不同体长南美白对虾摄食发声信号的变化规律
2023-10-19曹正良王秀秀李钊丞沈梦庭胡庆松吴迪
曹正良,王秀秀,李钊丞,沈梦庭,胡庆松,吴迪*
(1.上海海洋大学 海洋科学学院,上海 201306;2.上海海洋大学 工程学院,上海 201306)
对虾是一种具有较好经济效益的水产养殖种类[1]。南美白对虾(Litopenaeusvannamei)因具有抗病性强、生长快和肉质鲜美等优点,已成为对虾主要养殖品种之一[2]。提升养殖产量、降低肉饲比是对虾养殖的重要研究内容。目前,在对虾养殖中,对虾主要依赖饲料托盘观察摄食[3],由于虾塘水体环境浑浊复杂,通过人工很难准确掌握对虾摄食情况,需要使用其他技术手段加以观测。
近年来,被动声学技术逐渐应用到对虾养殖的饲料投喂与生长管理中。Silva等[4]研究表明,南美白对虾摄食时大颚闭合碰撞摩擦会产生声音,摄食前后声音信号特征不同[5-6],因此,可通过对虾摄食声音信号特征判断其摄食及行为状态。被动声学技术作为一种对生物无干扰监测方法,受虾塘水体环境影响较少,可应用于包括对虾在内的水中生物的发声研究[7-9]。目前,基于被动声学技术的自动声学投饵器已应用于实际生产,有效改善了饲料管理能力,提高了对虾的养殖效益[10-12]。研究对虾摄食发声信号特征,判断对虾摄食情况与行为状态[9,13],了解对虾的摄食发声行为规律,有助于构建科学智能的饲养管理体系,提升对虾养殖产量[14-16]。
本研究中,以5种不同体长的南美白对虾为研究对象,采集南美白对虾摄食颗粒饲料与虾壳的声音信息,通过声学信号及单因素方差分析,确定5种体长的南美白对虾摄食颗粒饲料与虾壳时发声信号的变化规律及二者差异特征,以期为精细化对虾养殖及智能投饵机等的应用开发提供数据基础。
1 材料与方法
1.1 材料
试验用虾选取养殖大池(直径3.4 m、高1.2 m)中培育的从幼虾至成虾5个生长阶段(体长2~3、4~5、6~7、8~9、9~10 cm)的南美白对虾,养殖水体温度、盐度、溶解氧和饲料无机盐均维持在适宜范围内,即24 h持续通氧,温度为28 ℃。试验前,将对虾放置在与养殖大池水体环境一致的水箱(44 cm×28 cm×30 cm)中暂养,禁食20 h,确保试验时对虾有足够的摄食欲望。试验时,随机挑选一组对虾(3尾)放入声学试验玻璃水箱(44 cm×28 cm×30 cm),保持与之前一样的水环境,分别投喂粒径为1.2 mm的颗粒饲料及虾壳,一组体长对虾试验结束后,更换另一组体长对虾进行试验。试验玻璃水箱外表面覆盖黑色薄膜,同时在地面垫一层泡沫以减少环境噪声干扰,每个水箱中均配有一个加热棒和增氧管(图1)。
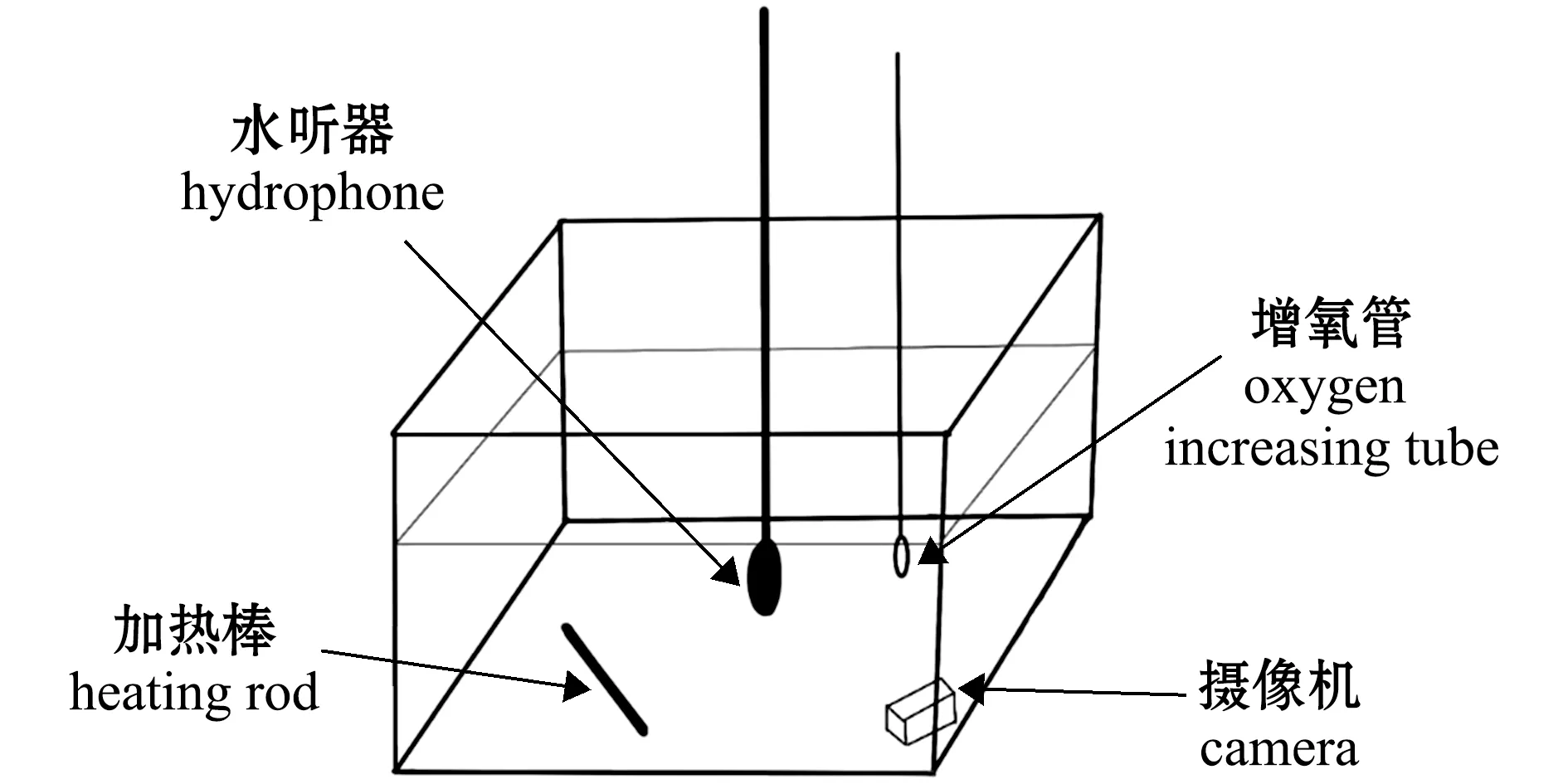
图1 试验装置Fig.1 Equipment of experiment
试验所用装置为大疆水下摄像机和声音采集装置。声音采集装置由带有储存卡的SM4型声音记录仪和1个HTI-96-Min水听器组成(图2)。水听器可接收带宽(2 Hz~30 kHz)声音,可以捕捉到对虾摄食时的声音,SM4型声音记录仪用于储存对虾摄食声音。南美白对虾从幼虾至成虾的最高摄食发声频率为40~48 kHz,因此,本研究中采样频率设置为96 kHz,储存格式为.wav。

图2 声音采集装置Fig.2 Sound acquisition device
1.2 方法
1.2.1 数据采集 采集数据时关闭增氧机,开启与水听器连接的SM4型声音记录仪,将水下摄像机置于相应位置并开启录制。等待约5 min对虾适应环境后,取15颗粒径为1.2 mm的颗粒饲料(或虾壳)润湿后投入水听器下方,记录对虾摄食时间,摄食后8~10 min时结束信号采集,之后进入下一组体长对虾摄食试验。通过对虾摄食饲料与虾壳的音频与视频信息对比,确定对虾摄食信号与行为活动的关系。
1.2.2 发声信号特征分析 使用 Matlab R2019b软件分析对虾摄食颗粒饲料和虾壳的音频信息,挑选有效音频数据对其时间序列信号采用傅里叶变换,分析信号中不同频率成分及信号特征[5]。离散时间傅里叶变换计算公式[25]为
(1)
式中:n为变换长度;ω为角频率;X为时域信号x的傅里叶变换(即频谱)。本文中,信号处理的采样频率为96 kHz,增益为20 dB,进行时频分析时按信号波形总时间的1/50分时窗,并利用Matlab中的Spectrogram 函数进行时频图的绘制。最后确定代表信号特征的各类声学参数。
1.3 数据处理
使用Matlab进行函数定义,将对应对虾摄食声学参数嵌套入单因素方差分析数学模型Analysis of Variance中[26],显著性水平设为0.05,即
MF58是轴向引线玻璃封装型,阻值范围0.1~5000KΩ,体积小、稳定性好、可靠性高,使用温度范围-40~+300℃,热感应快、灵敏度高。
(2)
式中:X为测量值;μ为真值;ε为误差。
由于使用单因素方差分析时,数据需符合正态分布条件,因此,首先对所得到的5种体长对虾摄食颗粒饲料与虾壳发声的最低频率、最高频率、峰值频率及单个脉冲信号时长等4项声学参数值的统计数据进行Lilliefors[27]检验,即
(3)
式中:D*为检验统计量;F^(x)为平均值为0、标准差为1的正态分布累积分布函数;G(x)为经验分布函数值。
2 结果与分析
2.1 摄食声学参数数据的确定
将不同体长对虾摄食的音频数据经过傅里叶变换后得到其摄食的时频、波形和频谱图。对各组9~10 cm的对虾观察采集到的信号发现,投饵前发声信号非常少,投饵后发声信号数量显著增加(图3(a)),可以确定为对虾摄食发声信号。选取其中一个波形信号放大观察(图3(b)),对虾单个摄食发声信号时长约22 ms,结合时频图和频谱图(图3(c)、(d)),确定对虾摄食发声信号的主要频率为5~13 kHz,峰值频率约7 kHz。同时还观察到,对虾摄食期间会产生较高的能量(图3(c)),波形和频率均有一定起伏变化。采用同样的方式,对其余4种体长对虾摄食声学数据进行分析,得到所有体长对虾的摄食声学参数。
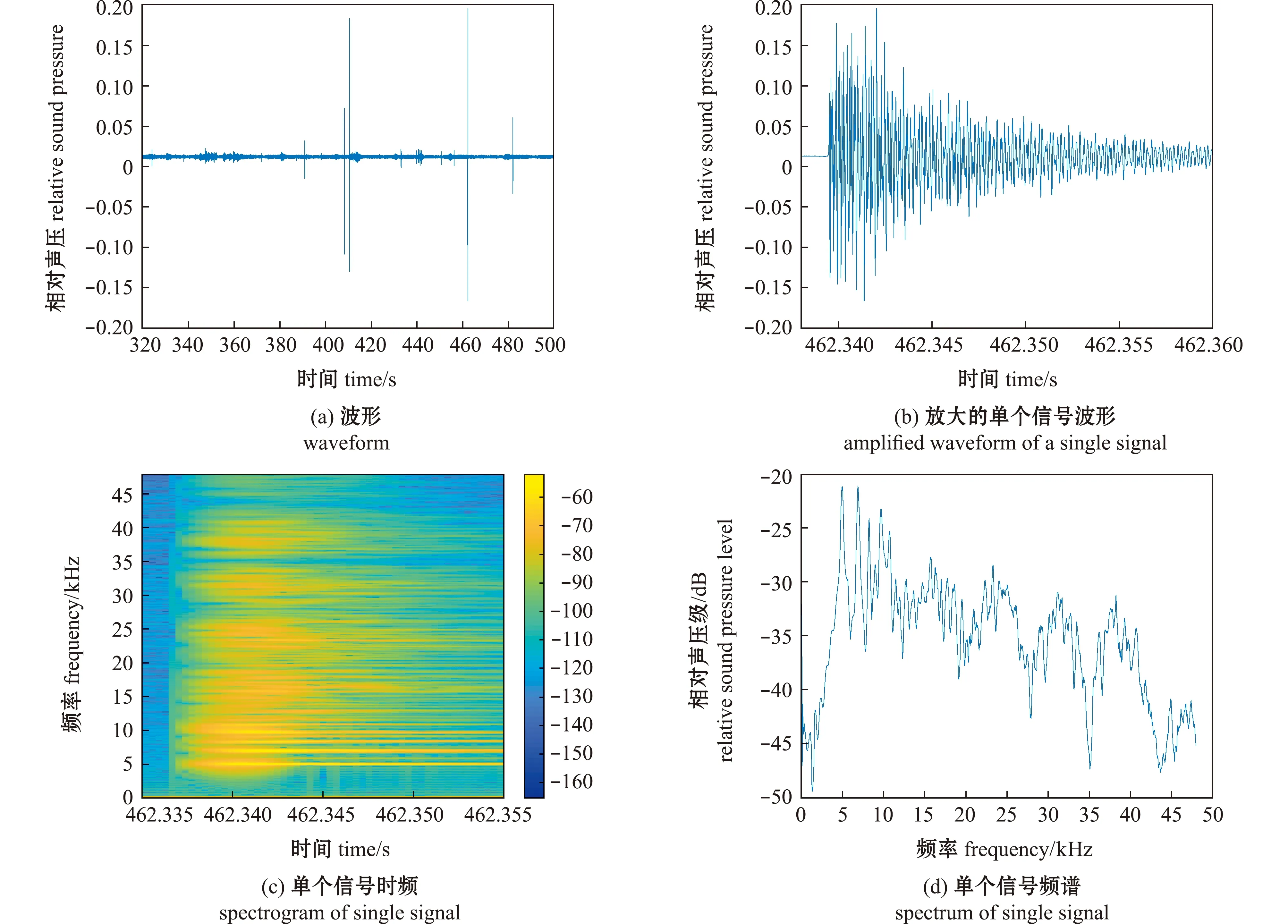
图3 体长9~10 cm对虾的声学信号特征Fig.3 Acoustic signals characteristics of the shrimp with body length of 9-10 cm
2.2 体长与对虾摄食声学参数数据的方差分析
根据Lilliefors检验结果(表1),获取的所有体长对虾的4项声学参数均满足正态分布,因此,可以对试验数据进行单因素方差分析。
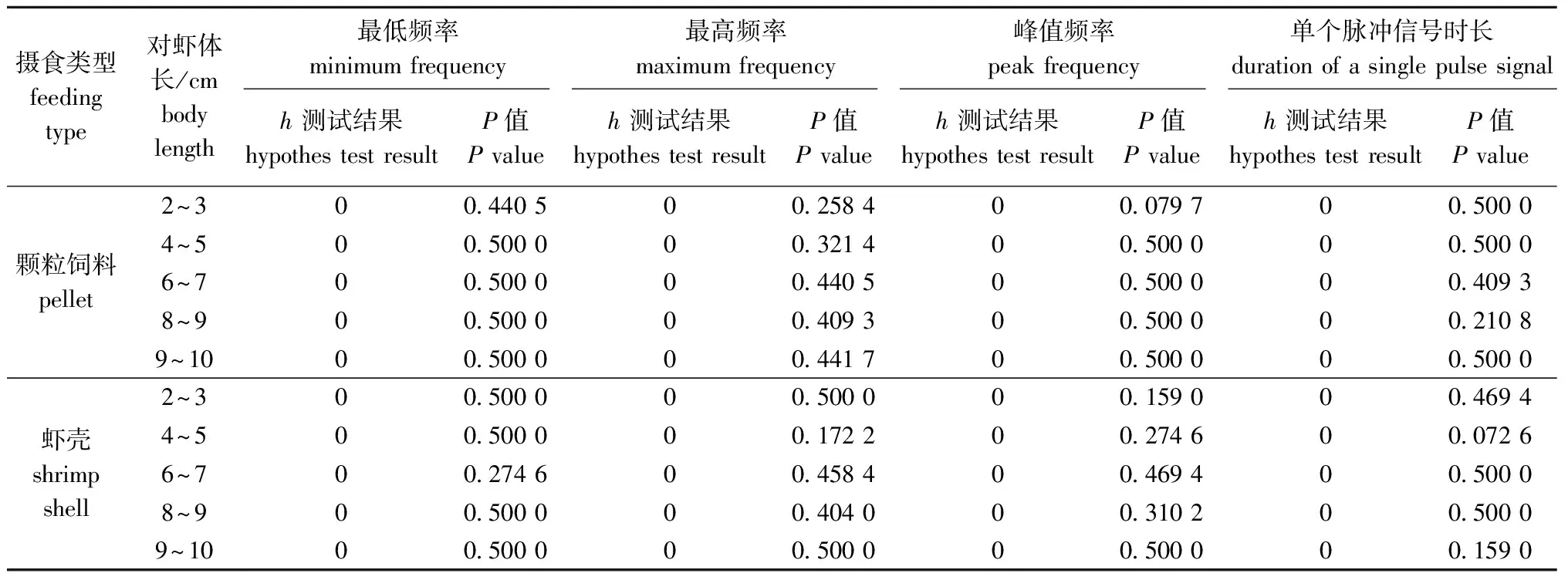
表1 Lilliefors检验Tab.1 Lilliefors test
对5种体长(2~3、4~5、6~7、8~9、9~10 cm)对虾摄食同一粒径颗粒饲料与虾壳的4项声学参数数据进行方差分析显示,对虾体长均极显著影响对虾摄食颗粒饲料与虾壳的4项声学参数(P<0.01)(表2和表3)。
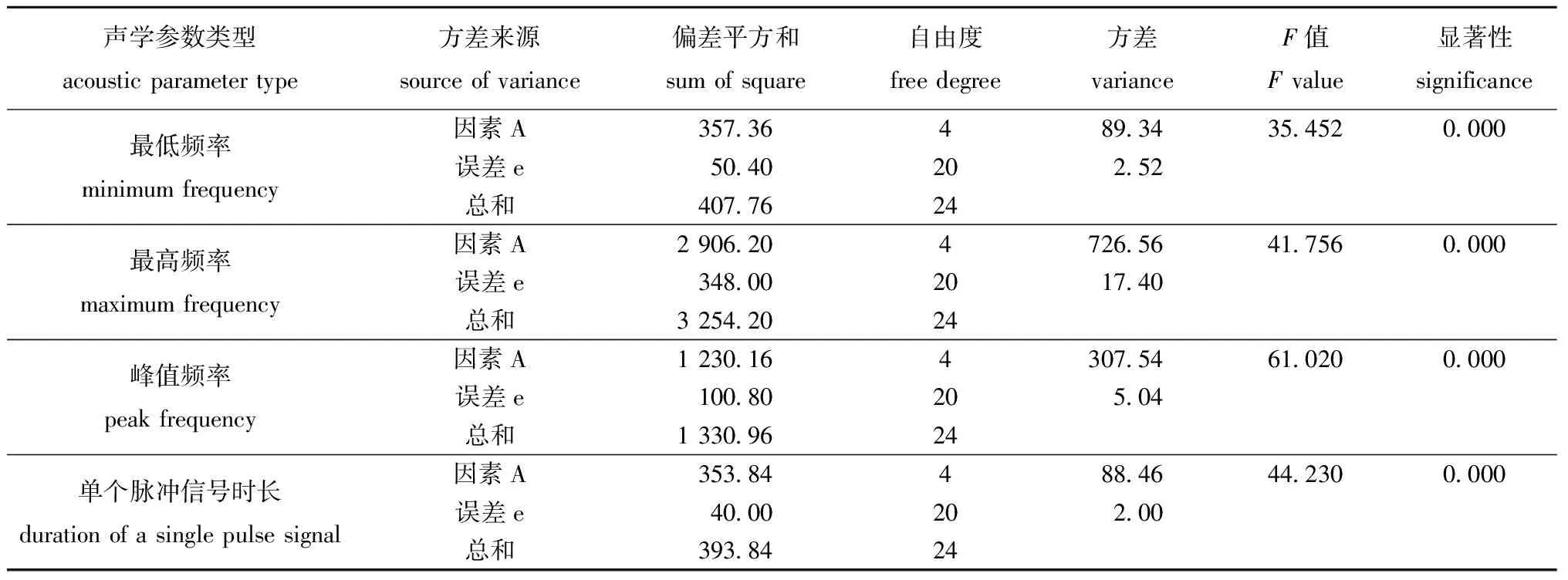
表2 5种体长对虾摄食颗粒饲料声学参数的单因素方差分析Tab.2 Single factor variance analysis of acoustic parameters in the shrimp with 5 body length fed the pellets
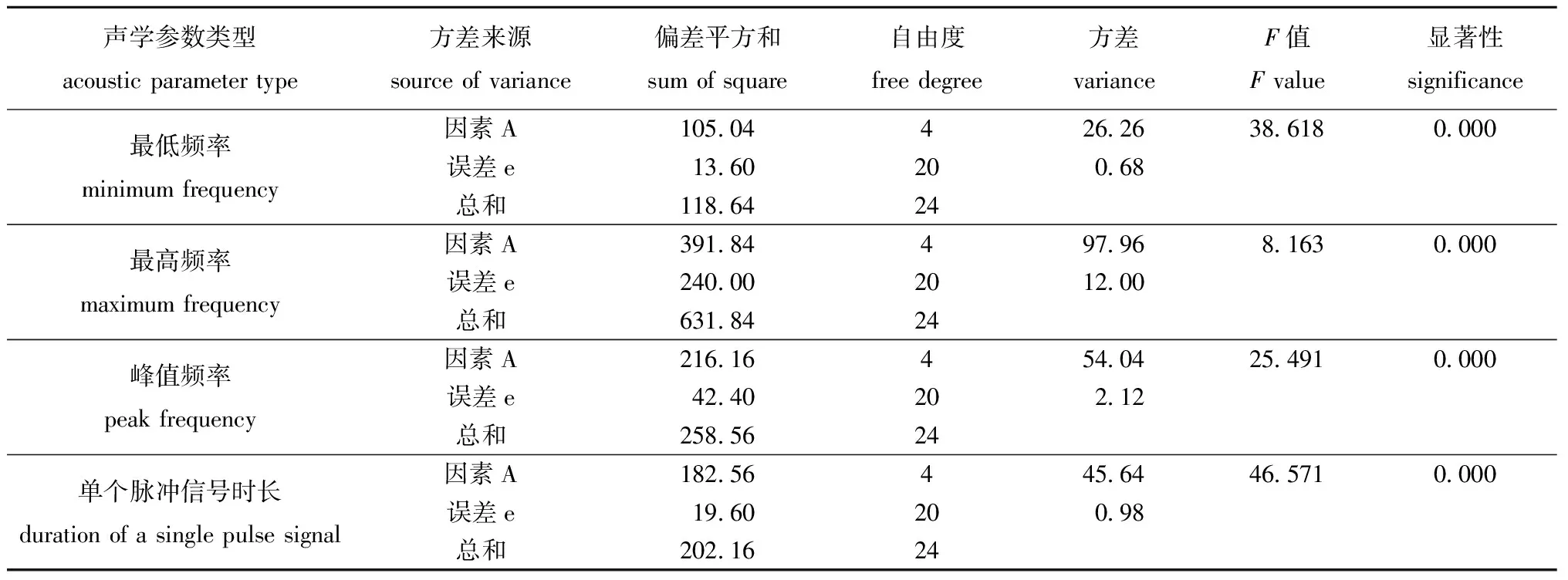
表3 5种体长对虾摄食虾壳声学参数的单因素方差分析Tab.3 Single factor variance analysis of acoustic parameters in the shrimp with 5 body length fed the shrimp shell
2.3 发声频率的变化
2.3.1 峰值频率变化趋势 从图4可见:对虾摄食颗粒饲料的峰值频率随对虾体长的增加而降低(拟合方程为1/y=0.181 1-0.140 65/x,R=0.999 6);对虾体长为2~3 cm时,发声峰值频率最高(约25 kHz),明显高于其他体长;对虾体长为9~10 cm时,发声峰值频率最低(约7 kHz);对虾体长从2~3 cm变化至4~5 cm时,发声峰值频率降低较显著,对虾体长从4~5 cm变化至9~10 cm时,发声峰值频率变化较小,总体呈缓慢降低趋势。
对虾摄食虾壳的发声峰值频率随体长增加无明显变化规律(拟合方程为y=8.856 4 e0.077 682x,R=0.432 2);对虾体长为2~3、8~9、9~10 cm时,发声峰值频率相对较高(分别约13.4、14.8、14.4 kHz);对虾体长为4~5、6~7 cm时,发声峰值频率相对较低(约9 kHz)(图4)。
2.3.2 发声频率范围变化趋势 从图5可见:对虾摄食颗粒饲料的频率范围随对虾体长的增加而缩小;发声最大频率随体长的增加显著降低(拟合方程为y=11.539 2 e1.361 8/x,R=0.99);发声最小频率也随体长的增加而降低(拟合方程为y=3.101 6 e1.412 6/x,R=0.963 7),体长从4~5 cm变化至9~10 cm时,发声最小频率无明显变化;对虾体长为2~3 cm时,发声频率范围最大(14~44 kHz),对虾体长为9~10 cm时,发声频率范围最小(5~15 kHz)。
从图6可见:对虾摄食虾壳的频率范围随对虾体长的增加也无明显变化规律;发声最大频率在对虾体长为6~7 cm时最低(约15 kHz)(拟合方程为y=17.671 8 e0.051 959x,R=0.413 8);发声最小频率随对虾体长的增加而增加(拟合方程为y=4.189 2 e0.150 73x,R=0.800 5);对虾体长为6~7 cm时,发声频率范围最小(5~15 kHz),对虾体长为4~5、8~9 cm时,频率范围最大(5~24 kHz)。
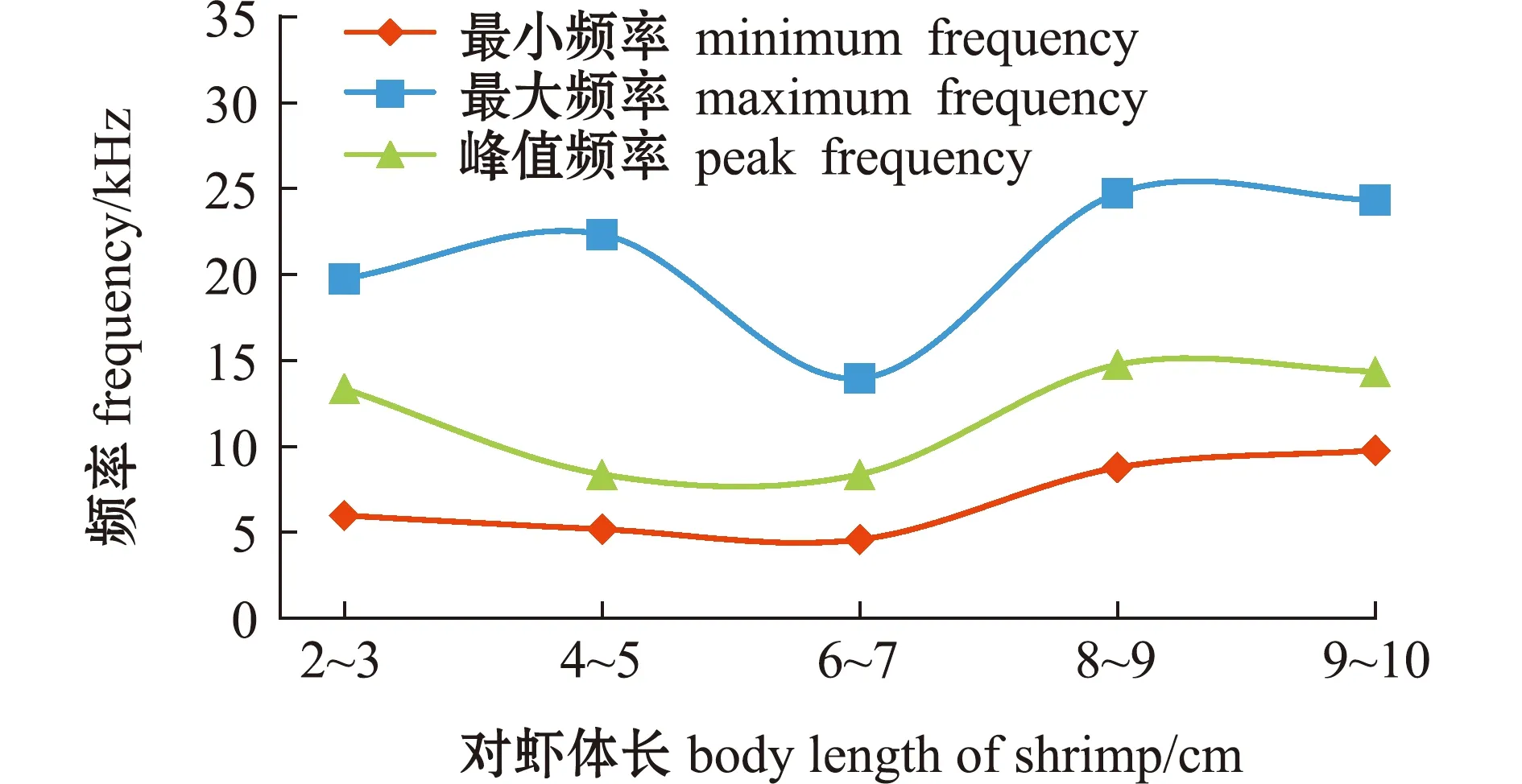
图6 对虾摄食虾壳频率范围与对虾体长的关系Fig.6 Relationship between frequency range of feeding shrimp shell and body length of the shrimp
2.4 单个脉冲信号时长的变化
从图7可见:对虾摄食颗粒饲料的单个脉冲信号时长随体长的增加而增加(拟合方程为y=6.311 1 e0.200 81x,R=0.927 1);对虾体长为9~10 cm时,单个脉冲信号时长最长(约18 ms),明显长于其他体长;对虾体长为2~3 cm时,单个脉冲信号时长最短(约7 ms);对虾体长从4~5变化至8~9 cm时,单个脉冲信号时长无明显变化。
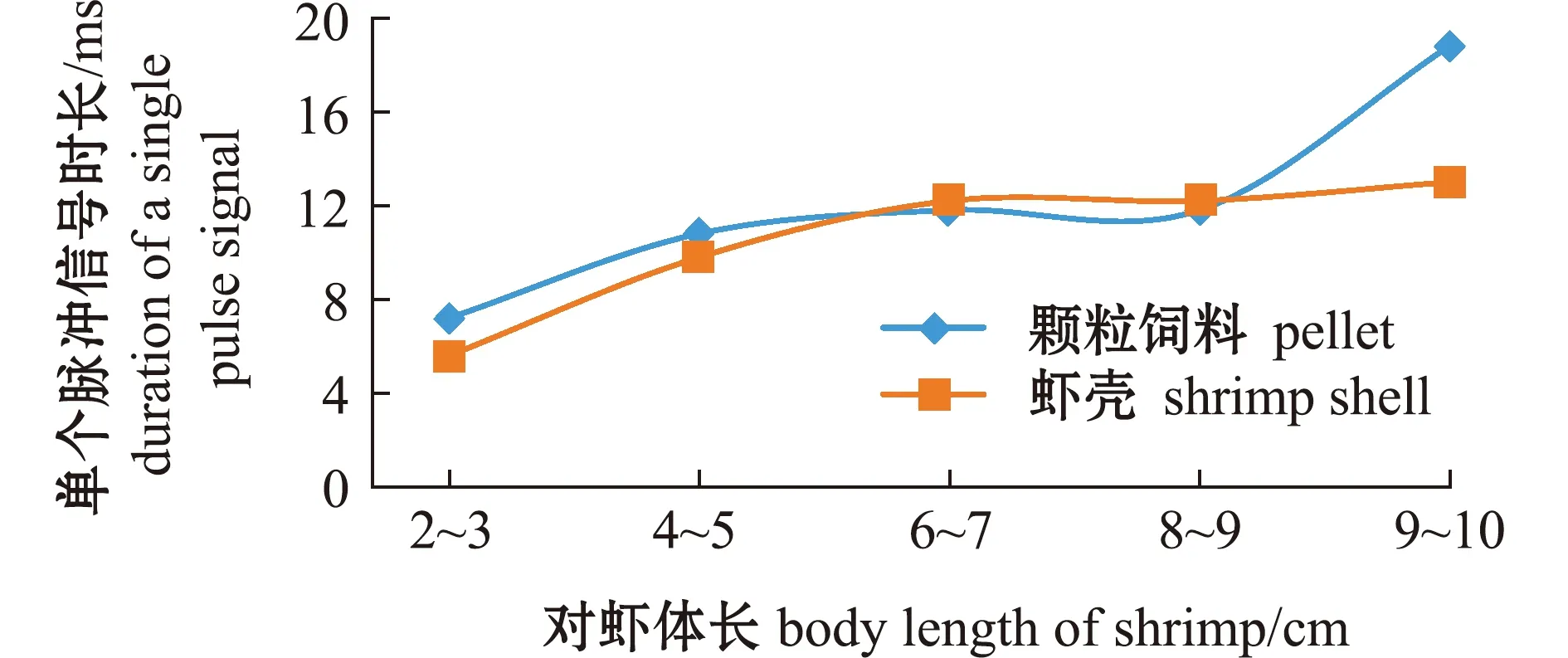
图7 单个脉冲信号时长与对虾体长变化的关系Fig.7 Relationship between duration of a single pulse signal and body length of the shrimp
对虾摄食虾壳的单个脉冲信号时长随体长的增加而增加(拟合方程为y=16.471 3 e-1.066 5/x,R=0.990 1),对虾体长为2~3 cm时,单个脉冲信号时长最短(约6 ms);对虾体长为9~10 cm时,单个脉冲信号时长最长(约13 ms);对虾体长从6~7 cm变化至9~10 cm时,单个脉冲信号时长随体长增加略有增加。
2.5 摄食颗粒饲料与虾壳的声学参数对比
不同体长对虾摄食颗粒饲料与虾壳的声学参数差异明显。峰值频率方面,对虾体长为2~3 cm时,摄食颗粒饲料的峰值频率较摄食虾壳时明显增加,而对虾体长为8~9、9~10 cm时,摄食颗粒饲料的峰值频率较摄食虾壳时明显降低,对虾体长为4~5、6~7 cm时,摄食颗粒饲料与摄食虾壳的峰值频率差异不大(图4)。在频率范围方面,摄食颗粒饲料的频率范围随对虾体长的增加呈明显缩小的趋势,摄食虾壳的频率范围随对虾体长的增加无明显变化规律(图5、图6)。单个脉冲信号方面,对虾摄食颗粒饲料与虾壳的信号时长均随体长增加而增加,区别在于小体长对虾(2~3、4~5 cm)与大体长对虾(9~10 cm)摄食颗粒饲料的单个脉冲信号时长明显长于摄食虾壳的信号时长(图7)。
3 讨论
3.1 对虾摄食颗粒饲料的发声特点
对虾摄食的咀嚼机制导致了发声信号特征的不同,不同体长对虾大颚咀嚼的咬合力不同,从而影响其摄食过程中产生信号的声学参数[21]。小体长对虾大颚发育不成熟导致咬合力弱,需要增加单位时间咬合次数以促进颗粒饲料的物理分解,故出现摄食发声频率高而单次摄食发声时间短的特点;随着对虾生长,大颚发育逐渐成熟,咬合力逐渐增强,单位时间内颗粒饲料的物理分解速率提高,因此,咬合次数不断减少,发声频率降低,而单次咬合时间则增加。本研究中,5种不同体长对虾摄食颗粒饲料发声信号的峰值频率和频率范围均随对虾体长的增加而降低,单个脉冲信号时长随对虾体长的增加而增加。Soares等[22]研究表明,大、小2种体长的对虾在平均声能上存在显著差异。Silva等[4]研究表明,3种体长(9~10、11~12、13~14 cm)对虾摄食发声信号的4项声学参数特征无显著性差异。由于其试验对虾体长均较大,未包含对虾从小到大的快速生长时期,不足以概括出对虾摄食声学信号特征随体长变化的规律。Reis等[28]综述了南美白对虾摄食发声信号在养殖中的应用情况,其中归纳了多篇论文中不同体质量(0.04~0.5、10.2、20.5、31.4 g)的对虾摄食声学信号特征,表明不同体质量对虾摄食颗粒饲料发声信号的峰值频率、最低频率和最高频率有差别,即体质量大的对虾发声频率小,体质量小的对虾发声频率大,但并未考察其变化规律。另外,国外研究中所用颗粒饲料与中国的颗粒饲料类型不同,因此,根据中国常用饲料类型及南美白对虾的生长阶段,确定发声信号的变化规律更具有实际应用意义。
此外,不同种类对虾摄食发声信号也有所不同。Berk[29]发现,白滨对虾(Litopenaeussetiferus)摄食发声信号时长约40 ms,是南美白对虾摄食时长(18 ms)的2.7倍;Daniel等[7]发现,斑节对虾(Penaeusmonodon)摄食发声频率达到50 kHz,均高于现有相关南美白对虾摄食发声研究结果。不同种类对虾摄食时发声信号存在明显的不同特征,主要在于对虾种类不同、摄食发声机制存在差异。中国对虾养殖种类较多,在某些地区还会采用混养方式,其他种类对虾的摄食发声信号是否存在规律,能否反映对虾生长状态有待进一步研究。
3.2 对虾摄食虾壳的发声信号利用
本研究中,5种不同体长对虾摄食虾壳发声信号的峰值频率、频率范围与对虾体长相关,但随着体长增加并无明显变化规律,单个脉冲信号时长则随着体长的增加而增加,4项声学参数特征较对虾摄食颗粒饲料时明显不同。曹正良等[20]研究表明,对虾(体长9~10 cm)摄食沙蚕与不同粒径颗粒饲料时的声学特征存在明显差异,摄食不同粒径颗粒饲料的频率为5~45 kHz,峰值频率为10~17 kHz;摄食沙蚕的频率为5~20 kHz,峰值频率为7 kHz。在虾塘对虾养殖过程中,为了给对虾提供更充足的营养,往往在育苗或特殊情况下补充沙蚕等生物饵料,而对虾生长过程也会出现摄食虾壳的行为。本研究中,对虾(9~10 cm)摄食虾壳的频率范围为10~24 kHz,峰值频率约14 kHz。可以发现,南美白对虾摄食颗粒饲料、沙蚕和虾壳时的声学信号特征存在显著性差异。差异原因可能是三者质地结构不同,沙蚕、虾壳较颗粒饲料坚硬且不易被水浸泡变软,会对南美白对虾摄食沙蚕、虾壳时步足的抓取及大颚的咬合产生一定影响。对虾通过蜕壳实现生长,对虾摄食虾壳发声蕴含着对虾蜕壳后的生长状态,通过研究对虾摄食虾壳发声信号特征可以了解对虾蜕壳生长及异常摄食情况,根据对虾的生长状态可及时更改饲料投喂策略。
3.3 实验室与虾塘条件下对虾摄食情况比较
曹正良等[20]研究表明,虾塘中检测出摄食信号大约在投饵后的2 min甚至更长时间,而实验室大约在投饵后的10 s,差异原因可能与对虾识别饲料并到达饲料处所需要的时间有关。本研究中,对虾被放置在空间较小的水箱中,因此,饥饿状态的对虾确定饲料较快,所得到的对虾摄食发声信号频率与虾塘具有较高的相似性,虾塘中对虾摄食发声信号的峰值频率为5~15 kHz[5],与本研究中体长为2~3 cm对虾摄食颗粒饲料的峰值频率不一致,可能与虾塘中对虾体长较大且大小分布不均、存在其他水生生物摄食发声等有关。而本研究中单个脉冲信号时长与虾塘中的结果存在显著差异,可能是受到试验环境的影响,虾塘声音环境复杂,受多种其他水生生物摄食饲料的声音影响,同时试验玻璃水箱存在声音反射等因素。因此,在后续虾塘摄食发声试验中,要更多考虑对虾摄食虾塘残余饲料和其他饲料(鱼、虾等),以及虾塘中其他水生生物及群虾的摄食发声。
4 结论
1)对虾摄食颗粒饲料时,随着对虾体长的增加,摄食发声信号的峰值频率降低,而频率范围和单个脉冲信号时长均增加。这些变化呈现一定的线性变化规律。
2)对虾摄食虾壳时,随着对虾体长的增加,摄食发声信号的峰值频率呈先降低后上升,频率范围呈先增加后降低再增加再降低的波动变化,单个脉冲信号时长则增加。与摄食颗粒饲料相比,这些摄食发声信号无明显的线性变化规律。
3)体长2~3 cm的对虾摄食颗粒饲料时峰值频率(25 kHz)显著高于摄食虾壳时的峰值频率(13 kHz);体长8~9 cm的对虾摄食颗粒饲料的峰值频率(8 kHz)较摄食虾壳(16 kHz)时明显降低;体长9~10 cm的对虾摄食颗粒饲料的单个脉冲信号时长(18 ms)明显长于摄食虾壳的信号时长(13 ms)。说明对虾摄食颗粒饲料与虾壳的发声信号特征明显不同。
