干旱-复水处理对吉林文冠果幼苗生理特性和光合参数的影响
2023-10-19李美玄刘英卉张友民
李 硕, 李美玄, 刘英卉, 张友民
(吉林农业大学林学与草学学院,吉林长春 130000)
干旱胁迫、低温胁迫、高温胁迫、盐碱胁迫、重金属胁迫等是严重影响植物生长的非生物胁迫,而在全球范围内,干旱胁迫的地理分布最广泛,同时也是危害植物生长最严重、限制农作物产量最主要的环境因子[1]。目前,全球干旱越来越严重,2022年第27届联合国气候变化大会上,正式成立了“全球抗击干旱联盟”以应对全球旱灾,2022年我国的平均降水量较常年同期减少了12.3%,雨量为近40年来最少[2]。可见,干旱已经成为影响作物产量、植物生态系统和生物多样性的全球性问题。
植物处在干旱胁迫时,根系活力会下降,其对土壤营养物质的吸收会受阻,且植物体内更多的营养物质会向根系传导,从而影响植株的生长发育[3]。干旱胁迫会导致植物体内尤其是叶片水势下降,影响水分代谢,进而使植物的生理特性和光合作用效率下降。研究表明,干旱胁迫会影响植物体内渗透调节物质含量变化,积累大量的活性氧,破坏细胞的氧化应激水平,使细胞内抗氧化酶和一些代谢酶活性降低,同时还会对叶片内叶绿素含量产生一定的影响,而严重的干旱胁迫会导致植株萎蔫、枯萎甚至死亡[4-5]。当植物经过一段时间的干旱胁迫后,对其进行复水处理,研究认为大多数的植物会利用自身的代谢调节作用进行自我恢复[6]。因此,研究干旱胁迫和旱后复水处理对植物的生理代谢和光合作用的影响,可以解析植物对逆境胁迫的响应机制,一方面可以揭示植物对干旱胁迫的应答机理,另一方面对植物的高效栽培和技术推广有着重要作用。
文冠果(Xanthocerassorbifolium)是我国重要的生态经济树种和园林观赏树种,同时也是特有的木本油料树种,主要分布于我国较干旱、寒冷的北方地区[7]。其中吉林省西南部文冠果的种植面积排在全国前列,是最适合文冠果产业发展的重要地区之一。目前,吉林省受全球气候影响,年均降水量逐渐减少,水分蒸发量逐渐增大,同时随着部分地区人口集中性增长,出现了不合理的资源开发利用现象,导致吉林省沙漠化土地面积逐渐增加,平均每年干旱期也越来越长,这对文冠果苗木的高效种植产生了严重影响[8]。此外,由于吉林地区独特的降雨格局,常常又会出现干旱与强降雨交替现象,使文冠果苗木在受到干旱胁迫的同时还会受到旱后复水的影响。因此,研究干旱胁迫和复水处理对文冠果苗木生长的影响,对深入了解文冠果苗木如何应对干旱-复水胁迫具有重要意义。
因此,本研究通过对一年生文冠果实生苗进行干旱胁迫和复水处理,模拟其从正常水分到干旱再经过复水的生长过程,从根系活力、渗透调节物质含量、氧化应激作用、叶绿素含量、光合作用等方面探究其体内生理特性变化,以期为文冠果实生苗的干旱-复水抗逆性评价提供理论依据。
1 材料与方法
1.1 材料
文冠果种子来源于吉林省白城市洮南市,2021年4月6日在吉林省文冠果科技有限公司进行育苗培养,2022年5月9日,选取株高、长势一致的一年生实生苗,全部移栽到育苗盆中,育苗盆规格为口径23 cm、高度22 cm、底径18 cm,装土5 L,栽培土壤为泥炭土,有机质含量为33.58 g/kg、有效钾含量为86.78 mg/kg、速效钾含量为115.73 mg/kg、全氮含量为1.23 g/kg、碱解氮含量为68.72 mg/kg、有效磷含量为9.03 mg/kg、速效磷含量为26.52 mg/kg,pH值为6.48,土壤含水量32%,每盆1株。试验地为吉林农业大学设施生产基地温室大棚(120.4°E,43.8°N)。
1.2 试验设计
盆栽苗常规管理1个月,期间土壤含水量在60%~75%,文冠果苗木成功定植后,进行干旱胁迫试验。利用TRASE土壤水分自动监测系统每天测量盆栽土壤含水量。设置干旱胁迫等级,含水量大于60%为正常对照组,含水量在50%~60%为轻度干旱,含水量40%~<50%为中度干旱,含水量在30%~<40%为重度干旱[9]。通过对土壤含水量测定预试验,对同一盆栽土壤连续监测含水量,确定盆栽土在第1天16:00时灌水,当有水从底部漏出时测得土壤平均含水量为66%,盆栽苗维持不浇水管理,在第5天同一时间测得土壤平均含水量为55%(轻度干旱),在第7天同一时间测得土壤平均含水量为46%(中度干旱),在第10天同一时间测得土壤平均含水量为37%(重度干旱)。在第10天16:00测完土壤含水量后及时复水处理,复水处理后测得土壤平均含水量为64%,之后每天定时补充水分,使土壤含水量不低于60%,连续复水处理10 d。
因此,本试验设置正常对照组(CK)和连续干旱-复水处理组,每组10盆为1个处理,3个重复。其中CK一直维持土壤含水量不低于60%。干旱-复水处理组分为2个阶段,第1阶段为干旱胁迫阶段,设置轻度干旱组(D5)、中度干旱组(D7)、重度干旱组(D10);第2阶段为复水处理阶段,设置复水处理3 d组(R3)、复水处理5 d组(R5)、复水处理10 d组(R10)。同时在整个试验过程中,确保CK组土壤含水量一直维持在65%~71%。
干旱-复水处理组样品采集节点:在干旱胁迫前1天16:00时,记为0;干旱胁迫的第5天16:00轻度干旱时,记为D5;干旱胁迫的第7天16:00中度干旱时,记为D7;干旱胁迫的第10天16:00重度干旱时,记为D10;D10的16:00样品采集后开始复水处理,复水处理后第3天16:00时采样,记为R3;复水处理后第5天16:00时采样,记为R5;复水处理后第10天16:00时采样,记为R10。CK组一直维持正常水分管理,样品采集节点同干旱-复水处理组。
在每个样品采集节点上测定土壤平均含水量,测得0的含水量为67%,D5含水量为53%,D7含水量为48%,D10含水量为35%,R3含水量为67%,R5含水量为64%,R10含水量为65%,每个节点上的土壤含水量符合试验设计。
1.3 试验方法
1.3.1 根系活力测定 分别在正常水分0时,干旱胁迫的D5、D7、D10时和复水后R3、R5、R10时采集根系,同时在每个时期均采集CK组幼苗根系。将根系清洗干净后用滤纸控干,称取1 g根系,参照 2,3,5-三苯基氯化四氮唑(TTC法)[10]测定根系活力。
1.3.2 叶片渗透物质含量测定 分别在干旱胁迫前0和干旱胁迫后的D5、D7、D10和复水后R3、R5、R10时采集文冠果幼苗的功能性叶片,功能性叶片位于顶部叶心往下5~6 cm处。采集完后的叶片放入干冰保鲜盒中,在实验室用无菌水清洗干净,并用滤纸吸干表面水分,液氮冷冻后放入-80 ℃冰箱保存,备用待测。叶片脯氨酸(proline,简称Pro)、可溶性糖(soluble sugar,简称SS)、可溶性蛋白(soluble protein,简称SP)含量参考张治安等的方法[10]测定。
1.3.3 叶片叶绿素含量测定 按照“1.3.2”节的方法采集功能性叶片,采用叶片打孔器取样,称取 2 g 叶片,加入无水乙醇研磨,参考丙酮提取法,采用分光光度计比色,测定645、663 nm处吸光度,分别计算叶片中叶绿素a、叶绿素b和叶绿素a+b含量[11]。
1.3.4 叶片丙二醛含量和抗氧化酶活性测定 称取0.1 g待测叶片,加入1 mL预冷的磷酸盐缓冲液,在冰上研磨,低温离心,取上清液待测,丙二醛(malondialdehyde,简称MDA)含量和超氧化物歧化酶(superoxide dismutase,简称SOD)、过氧化氢酶(catalase,简称CAT)、过氧化物酶(peroxidase,简称POD)活性测定均采用苏州科铭生物技术有限公司植物叶片测定试剂盒。
1.3.5 叶片光合参数测定 采用美国Li-cor 6400XT便携式光合作用测定系统,分别在干旱胁迫前0和干旱胁迫后的D5、D7、D10以及复水后R3、R5、R10的10:00—12:00,选取光照方向一致的文冠果幼苗顶部叶片,设置光照强度1 200 μmol/(m2·s),分别测定净光合速率[net photosynthetic rate,简称Pn,单位μmol/(m2·s)]、气孔导度[stomatal conductance,简称Gs,单位mol/(m2·s)]、蒸腾速率[transpiration rate,简称Tr,单位mmol/(m2·s)]、胞间CO2浓度(intercellular CO2concentration,简称Ci,单位μmol/mol)。
1.4 数据处理
全部数据均至少测量3次,采用Excel 2020对数据进行处理,SPSS 22对数据进行单因素方差分析和相关性分析,采用Origin 2018作图。
2 结果与分析
2.1 干旱-复水处理对文冠果幼苗根系活力的影响
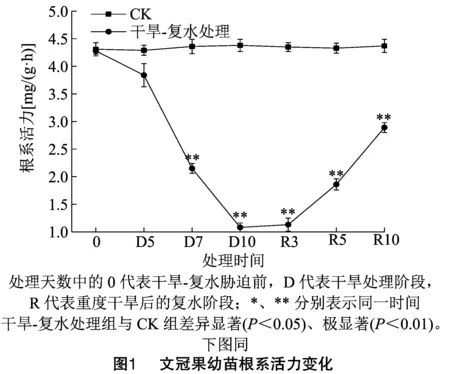
由图1可知,整个试验期间,正常处理组(CK)的文冠果幼苗根系活力一直稳定在4.0~4.5 mg/(g·h),而干旱-复水处理组中,随着干旱胁迫程度的增加,根系活力呈现下降趋势,与同时间节点的CK组相比,D5、D7、D10时的根系活力分别下降了10.48%、50.69%、75.34%,D5时的根系活力与CK组差异不显著,而D7和D10时的根系活力与CK组存在极显著差异,可见,文冠果幼苗具一定的耐轻度干旱作用,而中度干旱和重度干旱则会严重抑制根系活力。复水处理后的R3、R5、R10时,根系活力随着复水天数的增加逐渐升高,此时R3、R5、R10时的根系活力分别为1.13、1.86、2.89 mg/(g·h),与重度干旱时相比,分别提高了4.63%、72.22%、167.60%。R10时的根系活力与CK组相比,仍存在极显著差异,未恢复到干旱胁迫前水平,可见,复水能逐渐缓解干旱对根系活力的抑制,起到一定的补偿作用,但根系活力的恢复需要一定的时间。
2.2 干旱-复水处理对文冠果幼苗叶片渗透调节物质的影响
由图2-A可知,CK组文冠果叶片内Pro含量维持在20~30 μg/g,含量相对稳定;在干旱-复水处理组中,随着干旱胁迫程度的增加,叶片内Pro含量显著增加,与同时期CK组相比,D5、D7、D10分别增加了34.49%、142.28%、313.06%。复水处理后,R3、R5、R10时,Pro含量呈直线下降,在R10时,叶片内Pro含量恢复到了胁迫前的水平,与CK组差异不显著。
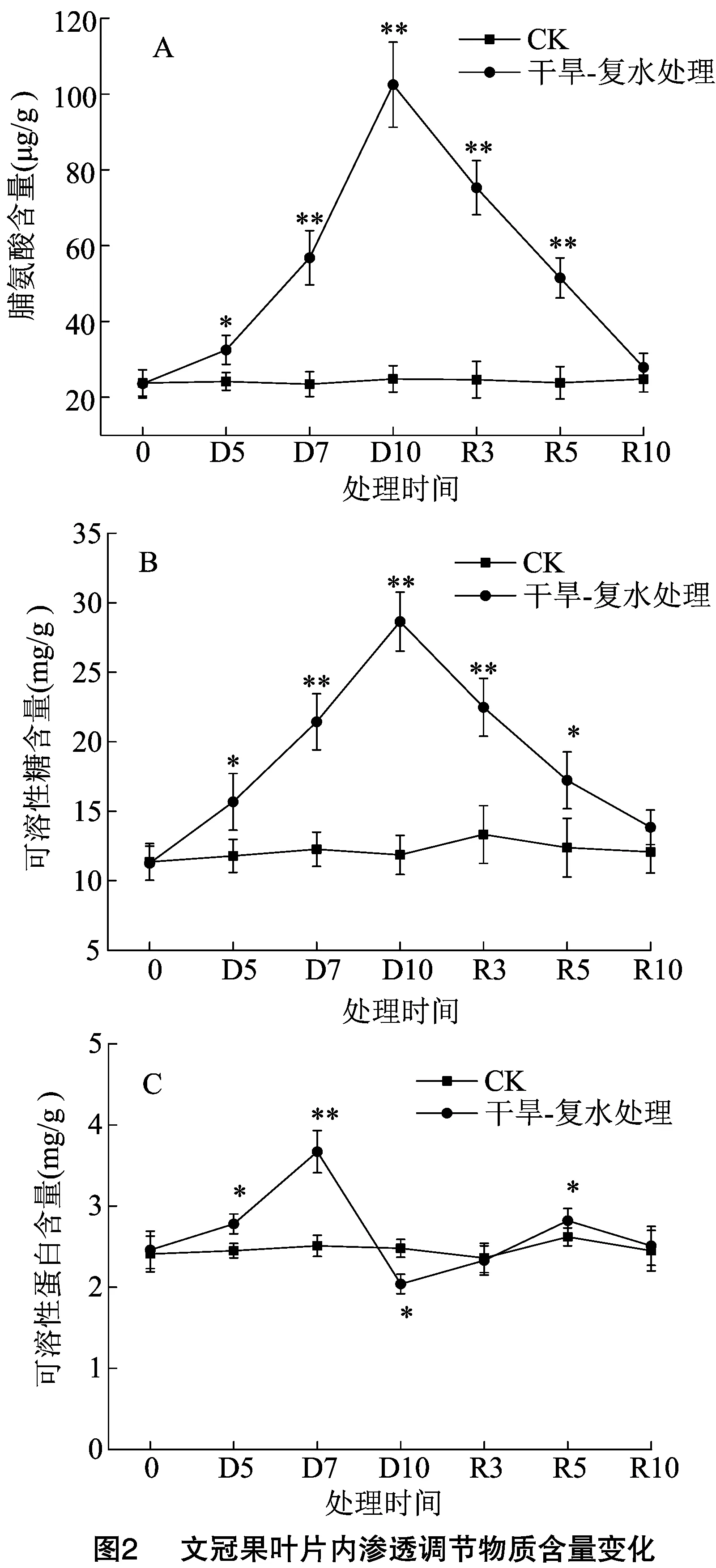
由图2-B可知,文冠果叶片中SS含量的变化趋势与Pro大致相同,土壤干旱程度越大,SS含量越高,与CK相比,D5、D7、D10分别增加了33.02%、74.79%、141.69%。复水处理后,R3、R5、R10时,Pro含量均下降,同样在R10时,叶片内Pro含量与CK组差异不显著。
由图2-C可知,叶片中SP含量的变化趋势与Pro和SS存在一定的差异,土壤干旱程度从轻度到中度时,叶片中SP含量显著增大,然而在重度胁迫时,叶片中SP含量显著下降,出现这一原因可能是在干旱初期,叶片中氧化自由基逐渐增多,为清除自由基,细胞内一些抗氧化酶和蛋白质随之增多,然而在胁迫程度继续增大时,大量的自由基抑制了抗氧化酶和蛋白质的活性,使之急剧下降,从而导致叶片内SP含量显著降低[12]。在复水处理后,R3时SP含量立即恢复到了胁迫前水平,在R5时,叶片内SP含量出现了超补偿性增加,显著大于同期CK组,在R10时,又恢复正常。
2.3 干旱-复水处理对文冠果幼苗叶片MDA含量的影响
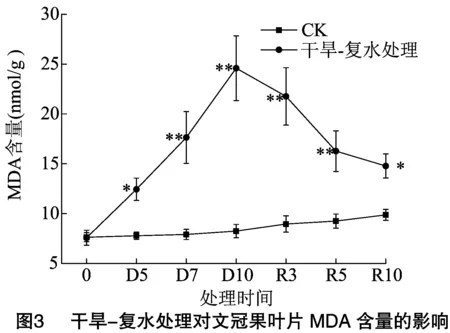
研究表明,干旱胁迫会使植物体内产生大量的氧自由基,尤其是叶片细胞内氧自由基会大量堆积,进而攻击细胞膜产生脂质过氧化物丙二醛(MDA),一定程度上破坏叶片的光合作用,从而抑制植物的生长[13]。由图3可以看出,在试验期间,CK组文冠果叶片内MDA含量有小幅的上升,但变化不显著,而干旱-复水处理组文冠果在干旱胁迫阶段,MDA含量呈现陡坡势增加,与同期CK组相比,D5、D7、D10分别显著或极显著增加了59.89%、123.01%、198.06%。经过复水处理后,从R3到R10,MDA含量显著降低,与D10相比,R3降低了11.51%,R5降低了33.83%,R10降低了39.90%,此外,复水处理阶段,R3、R5、R10与同期CK相比,仍存在显著或极显著差异。结果表明,干旱对文冠果的胁迫作用使叶片内MDA含量显著增加,通过复水处理可以降低MDA含量,然而需要一定恢复时间。
2.4 干旱-复水处理对文冠果幼苗叶片抗氧化酶活性的影响
对干旱胁迫下体内积累的氧自由基,植物有一套完整的抗氧化酶系统,SOD、CAT、POD等组成了抗氧化酶系统,在清除氧自由基过程中发挥着重要作用[14]。由图4可以看出,CK组中文冠果叶片内SOD、CAT、POD活性处在一个较稳定的动态平衡中,而干旱-复水处理组中SOD、CAT、POD活性变化趋势相似,在干旱胁迫阶段,先升高后降低,在复水处理阶段,同样是先升高后降低。
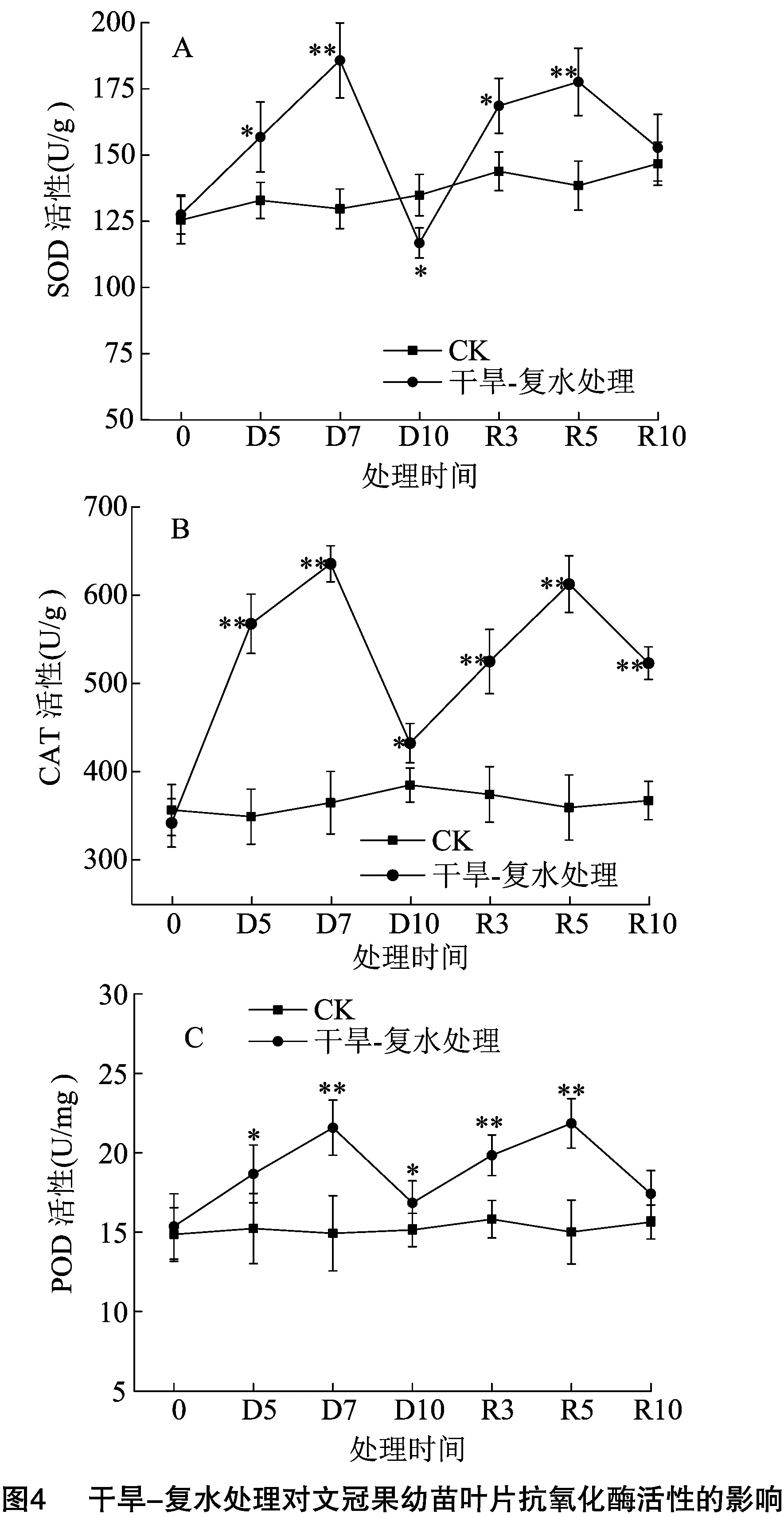
在干旱胁迫阶段,与同期CK相比,D5时,SOD、CAT、POD活性分别显著或极显著升高了18.04%、62.62%、22.59%,D7时,SOD、CAT、POD活性分别极显著升高了43.23%、74.22%、44.54%,而在D10时,SOD、CAT、POD活性均呈现断崖式下降,与D7时相比,分别降低了37.13%、31.99%、21.92%,这一结果与同期叶片内SP含量下降趋势相似,也进一步验证了SP含量下降的原因。
在复水处理阶段,R3、R5、R10时,叶片内SOD、CAT、POD活性均高于同期CK组,R3和R5时,SOD、CAT、POD活性均呈现升高趋势,在R5时达到最大值,分别为177.63、612.47、21.85 U/mg,在R5到R10时,又呈下降趋势,此时的SOD、POD均略高于CK组,但差异不显著,而CAT则与CK组存在极显著差异。出现这一结果原因可能是重度干旱期叶片内氧自由基和MDA大量积累,使抗氧化酶活性显著降低,在复水恢复阶段,干旱胁迫逐渐解除,抗氧化酶活性逐渐增强以清除大量积累的氧自由基和MDA,当细胞内积累的代谢产物被逐渐分解后,细胞的氧化应激作用得到恢复,从而使体内抗氧化酶活性下降,慢慢恢复到正常水平[15]。上述结果表明,干旱胁迫初期显著提高了抗氧化酶活性,在重度胁迫时,抗氧化酶活性显著降低,这一结果在一定程度上反映了文冠果幼苗难以抵抗重度干旱,而复水处理可以缓解干旱的胁迫,通过提高抗氧化酶活性,使细胞内氧化和抗氧化逐渐恢复到动态平衡,从而改善干旱对叶片细胞造成的氧化应激损伤。
2.5 干旱-复水处理对文冠果幼苗叶片叶绿素含量的影响
植物的生长发育离不开光合作用,而叶绿素是光合作用的机器,可以将光能转化为碳水化合物,为植物生长提高能量,在受到环境胁迫时,植物可以动态地调节叶绿素含量和比例,协调控制光合作用强弱[16]。因此,植物体内叶绿素含量变化在一定程度上可以反映环境的胁迫作用。由图5可以看出,叶绿素a、叶绿素b和叶绿素a+b的含量变化趋势大致相同,随着土壤干旱程度增加,其含量均先升高后降低,在轻度干旱时达到最高,分别为1.55、0.38、1.93 mg/g,与CK组相比,均存在显著差异,在重度干旱时,含量均最低,分别为1.32、0.26、1.58 mg/g,同样与CK组相比,均存在显著差异。在复水处理后,R3到R10之间,叶绿素a、叶绿素b和叶绿素a+b的含量均先增加后降低,在R5时,其含量均出现了超补偿效应,与CK组相比,分别增加了6.18%、12.9%、7.26%,在R10时,其含量又恢复到了CK组水平。可见,文冠果在干旱-复水处理期间,叶片内叶绿素含量一直在进行动态调整,经过干旱胁迫后,复水处理可以促进幼苗叶片内叶绿素的合成,从而维持正常的光合作用。

2.6 干旱-复水处理对文冠果幼苗叶片光合参数的影响
由图6-A可知,CK组的Pn在试验期间波动较小,而干旱-复水处理组中,干旱和复水处理均对文冠果叶片Pn产生了较大影响。在干旱胁迫阶段,随着干旱胁迫程度的增加,Pn呈显著下降趋势,D5、D7、D10分别较同期CK组下降了20.65%、54.35%、67.49%。可见,干旱对文冠果叶片的光合作用具有显著负影响。在复水处理阶段,从D10开始复水到R10期间,Pn显著增加,与D10相比,R3、R5、R10的Pn分别增加了1.05、1.64、2.24倍,在R10时,干旱-复水处理组Pn甚至高于CK组,但差异不显著。可见,复水处理能充分缓解干旱对文冠果造成的胁迫作用,在经过10 d的复水处理后,Pn出现了超补偿现象。
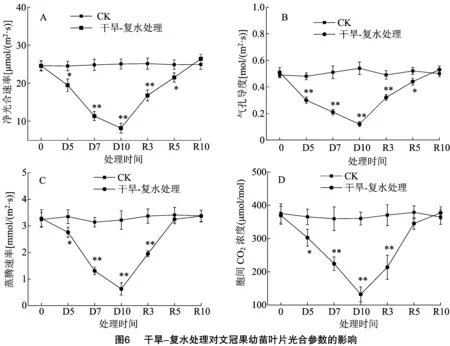
由图6-B可知,文冠果叶片的Gs变化趋势与Pn相似,干旱和复水处理均对Gs造成了显著影响。在干旱胁迫阶段,土壤干旱程度越大,叶片Gs越小,这意味着叶片可通过关闭气孔降低光合作用来响应干旱胁迫作用。与同期CK组相比,D5、D7、D10的Gs分别下降了27.08%、58.82%、77.77%,与Pn相比,干旱胁迫下Gs的下降速度较快,说明在干旱胁迫过程中,叶片气孔的关闭速度大于光合作用的下降速度。在复水处理阶段,复水处理显著或极显著提高了叶片的Gs,随着复水时间的增加,Gs逐渐升高,在R10时,干旱-复水处理组的Gs高于同期的CK组,可见,复水处理下文冠果叶片Gs同样出现了超补偿效应。
由图6-C和图6-D可知,干旱处理下,叶片的Tr和Ci均显著或极显著降低,同样随着干旱胁迫程度的加深,Tr和Ci下降程度越大。在D5、D7、D10时,与同期CK相比,Tr和Ci分别下降了17.61%、58.28%、80.43%和17.17%、37.64%、63.31%。复水处理下,D10到R5时,叶片Tr和Ci呈现陡坡势上升,在R5到R10时,叶片Tr和Ci上升幅度较小,与D10相比,R3、R5、R10时Tr和Ci分别提高了2.09、4.15、4.34倍和0.62、1.61、1.85倍,在R5、R10时,Tr和Ci与同期CK差异不显著。可见,复水处理5 d后,能使干旱胁迫下的Tr和Ci恢复到CK组水平。
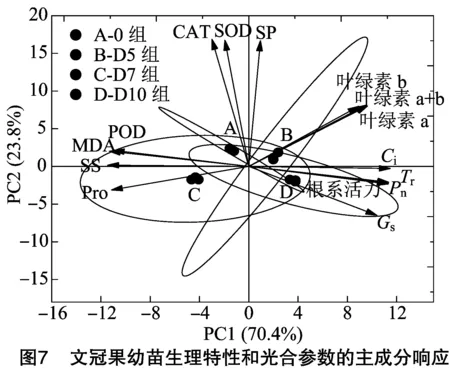
2.7 干旱-复水处理下文冠果幼苗生理特性和光合参数的相关性分析
由图7可知,4个样本A、B、C、D分别在4个不同的象限内,组间样本比较分散,而各组内样本之间距离很短,且所有样本均在95%置信椭圆内,说明组间样本可以通过变量区分开,组内样本间的重复性比较好,均具有统计学意义。从坐标轴上可知,PC1的变异系数为70.4%,PC2的变异系数为23.8%,总共可以解释94.2%以上的方差,说明主成分图里显示的文冠果生理特性指标和光合参数指标能够比较好地对干旱-复水处理作出响应。
从表1可以看出,SS含量与叶绿素含量、光合参数呈现极显著负相关,SP与叶绿素含量呈现一定的正相关,但相关系数均低于0.5,而与光合参数相关系数均低于0.25,显示出较差的相关性;Pro、MDA含量均与叶绿素含量、光合参数呈显著或极显著负相关,SOD、CAT活性与叶绿素含量呈正相关而与光合参数呈现负相关,但相关性均不显著。结果表明,叶绿素含量、光合参数、SS含量、Pro含量、MDA含量对干旱-复水处理下文冠果的响应较大,干旱-复水处理下叶绿素含量、光合参数和SS、SP、MDA含量是耦联在一起的。
3 讨论
3.1 渗透调节作用对文冠果幼苗干旱-复水处理的响应
渗透调节作用是植物应对逆境胁迫维持正常生长的主要响应机制之一,植物体内的渗透调节有2种,一种是由Pro、SS和SP组成的有机渗透调节物质,其主要通过调节细胞质的渗透势来缓解外界环境的胁迫作用;另一种是由大量无机离子组成的无机渗透调节物质,其主要通过调控无机离子浓度维持细胞的膨压,从而提高植物的抗逆性[17]。研究表明,植物叶片中的渗透调节作用主要依靠有机渗透调节物质,而植物根系中的渗透调节作用主要依靠无机离子[18]。干旱对植物造成的渗透胁迫,会使叶片中Pro、SS大量积累,从而提高细胞内的渗透势,保护细胞内正常的活性代谢[19]。Wang等在研究干旱对番茄的胁迫作用时,发现干旱胁迫提高了叶片中Pro、SS含量,干旱程度越大,渗透物质的积累越高[20];Shen等在研究干旱对越南红山茶的作用机制时同样发现,干旱下叶片内渗透调节物质含量明显升高[21]。本研究的结果同样显示,干旱胁迫程度越大,文冠果叶片内Pro、SS含量越高,这与上述学者研究干旱对植物叶片渗透的作用相似。另外,本研究中文冠果叶片中SP含量随着土壤胁迫程度的升高而出现先升高后降低的趋势,这主要是由于在重度干旱胁迫下,叶片内积累的大量活性氧抑制了细胞内抗氧化酶和一些活性蛋白活性,从而导致SP含量下降,这一结果与Song等[22]、马亚珺等[23]研究大豆、紫花苜蓿对干旱胁迫的响应机制结果相同。结果表明,文冠果幼苗在受到干旱胁迫时,可通过调节叶片内Pro、SS和SP的含量来响应干旱的胁迫作用。
当植物所处的逆境胁迫解除后,其独特的自我调节能力和相应的作用机制会发挥作用。研究表明,植物在干旱胁迫后进行复水处理,一定程度上可以缓解幼苗的生长发育能力[24]。徐珍珍等研究显示,紫花苜蓿在干旱胁迫后通过复水处理,其体内Pro、SS、SP可以恢复到正常水平[25];胡艳等研究表明,在干旱胁迫后进行复水处理,黑果腺肋花楸可通过自身调节作用来调控叶片内渗透物质含量,使其适应土壤的干旱胁迫[26]。本研究结果与这些研究结果相似,在本研究中,文冠果在干旱复水处理后,叶片内的Pro、SS和SP含量均逐渐恢复到正常对照组水平,结果表明,文冠果幼苗在受到一定程度的干旱胁迫后可通过复水处理进行缓解,可见,文冠果具有一定的干旱适应能力。
3.2 氧化应激作用对文冠果幼苗干旱-复水处理的响应
植物在干旱胁迫下,细胞内会积累大量的氧自由基,产生氧化胁迫,使细胞内氧化应激水平急剧升高,另外,氧自由基还会诱发膜脂过氧化产生脂质过氧化物丙二醛(MDA),导致细胞膜结构出现一定程度的损伤,从而导致植物的生长发育受到抑制。本研究中,文冠果幼苗在受到不同程度的干旱胁迫时,叶片内MDA含量随着干旱程度加深而逐渐升高,抗氧化酶活性则出现先升高后降低趋势,这主要是因为在轻度干旱和中度干旱时,文冠果叶片内积累的活性氧,使细胞内氧化和抗氧化平衡被打破,氧化应激水平升高,MDA含量随之升高,为防止细胞受到氧化损伤,体内的抗氧化防御系统会被激活,从而提高抗氧化酶(SOD、CAT、POD)活性来清除过多的活性氧,而当文冠果在重度干旱胁迫时,大量积累的活性氧导致细胞受到严重的氧化损伤,抗氧化酶活性反而被抑制。邢钰坤等研究显示,干旱胁迫下内蒙古文冠果幼苗叶片内抗氧化酶活性会急剧上升[27];权伍荣研究同样显示,文冠果幼苗的抗氧化酶活性随干旱程度的加深,抗氧化酶活性会逐渐降低[28]。本研究结果与上述学者的研究结果相似。
研究表明,植物经过干旱胁迫后,采用复水处理,在短期内可以提高抗氧化酶活性,清除过多的活性氧,降低细胞的氧化应激水平,表现一定的缓解干旱胁迫的自我修复能力[29]。本研究结果显示,文冠果在重度干旱后经复水处理,叶片内抗氧化酶活性急剧升高,随着复水时间的延长,抗氧化酶活性又恢复到正常对照组水平。Mu等发现,干旱胁迫初期激活了冬小麦的抗氧化酶活性,在胁迫后期抗氧化活性呈下降趋势,在复水处理后,抗氧化酶活性先升高后降低[30];谢志玉等以二年生文冠果苗为试验材料研究干旱和复水处理,结果显示,随着干旱胁迫程度提高,CAT活性先升高后降低,复水处理后,CAT活性显著升高,后逐渐降低[31]。本研究结果均与这些研究结果一致。
3.3 光合作用对文冠果幼苗干旱-复水处理的响应
在干旱胁迫下,保持较高的光合作用对维持植物的生长发育和作物产量具有重要意义。在逆境胁迫下,光合系统活性会被抑制,主要体现在水分利用率下降、光合速率下降、叶片气孔导度关闭,蒸腾作用减弱,胞间CO2浓度降低等。研究表明,干旱胁迫会显著降低植物的光合性能,且胁迫作用越强,光合性能下降趋势越明显,然而在复水后光合性能会产生过补偿或补偿效应[32]。植物在轻度的干旱胁迫下,光合作用下降主要是叶片气孔关闭所致,而在重度干旱时,则是由非气孔因素引起。为了适应干旱胁迫环境,植物通过降低蒸腾作用、关闭气孔等来降低光合作用,对体内的能量进行重新分配。本研究结果显示,随着干旱胁迫程度增加,文冠果叶片的Pn、Gs均显著降低,在轻度胁迫时,Gs的下降速度明显快于Pn,此时气孔因素是导致文冠果光合作用下降的主要因素,在重度胁迫时,观察到Pn、Gs的下降速度较一致,这可能是由于体内复杂的抗旱机制起着主要作用。Tr和Ci同样随着干旱程度的加大而逐渐降低,说明文冠果在受到干旱胁迫后可通过降低Tr和Ci来减少植物体内水分的丧失,从而维持一定的光合作用,本研究结果与Hao等[33]、Wang等[34]对干旱复水下小粒咖啡和抗旱稻的光合作用结果相似。在干旱后的复水处理阶段,随着复水时间的增加,文冠果Pn、Gs、Tr和Ci均逐渐恢复到同期对照组水平,可见,短期的重度干旱对文冠果的胁迫作用是可逆的。此外,文冠果在干旱胁迫下,叶绿素含量还会出现一定变化。本研究结果显示,文冠果在干旱胁迫后,叶片内叶绿素a、叶绿素b和叶绿素a+b含量均发生了不同程度的变化,轻度干旱时,叶绿素含量出现小幅上升,在中度干旱和重度干旱时,叶绿素含量为下降趋势,在复水处理后,叶绿素含量又呈现上升趋势,在复水后10 d时,叶绿素含量维持在正常对照组水平。南思睿等研究表明,新疆大叶苜蓿在干旱7 d时,叶绿素含量随着干旱胁迫程度升高而降低,在复水后3 d时,叶绿素含量与对照组差异不显著[6],本研究结果与之类似。可见,复水处理对干旱后文冠果叶绿素含量的恢复具有一定作用。
4 结论
随着土壤干旱胁迫程度的增加,吉林文冠果幼苗的根系活力逐渐下降,叶片中Pro、SS、MDA含量逐渐升高,SP含量和SOD、CAT、POD活性以及叶绿素含量均先升高后降低,叶片光合参数Pn、Gs、Tr和Ci则逐渐下降,另外,通过相关性显示,叶绿素含量、光合参数和SS、SP、MDA之间的耦联较好。在复水后,根系活力逐渐上升,MDA含量逐渐下降,SOD、CAT、POD活性先升高后降低,但在复水10 d时,根系活力、MDA含量、CAT活性均未恢复到正常对照组水平,而叶片中Pro、SS、SP、叶绿素含量和SOD、POD活性以及光合参数均恢复到了同期对照组水平。结果表明,文冠果幼苗在复水处理后,其生理特性指标均出现了补偿或过补偿现象,由此反映出吉林文冠果幼苗具有较强的干旱适应性和复水后的自我调节能力。
