019 、SPSS 25.0、Origin 2022软件进行数据处理和绘图。
2 结果
2.1 不同有机物料与化肥长期配施对土壤养分的影响
连续10年有机无机肥配施显著提高了土壤肥力(表2)。在小麦季,与NPK处理相比,NPKS处理土壤有机质(SOM)和碱解氮(AN)分别提升 9.2%和12.0%(<0.05);NPKP处理AP提升33.8%;NPKC处理显著提高SOM、TN、AN和AP含量,分别为13.8%、15.5%、15.9%和38.5%。在玉米季,与NPK处理相比,NPKS处理SOM显著升高9.5%;NPKP处理SOM、TN、AN和AP含量分别提升9.1%、7.3%、6.5%和26.6%,pH则降低0.08个单位;NPKC处理SOM、TN、AN和AP含量分别提高15.4%、9.7%、7.2%和13.6%。

表2 不同有机物料与化肥长期配施对土壤养分的影响
NF:不施肥,NPK:单施化肥,NPKS:化肥配合秸秆还田,NPKP:50%化肥配施6 000 kg·hm-2猪粪,NPKC:50%化肥配施6 000 kg ·hm-2牛粪;同列数据后小写字母表示处理间差异显著(<0.05)
NF: No fertilization, NPK: Chemical fertilizer, NPKS: Chemical fertilizer with straw return, NPKP: 50% chemical fertilizer with 6 000 kg·hm-2pig manure, NPKC: 50% chemical fertilizer with 6 000 kg·hm-2cow manure; Values followed by different letters in same column indicate significant difference among treatments (<0.05)
2.2 不同有机物料与化肥长期配施对土壤酶活性的影响
长期有机无机肥配施可显著提高土壤酶活性(图1)。在小麦季,与NPK处理相比,NPKS处理β-GC和DHA分别显著提高17.1%和37.2%;NPKP处理提高DHA、UE和ALP活性3.9%、2.0%和1.6%;NPKC处理提高DHA、UE和ALP活性分别为3.9%、14.5%和2.5%。在玉米季,与NPK处理相比,NPKS处理提高β-GC、UE和ALP活性,分别为2.0%、1.0%和12.5%,其中β-GC在该处理中最高值为75.05 μg·h-1;NPKP处理可提高UE和ALP活性0.8%和13.3%;NPKC处理提高β-GC、UE和ALP活性1.2%、5.9%和13.1%。
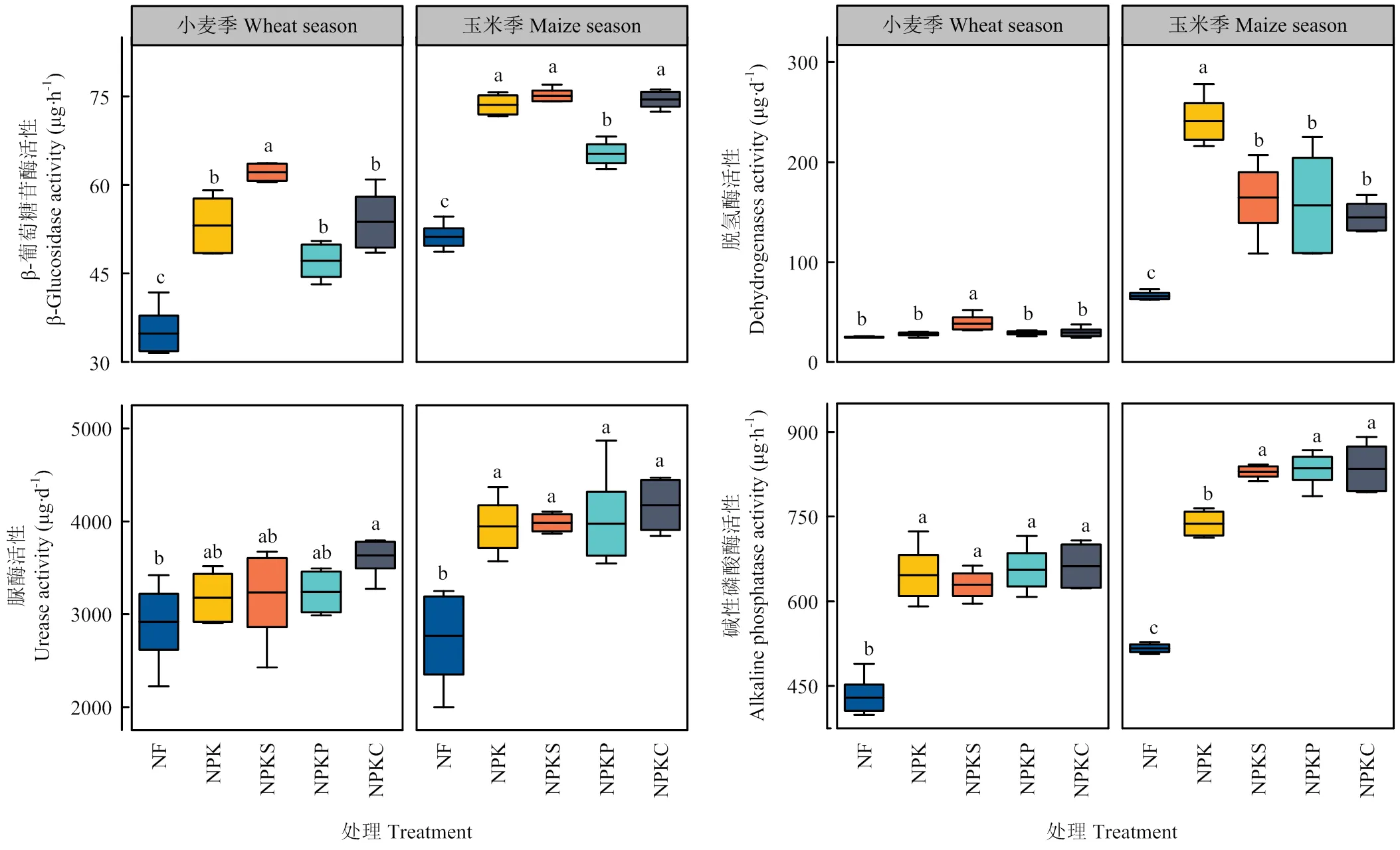
图中箱体代表酶活性范围,箱体中短线代表酶活性均值,箱体上误差棒代表酶活性变异大小,方柱上不同字母表示不同处理间差异显著(P<0.05)
2.3 不同有机物料与化肥长期配施对土壤细菌群落的影响
2.3.1 土壤细菌群落结构 基于ASV水平的Bray- Curtis距离进行主坐标轴分析(图2),结果发现主坐标第一轴解释了28.97%的群落差异,并将小麦季和玉米季细菌群落分开;第二轴解释了5.68%的群落差异,并将不同施肥处理分开。ADONIS多元方差分析结果表明,不同作物生长季(2=0.29,=0.0001)和施肥处理(2=0.12,=0.0050)间细菌群落结构差异均达到极显著水平。
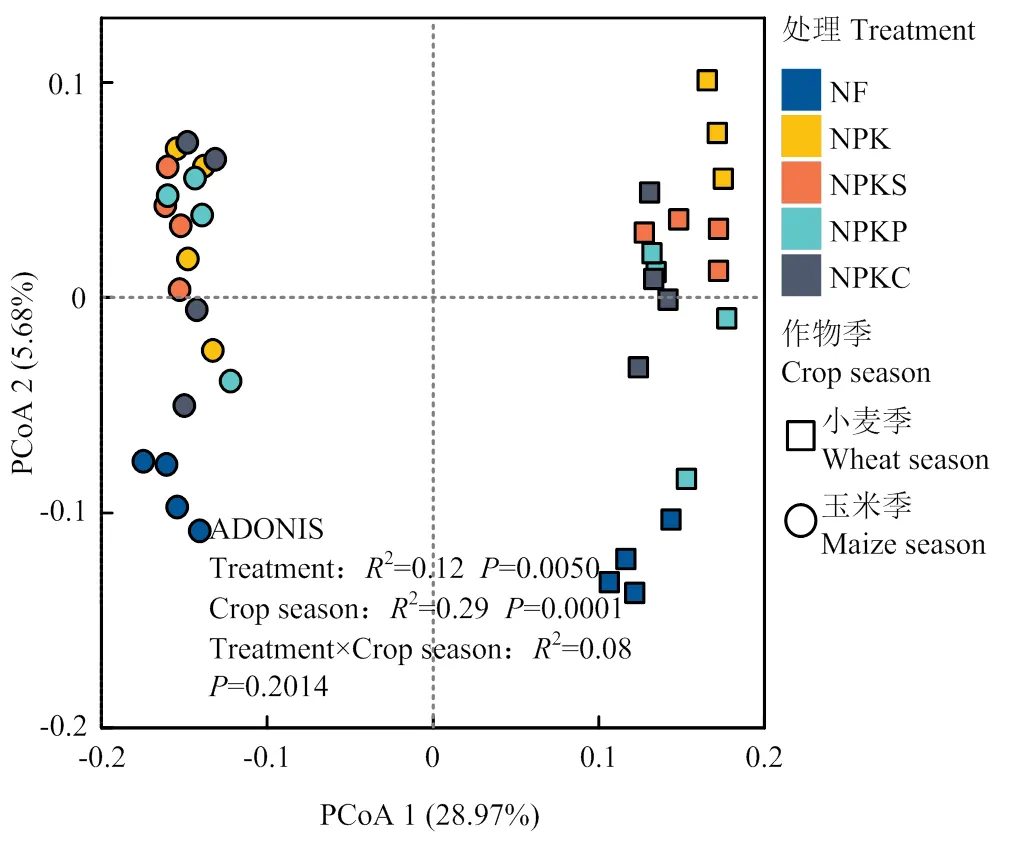
图2 长期不同有机物料与化肥配施对细菌群落结构的影响
2.3.2 土壤细菌群落组成 不同施肥处理土壤细菌群落组成如图3所示。变形菌门(Proteobacteria)是土壤中的优势菌门,其相对丰度在33.5%— 46.6%之间,其余相对丰度大于1%的物种包括酸杆菌门(Acidobacteria)(12.4%—16.6%)、放线菌门(Actinobacteria)(8.4%—11.5%)、浮霉菌门(Planctomycetes)(4.6%—5.4%)、拟杆菌门(Bacteroidetes)(2.8%—6.3%)、绿弯菌门(Chloroflexi)(2.0%—2.8%)和厚壁菌门(Firmicutes)(1.8%—2.4%)。小麦季,与NPK处理相比,有机无机肥配施处理(尤其是NPKP处理)提高了酸杆菌门、厚壁菌门相对丰度,分别为15.6%—23.7%、18.6%—24.9%,但变形菌门、拟杆菌门、绿弯菌门相对丰度则分别降低4.1%—7.8%、13.3%—24.3%和0.8%—22.1%。玉米季,与NPK处理相比,有机无机肥配施处理提高浮霉菌门相对丰度0.7%—10.7%,降低了绿弯菌门相对丰度1.8%—11.3%,其他物种差异未达显著水平。

图3 长期不同有机物料与化肥配施对细菌群落组成的影响
采用Welch’s t-test,在属水平进行差异物种分析,结果如图4所示。在小麦季,与NPK处理相比,NPKS处理显著上调的物种为ASV4672()和ASV7953(),而显著下调的物种为ASV7233()和ASV11505()。NPKP处理显著提高ASV10744()、ASV8921()和ASV6409()物种丰度,显著降低ASV12167()、ASV7233()、ASV11505()和ASV3070()物种丰度。NPKC处理中ASV2758()丰度显著升高,ASV11505()丰度则显著降低。在玉米季,NPKS处理显著上调的物种为ASV5891()和ASV12277(),而显著下调的物种为ASV10358()和ASV7525()。NPKP处理显著提高ASV11154()物种丰度,而显著降低ASV10358()、ASV7462()、ASV6569()和ASV9072()物种丰度。NPKC处理中ASV10805()和ASV5548()物种丰度显著提高,ASV10515()、ASV8136()和ASV9072()物种丰度则显著降低。
2.3.3 土壤细菌功能预测 采用FAPROTAX对细菌群落功能进行预测,共筛选出10个与土壤碳、氮循环相关功能(图5)。小麦季,与NPK处理相比,NPKS处理提升硝酸盐还原作用、硝化作用、反硝化作用和石油烃降解功能;NPKP处理提高硝酸盐还原作用、反硝化作用、木聚糖分解和石油烃降解功能;NPKC处理则提升硝酸盐还原作用、硝化作用、反硝化作用和石油烃降解功能。玉米季时,与NPK处理相比,NPKS处理提升硝酸盐还原作用、固氮作用、硝化作用、反硝化作用、纤维素降解作用和石油烃降解功能;NPKP处理提高硝化作用、尿素溶解、反硝化作用、芳香族化合物降解、纤维素降解作用和石油烃降解功能;NPKC处理则提升固氮作用、尿素溶解、反硝化作用、芳香族化合物降解、纤维素降解作用和石油烃降解功能。

图4 不同有机物料与化肥长期配施差异物种分析
2.4 土壤细菌群落、酶活性与土壤养分相关性分析
采用相关性分析和蒙特尔分析研究土壤养分、酶活性、细菌群落结构和功能之间的互作关系(图6)。结果表明,β-GC与pH(=0.47)、SOM(=0.42)、TN(=0.39)、AN(=0.53)呈显著或极显著正相关,DHA与pH(=0.66)、AN(=0.42)呈极显著正相关,UE与SOM(=0.45)、TN(=0.47)、TP(=0.46)、AN(=0.55)呈极显著正相关,ALP与SOM(=0.54)、TN(=0.57)、TP(=0.54)、AN(=0.67)呈极显著正相关。细菌群落结构与pH(=0.64)、AP(=0.56)、DHA(=0.46)、AK(=0.39)、β-GC(=0.37)、ALP(=0.30)呈极显著正相关(=0.001);细菌生态功能与pH(=0.62)、DHA(=0.52)、AP(=0.47)、β-GC(=0.41)、AK(=0.34)、ALP(=0.32)、UE(=0.22)呈极显著正相关(=0.001)。

图5 基于FAPROTAX预测长期不同有机物料与化肥配施下细菌生态功能
3 讨论
3.1 有机物料与化肥长期配施下土壤养分和酶活性
本研究发现,长期施肥降低pH促使土壤酸碱度偏向中性,这与前人研究结果一致[16]。尽管施肥均导致pH下降,但出现这一现象的原因却存在差异。单施化肥引起pH下降可能由于长期施用化肥发生氮的硝化作用产生H+降低pH[17];而有机无机肥配施则可能由于有机物料(秸秆和有机肥)在分解过程中产生有机酸降低pH[18-19]。有机质是土壤肥力的基础与核心,对农田系统的长期生产力至关重要[20]。刘国辉等[21]利用meta分析发现,施入有机肥后土壤有机质含量提高33.4%。本研究同样发现有机无机肥配施土壤有机质增幅为7.5%—15.4%,其中化肥配施牛粪增幅高达13.8%—15.4%;说明长期有机无机肥配施(尤其是牛粪)有助于提高土壤有机质含量。值得注意的是,与NPK处理相比,化肥减量50%的NPKP和NPKC处理的土壤全氮、全磷、碱解氮和有效磷等养分含量显著升高,这说明粪肥替代50%化肥可提高土壤养分含量及有效性。与NPKS处理相比,NPKC处理显著提高全氮、碱解氮含量,而NPKP则显著提高磷含量。这可能是粪肥为土壤提供大量有机氮和有机磷,增加土壤氮、磷含量,且粪肥较秸秆具有较低C/N和C/P,有机质在降解过程中会释放氮、磷养分,从而提高土壤养分的有效性。
土壤酶参与土壤物质循环和能量流动,是衡量土壤生态功能的重要指标[22]。前人大量研究表明添加有机物料可提高土壤酶活性,但不同有机物料效果存在较大差异[23]。本研究发现化肥配施秸秆可显著提升β-葡萄糖苷酶活性,这是由于秸秆含有大量纤维素、半纤维素,可为细菌提供更多酶促反应底物,刺激微生物分泌β-葡萄糖苷酶。而化肥配施粪肥均可提升碱性磷酸酶活性。与我们研究结果一致,HU等[24]在10年的田间试验研究同样表明,碱性磷酸酶活性和基因在化肥配施粪肥时显著高于配施秸秆处理。一方面,可能由于粪肥较秸秆含有更多易分解有机组分[25],激发细菌分泌酶执行土壤养分循环功能;另一方面,可能由于长期添加外源粪肥增加细菌多样性和数量,促进酶分泌。

用Spearman相关系数和方块大小表示土壤养分与酶活性之间的相关性。***:P<0.001,**:P<0.01,*:P<0.05。方块由小到大表示相关性由低到高。SOM:土壤有机质,TN:全氮,TP:全磷,AN:碱解氮,AP:速效磷,AK:有效钾。β-GC:β-葡萄糖苷酶,DHA:脱氢酶,UE:脲酶, ALP:碱性磷酸酶
3.2 有机物料与化肥长期配施下细菌群落
与前人研究结果一致,变形菌门、酸杆菌门、放线菌门是潮土中的优势菌门[8]。本研究中,与单施化肥相比,添加有机物料增加了酸杆菌门、厚壁菌门丰度,而降低了绿弯菌门丰度。酸杆菌门可降解根系分泌物和枯枝落叶,稳定土壤有机质[26];厚壁菌门适合在养分均衡的土壤环境中生长[27];绿弯菌门为厌氧菌,是土壤缺氧区多糖的主要降解者[28]。因此,有机无机肥配施下养分均衡的环境更适合酸杆菌门和厚壁菌门的生长。本研究中长期化肥配施秸秆显著富集、、和;其中可通过分泌酶(β-葡萄糖苷酶、碱性磷酸酶、酸性磷酸酶、萘酚-AS-BI-磷酸水解酶等)参与秸秆内大量纤维素的降解及有机磷的矿化[29];可参与有机质降解与释放,亦可氧化脂肪酸维持土壤酸碱平衡[30]。化肥配施猪粪显著提高溶磷菌、等物种丰度,可分泌酶矿化有机磷或分泌有机酸活化磷素[31-32],因此NPKP处理中溶磷菌的富集可能是土壤全磷、有效磷和碱性磷酸酶升高的重要原因。、和物种在化肥配施牛粪处理中显著升高。其中可利用牛粪中的蛋白质作为底物进行化学有机营养生长[33];通过分泌赖氨酸脱羧酶、鸟氨酸脱羧酶等,增强作物抗逆性[34];可将硫酸盐、亚硫酸盐和硫代硫酸盐还原为H2S,维持土壤氧化还原平衡[35]。综上所述,有机无机肥配施较单施化肥显著增加有益菌群丰度,改善细菌群落结构。
有机无机肥配施不仅改变了细菌群落组成,还可调控土壤碳、氮循环功能。本研究发现,有机无机肥配施富集的、、、和物种均可参与有机碳的降解[29-30,36-37]。这主要由于碳对大多数细菌有决定性影响,而秸秆和粪肥含有丰富的有机碳,有助于细菌的生长[38],进而可提升碳循环功能。同时,有机无机肥配施提供了充足的低分子有机氮,为微生物氮素转化提供充足底物,刺激细菌群落进行有机氮矿化、无机氮转化等过程,增加固氮菌等功能。李胜君等[39]亦发现有机无机肥配施较单施化肥可提升参与有机氮矿化、固氮和氮还原等异养功能菌数量。值得注意的是,与秸秆和猪粪相比,化肥配施牛粪对碳、氮循环均有较强的影响作用。这可能是由于化肥配施牛粪通过提高土壤大团聚体比例协调细菌群落执行生态功能,参与牛粪中不稳定物质的降解,加快养分周转[40]。
本研究发现pH是土壤细菌群落结构和生态功能的主要影响因素。一方面,大多数细菌对pH较为敏感且最适生长的pH范围较窄,细微的pH变化可直接影响细菌群落结构,如绿弯菌门与pH呈显著正相关[11],酸杆菌门中某些亚群与pH呈负相关[41]。另一方面,pH还可通过调节养分的可用性间接改变细菌群落结构[42]。pH不仅在调控细菌群落中发挥重要作用,还可明显影响生态功能。这可能由于pH驱动细菌群落发生变化,进而改变生态功能[43]。目前,针对细菌群落与生态功能间的联系已有大量研究。例如,变形菌门中的α-变形菌纲参与碳循环[44];放线菌门中部分群落参与氮循环相关的尿素分解、木质素降解[45]。MUNEER等[46]和DELGADO- BAQUERIZO等[47]亦证明pH是细菌群落结构的主要调控因素,且大多数土壤碳、氮循环受pH影响。由此可见,pH在调控细菌群落结构和生态功能中发挥积极作用。
4 结论
有机物料与化肥长期配施可显著提升土壤肥力。不同有机物料与化肥配施对细菌群落影响不同,其中化肥配合秸秆(NPKS)显著富集、等物种,50%的化肥配施猪粪(NPKP)显著提高溶磷菌、等物种的丰度,50%的化肥配施牛粪(NPKC)处理显著提升、r等物种丰度。pH是土壤细菌群落和功能的最主要调控因子。本试验条件下化肥与有机物料配施(尤其与牛粪配施)有利于土壤形成健康的生态系统,实现可持续利用。
[1] BARDGETT R D, VAN DER PUTTEN W H. Belowground biodiversity and ecosystem functioning. Nature, 2014, 515(7528): 505-511.
[2] BAHRAM M, HILDEBRAND F, FORSLUND S K, ANDERSON J L, SOUDZILOVSKAIA N A, BODEGOM P M, BENGTSSON-PALME J, ANSLAN S, COELHO L P, HAREND H, HUERTA-CEPAS J, MEDEMA M H, MALTZ M R, MUNDRA S, OLSSON P A, PENT M, PÕLME S, SUNAGAWA S, RYBERG M, TEDERSOO L, BORK P. Structure and function of the global topsoil microbiome. Nature, 2018, 560(7717): 233-237.
[3] SCHMID C A O, SCHRÖDER P, ARMBRUSTER M, SCHLOTER M. Organic amendments in a long-term field trial-consequences for the bulk soil bacterial community as revealed by network analysis. Microbial Ecology, 2018, 76(1): 226-239.
[4] LI J, COOPER J M, LIN Z A, LI Y T, YANG X D, ZHAO B Q. Soil microbial community structure and function are significantly affected by long-term organic and mineral fertilization regimes in the North China Plain. Applied Soil Ecology, 2015, 96: 75-87.
[5] LIU D M, ZHANG S R, FEI C, DING X D. Impacts of straw returning and N application on NH4+-N loss, microbially reducible Fe(III) and bacterial community composition in saline-alkaline paddy soils. Applied Soil Ecology, 2021, 168: 104115.
[6] 张雅丽, 郭晓明, 胡慧, 郭暖, 徐小涛, 李建林. 牛粪还田对土壤微生物群落特征的影响. 环境科学, 2023, 44(3): 1792-1800.
ZHANG Y L, GUO X M, HU H, GUO N, XU X T, LI J L. Effects of cow manure application on soil microbial community in farmland. Environmental Science, 2023, 44(3): 1792-1800. (in Chinese)
[7] SUN R B, ZHANG X X, GUO X S, WANG D Z, CHU H Y. Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biology and Biochemistry, 2015, 88: 9-18.
[8] LI F, CHEN L, ZHANG J B, YIN J, HUANG S M. Bacterial community structure after long-term organic and inorganic fertilization reveals important associations between soil nutrients and specific taxa involved in nutrient transformations. Frontiers in Microbiology, 2017, 8: 187.
[9] 赵凤艳, 张勇勇, 张玥琦, 张天实, 张国显, 张慧, 杨丽娟. 有机物料对设施番茄长期连作土壤细菌群落结构的影响. 生态学杂志, 2019, 38(6): 1732-1740.
ZHAO F Y, ZHANG Y Y, ZHANG Y Q, ZHANG T S, ZHANG G X, ZHANG H, YANG L J. Effects of organic amendments on soil bacterial community structure with long-term tomato planting in greenhouse. Chinese Journal of Ecology, 2019, 38(6): 1732-1740. (in Chinese)
[10] XUN W B, HUANG T, ZHAO J, RAN W, WANG B R, SHEN Q R, ZHANG R F. Environmental conditions rather than microbial inoculum composition determine the bacterial composition, microbial biomass and enzymatic activity of reconstructed soil microbial communities. Soil Biology and Biochemistry, 2015, 90: 10-18.
[11] 敖金成, 李博, 阎凯, 李永梅. 连作对云南典型烟区植烟土壤细菌群落多样性的影响. 农业资源与环境学报, 2022, 39(1): 46-54.
AO J C, LI B, YAN K, LI Y M. Effects of continuous cropping on tobacco-planting soil bacterial community diversity in typical tobacco-growing areas of Yunnan Province. Journal of Agricultural Resources and Environment, 2022, 39(1): 46-54. (in Chinese)
[12] CUI X W, ZHANG Y Z, GAO J S, PENG F Y, GAO P. Long-term combined application of manure and chemical fertilizer sustained higher nutrient status and rhizospheric bacterial diversity in reddish paddy soil of Central South China. Scientific Reports, 2018, 8: 16554.
[13] MEI N, ZHANG X Z, WANG X Q, PENG C, GAO H J, ZHU P, GU Y. Effects of 40 years applications of inorganic and organic fertilization on soil bacterial community in a maize agroecosystem in northeast China. European Journal of Agronomy, 2021, 130: 126332.
[14] FAN K K, DELGADO-BAQUERIZO M, GUO X S, WANG D Z, ZHU Y G, CHU H Y. Microbial resistance promotes plant production in a four-decade nutrient fertilization experiment. Soil Biology and Biochemistry, 2020, 141: 107679.
[15] 鲍士旦. 土壤农化分析. 3版. 北京: 中国农业出版社, 2000.
BAO S D. Soil and Agricultural Chemistry Analysis. 3rd ed. Beijing: China Agriculture Press, 2000. (in Chinese)
[16] WEYERS E, STRAWN D G, PEAK D, MOORE A D, BAKER L L, CADE-MENUN B. Phosphorus speciation in calcareous soils following annual dairy manure amendments. Soil Science Society of America Journal, 2016, 80(6): 1531-1542.
[17] GUO J H, LIU X J, ZHANG Y, SHEN J L, HAN W X, ZHANG W F, CHRISTIE P, GOULDING K T, VITOUSEK P M, ZHANG F S. Significant acidification in major Chinese croplands. Science, 2010, 327(5968): 1008-1010.
[18] CHEN X D, OPOKU-KWANOWAA Y, LI J M, WU J G. Application of organic wastes to primary saline-alkali soil in northeast China: Effects on soil available nutrients and salt ions. Communications in Soil Science and Plant Analysis, 2020, 51(9): 1238-1252.
[19] YAN Z J, CHEN S, DARI B, SIHI D, CHEN Q. Phosphorus transformation response to soil properties changes induced by manure application in a calcareous soil. Geoderma, 2018, 322: 163-171.
[20] 温延臣, 李海燕, 袁亮, 徐久凯, 马荣辉, 林治安, 赵秉强. 长期定位施肥对潮土剖面养分分布的影响. 中国农业科学, 2020, 53(21): 4460-4469. doi: 10.3864/j.issn.0578-1752.2020.21.014.
WEN Y C, LI H Y, YUAN L, XU J K, MA R H, LIN Z A, ZHAO B Q. Effect of long-term fertilization on nutrient distribution of fluvo-aquic soil profile. Scientia Agricultura Sinica, 2020, 53(21): 4460-4469. doi: 10.3864/j.issn.0578-1752.2020.21.014.(in Chinese)
[21] 刘国辉, 买文选, 田长彦. 施用有机肥对盐碱土的改良效果:Meta分析. 农业资源与环境学报, 2023, 40(1): 86-96.
LIU G H, MAI W X, TIAN C Y. Effects of organic fertilizer application on the improvement of saline soils: Meta analysis. Journal of Agricultural Resources and Environment, 2023, 40(1): 86-96. (in Chinese)
[22] ZHAO S C, LI K J, ZHOU W, QIU S J, HUANG S W, HE P. Changes in soil microbial community, enzyme activities and organic matter fractions under long-term straw return in north-central China. Agriculture, Ecosystems & Environment, 2016, 216: 82-88.
[23] LIU E K, YAN C R, MEI X R, HE W Q, BING S H, DING L P, LIU Q, LIU S, FAN T L. Long-term effect of chemical fertilizer, straw, and manure on soil chemical and biological properties in northwest China. Geoderma, 2010, 158(3/4): 173-180.
[24] HU Y J, XIA Y H, SUN Q, LIU K P, CHEN X B, GE T D, ZHU B L, ZHU Z K, ZHANG Z H, SU Y R. Effects of long-term fertilization on phoD-harboring bacterial community in Karst soils. The Science of the Total Environment, 2018(628/629): 53-63.
[25] XING L, ZHANG Y M, HU C S, DONG W X, LI X X, LIU X P, ZHANG L J, WEN H D. Effects of long-term nutrient recycling pathways on soil nutrient dynamic and fertility in farmland. Chinese Journal of Eco-Agriculture,2022, 30(6): 937-951.
[26] ZHANG Q Q, ZHAO W W, ZHOU Z Z, HUANG G H, WANG X B, HAN Q, LIU G F. The application of mixed organic and inorganic fertilizers drives soil nutrient and bacterial community changes in teak plantations. Microorganisms, 2022, 10(5): 958.
[27] 杨旭, 刘海林, 黄艳艳, 杨红竹, 贝美容, 林清火. 有机无机复混肥施用量对热带水稻土微生物群落和酶活性的影响. 植物营养与肥料学报, 2021, 27(4): 619-629.
YANG X, LIU H L, HUANG Y Y, YANG H Z, BEI M R, LIN Q H. Effects of application of organic-inorganic compound fertilizers on microbial communities and enzyme activities in tropical paddy soil. Journal of Plant Nutrition and Fertilizers, 2021, 27(4): 619-629. (in Chinese)
[28] LIU Y R, LV Z Z, HOU H Q, LAN X J, JI J H, LIU X M. Long-term effects of combination of organic and inorganic fertilizer on soil properties and microorganisms in a Quaternary Red Clay. PLoS One, 2021, 16(12): e0261387.
[29] FUKUNAGA Y, KURAHASHI M, SAKIYAMA Y, OHUCHI M, YOKOTA A, HARAYAMA S.Gen. nov., sp. nov., isolated from a marine alga, and proposal of Phycisphaeraceae fam. nov., Phycisphaerales ord. nov. and Phycisphaerae classis nov. in the Phylum Planctomycetes. The Journal of General and Applied Microbiology, 2009, 55(4): 267-275.
[30] NGATCHOU DJAO O D, ZHANG X J, LUCAS S, LAPIDUS A, GLAVINA DEL RIO T, NOLAN M, TICE H, CHENG J F, HAN C, TAPIA R, GOODWIN L, PITLUCK S, LIOLIOS K, IVANOVA N, MAVROMATIS K, MIKHAILOVA N, OVCHINNIKOVA G, PATI A, BRAMBILLA E, CHEN A, PALANIAPPAN K, LAND M, HAUSER L, CHANG Y J, JEFFRIES C D, ROHDE M, SIKORSKI J, SPRING S, GÖKER M, DETTER J C, WOYKE T, BRISTOW J, EISEN J A, MARKOWITZ V, HUGENHOLTZ P, KYRPIDES N C, KLENK H P. Complete genome sequence oftype strain (TGB-C1^T). Standards in Genomic Sciences, 2010, 3(3): 267-275.
[31] REZAKHANI L, MOTESHAREZADEH B, TEHRANI M M, ETESAMI H, MIRSEYED HOSSEINI H. Phosphate-solubilizing bacteria and silicon synergistically augment phosphorus (P) uptake by wheat (L.) plant fertilized with soluble or insoluble P source. Ecotoxicology and Environmental Safety, 2019, 173: 504-513.
[32] 田美, 刘汉湖, 申欣, 赵方庆, 陈帅, 姚永佳. 百乐克(BIOLAK)活性污泥宏基因组的生物多样性及功能分析. 环境科学, 2015, 36(5): 1739-1748.
TIAN M, LIU H H, SHEN X, ZHAO F Q, CHEN S, YAO Y J. Biodiversity and function analyses of BIOLAK activated sludge metagenome. Environmental Science, 2015, 36(5): 1739-1748. (in Chinese)
[33] VIEIRA S, LUCKNER M, WANNER G, OVERMANN J.Gen. nov., sp. nov. a new member of subdivision 6 Acidobacteria isolated from temperate grassland soil. International Journal of Systematic and Evolutionary Microbiology, 2017, 67(5): 1408-1414.
[34] SRINIVAS T N R, ANIL KUMAR P, TANK M, SUNIL B, POORNA M, ZAREENA B, SHIVAJI S.sp. nov., isolated from a soil sample of a mud volcano. International Journal of Systematic and Evolutionary Microbiology, 2015, 65(8): 2391-2396.
[35] GRÉGOIRE P, FARDEAU M L, GUASCO S, LAGIÈRE J, CAMBAR J, MICHOTEY V, BONIN P, OLLIVIER B.sp. nov., a thermophilic sulfate-reducing bacterium isolated from a deep terrestrial geothermal spring in France. Antonie Van Leeuwenhoek, 2012, 101(3): 595-602.
[36] CAO Y R, JIN R X, JIANG Y, HE W X, JIANG C L.soli sp. nov., an actinomycete isolated from soil. International Journal of Systematic and Evolutionary Microbiology, 2012, 62(7): 1587-1591.
[37] DE OLIVEIRA PARANHOS A G, PEREIRA A R, DA FONSECA Y A, DE QUEIROZ SILVA S, DE AQUINO S F. Tylosin in anaerobic reactors: Degradation kinetics, effects on methane production and on the microbial community. Biodegradation, 2022, 33(3): 283-300.
[38] LI C X, MA S C, SHAO Y, MA S T, ZHANG L L. Effects of long-term organic fertilization on soil microbiologic characteristics, yield and sustainable production of winter wheat. Journal of Integrative Agriculture, 2018, 17(1): 210-219.
[39] 李胜君, 胡菏, 李刚, 王蕊, 赵建宁, 张贵龙, 修伟明. 化肥减量与有机物料添加对华北潮土微生物氮循环功能基因丰度和氮转化遗传潜力的影响. 环境科学, 2022, 43(10): 4735-4744.
LI S J, HU H, LI G, WANG R, ZHAO J N, ZHANG G L, XIU W M. Impacts of co-application of chemical fertilizer reduction and organic material amendment on fluvo-aquic soil microbial N-cycling functional gene abundances and N-converting genetic potentials in Northern China. Environmental Science, 2022, 43(10): 4735-4744. (in Chinese)
[40] FENG H J, PAN H, LI C L, ZHUGE Y P. Microscale heterogeneity of soil bacterial communities under long-term fertilizations in fluvo- aquic soils. Soil Ecology Letters, 2022, 4(4): 337-347.
[41] ROUSK J, BÅÅTH E, BROOKES P C, LAUBER C L, LOZUPONE C, CAPORASO J G, KNIGHT R, FIERER N. Soil bacterial and fungal communities across a pH gradient in an arable soil. The ISME Journal, 2010, 4(10): 1340-1351.
[42] ZHALNINA K, DIAS R, DE QUADROS P D, DAVIS-RICHARDSON A, CAMARGO F A O, CLARK I M, MCGRATH S P, HIRSCH P R, TRIPLETT E W. Soil pH determines microbial diversity and composition in the park grass experiment. Microbial Ecology, 2015, 69(2): 395-406.
[43] 林惠瑛, 周嘉聪, 曾泉鑫, 孙俊, 李锦隆, 刘苑苑, 谢欢, 吴玥, 张秋芳, 崔琚琰, 程栋梁, 陈岳民. 武夷山黄山松林土壤细菌群落特征沿海拔梯度的分布模式. 生态学杂志, 2022, 41(8): 1482-1492.
LIN H Y, ZHOU J C, ZENG Q X, SUN J, LI J L, LIU Y Y, XIE H, WU Y, ZHANG Q F, CUI J Y, CHENG D L, CHEN Y M. Distribution pattern of soil bacterial community characteristics in aforest along an elevational gradient of Wuyi Mountains. Chinese Journal of Ecology, 2022, 41(8): 1482-1492. (in Chinese)
[44] DE SCALLY S Z, MAKHALANYANE T P, FROSSARD A, HOGG I D, COWAN D A. Antarctic microbial communities are functionally redundant, adapted and resistant to short term temperature perturbations. Soil Biology and Biochemistry, 2016, 103: 160-170.
[45] 刘泽勋, 庄家尧, 刘超, 郑康, 陈玲. 大同铅锌尾矿不同污染程度土壤细菌群落分析及生态功能特征. 环境科学, 2023, 44(7): 4191-4200.
LIU Z X, ZHUANG J Y, LIU C, ZHENG K, CHEN L. Analysis of soil bacterial community structure and ecological function characteristics in different pollution levels of lead-zinc tailings in Datong. Environmental Science, 2023, 44(7): 4191-4200. (in Chinese)
[46] MUNEER M A, HOU W, LI J, HUANG X M, UR REHMAN KAYANI M, CAI Y Y, YANG W H, WU L Q, JI B M, ZHENG C Y. Soil pH: a key edaphic factor regulating distribution and functions of bacterial community along vertical soil profiles in red soil of pomelo orchard. BMC Microbiology, 2022, 22(1): 38.
[47] DELGADO-BAQUERIZO M, ELDRIDGE D J, OCHOA V, GOZALO B, SINGH B K, MAESTRE F T. Soil microbial communities drive the resistance of ecosystem multifunctionality to global change in drylands across the globe. Ecology Letters, 2017, 20(10): 1295-1305.
Effects of Long-Term Synergistic Application of Organic Materials and Chemical Fertilizers on Bacterial Community and Enzyme Activity in Wheat-Maize Rotation Fluvo-Aquic Soil
ZHANG LingFei1, 2, MA Lei2, LI YuDong2, ZHENG FuLi2, WEI JianLin2, TAN DeShui2, CUI XiuMin, LI Yan
1College of Resources and Environment, Shandong Agricultural University, Tai’an 271018, Shandong;2Institute of Agricultural Resources and Environment, Shandong Academy of Agricultural Sciences, Jinan 250100
【Objective】This experiment studied the effects of long-term synergistic application of organic materials and chemical fertilizers on soil bacterial community and enzyme activity, and revealed the relationship between soil nutrients, extracellular enzyme activity and bacterial community, so as to provide a theoretical basis for formulating long-term and reasonable fertilization strategies under wheat-maize rotation system in fluvo-aquic soil. 【Method】Based on a 10-year located experiment, five treatments were set up, including no fertilization (NF), chemical fertilizer (NPK), chemical fertilizer with straw return (NPKS), 50% chemical fertilizer with 6 000 kg·hm-2pig manure (NPKP), and 50% chemical fertilizer with 6 000 kg·hm-2cow manure ( NPKC ). 【Result】(1) The combined application of organic materials and chemical fertilizers ( NPKS, NPKP and NPKC ) could significantly improve soil fertility and extracellular enzyme activity, among which NPKC treatment had the most significant effect. Compared with NPK treatment, the contents of organic matter, total nitrogen, available nitrogen, available phosphorus and alkaline phosphatase activity were increased by 13.8%-15.4%, 9.7%-15.5%, 7.2%-15.9%, 13.6%-38.5%和2.5%-13.1%. (2) Long-term combined application of organic and inorganic fertilizer significantly changed the bacterial community structure and composition. In the wheat season, compared with NPK treatment, NPKS treatment significantly increased the abundance ofand, NPKP treatment significantly increased the abundance of,and, while NPKC treatment only significantly increased the abundance of. In the maize season, compared with NPK treatment, NPKS treatment significantly increased the abundance ofand, NPKP treatment significantly increased the abundance of, and NPKC treatment significantly increased the abundance ofand. (3) The results of functional prediction showed that combined application of organic and inorganic fertilizers could promote soil carbon and nitrogen cycling compared with long-term single application of chemical fertilizer. In particular, the NPKC treatment had a strong effect on nitrification, ureolysis, aromatic compound degradation, xylanolysis and cellulolysis. (4) Mental analysis showed that soil pH was the main factor regulating bacterial community structure and ecological function in fluvo-aquic soil. 【Conclusion】Long-term application of organic and inorganic fertilizers (especially chemical fertilizers combined with cow manure) could improve soil fertility and extracellular enzyme activity, increase the abundance of beneficial bacteria, significantly change the structure and composition of bacterial communities, and promote the circulation of carbon, nitrogen and phosphorus, thus construct an environment suitable for crop and bacterial growth in fluvo-aquic soil.
wheat-maize rotation; organic material; chemical fertilizer; long-term application of fertilizers; bacterial community; enzyme activity; nutrient; fluvo-aquic soil
10.3864/j.issn.0578-1752.2023.19.011
2022-11-03;
2022-12-31
国家重点研发计划(2021YFD1901003)、国家小麦产业技术体系(CARS-03)、山东省农业科学院农业科技创新工程项目(CXGC2022A09)、山东省自然科学基金(ZR2021QD033)
张灵菲,E-mail:zhanglingfei199811@163.com。通信作者崔秀敏,E-mail:xiumincui@sdau.edu.cn。通信作者李燕,E-mail:liyan1008@163.com
(责任编辑 李云霞)
