干旱-复水处理下玉米差异表达bHLH 基因的鉴定与分析
2023-10-18谢小文张心悦付家旭温鹏飞王同朝
谢小文,张心悦,付家旭,邵 京,温鹏飞,王同朝,卫 丽
(河南农业大学 农学院,河南 郑州 450046)
玉米(Zea mays)是我国主要的粮食作物,对我国粮食安全和国民经济发展起着重要的作用。玉米是高耗水作物,对干旱胁迫较敏感[1-2]。干旱胁迫下,玉米植株为应对水分缺失造成的压力,在表型、代谢、基因及蛋白质水平会产生相应的应急反应[3-4]。干旱胁迫下,细胞膜上的信号蛋白质会感知和捕获到响应信号并通过级联反应向下传导,激活抗旱相关基因的表达[5]。研究表明,响应干旱胁迫的基因主要包括蛋白激酶基因、蛋白磷酸酶基因和转录因子基因等[6-8]。其中,转录因子在干旱胁迫响应中具有重要作用,它通过与下游靶基因启动子中的顺式作用元件结合,调控干旱胁迫相关基因的表达,减轻干旱胁迫对玉米生长发育的影响[9-11]。目前,已报道的与干旱胁迫相关的转录因子家族有bZIP(Basic leucine zipper)[12-15]、AP2/ERF(APETALA2/ethylene responsive factor)[16-17]、MYB(V-myb avian myeloblastosis viral oncogene homolog)[18]、bHLH(Basic helix-loop-helix)[19-26]等。其中,bHLH 是植物中重要的转录因子家族之一,因含有约60个氨基酸组成的bHLH 结构域而得名,其不仅参与植物的生长发育及信号转导,而且调控植物对干旱等非生物胁迫的响应[19-26]。OsbHLH148基因通过参与茉莉酸信号途径增强了水稻(Oryza sativaL.)对干旱的耐受性[24];过表达番茄(Selaginella lepidophylla)SlbHLHopt基因促进了拟南芥(Arabidopsis thaliana)植株生长发育,提高了水分利用效率和对干旱、高盐的耐受性[26]。前期,河南农业大学农学院旱作节水课题组以豫882 为试验材料,在三叶期进行干旱-复水处理,取胁迫处理60 h、96 h 和复水3 d 的叶片进行转录组测序分析[12],得到11 002个差异表达基因,其中转录因子基因有556 个[27]。在此研究基础上,对干旱-复水处理下玉米中差异表达的bHLH 基因进行鉴定,并对其蛋白质理化性质、系统进化、染色体分布和复制关系、基因结构和蛋白质保守基序、启动子区顺式作用元件及干旱-复水处理下的表达量等进行分析,为bHLH 基因的进一步利用提供理论依据。
1 材料和方法
1.1 试验材料与干旱处理
供试材料为玉米品种豫882,选取籽粒饱满、大小均匀一致的种子灭菌后播种在土壤∶蛭石为3∶1的混合土样中,放置于光照培养箱中[温度28 ℃/22 ℃,光照周期14 h/10 h,光照强度300 μmol/(m2·s),相对湿度60%]。待玉米生长至两叶一心时,将玉米幼苗移栽到霍格兰(Hoagland)营养液中,营养液每2 d更换一次。玉米生长至三片叶展开时,将长势一致的幼苗分为两部分,一部分转移到含有20%聚乙二醇(PEG)6000 的营养液中,模拟干旱处理,分别处理60 h 和96 h,然后复水3 d,记为T60、T96、TR3d;对照组不进行干旱处理,分别记为CK60、CK96、CK3d。取叶片,用液氮速冻后置于-80 ℃冰箱中保存。
1.2 玉米bHLH基因的鉴定
利 用Pfam 数 据 库(http://pfam.xfam.org/)和SMART(http://smart.embl-heidelberg.de/)在线网站分析玉米bHLH 基因编码蛋白质,去除冗余和重复的基因并删除缺少完整bHLH 结构域的蛋白质序列。通过RPKM 算法计算对照组与处理组基因表达量之间的差异倍数,以∣log2(差异倍数)∣≥1 为标准,筛选干旱-复水处理下玉米中差异表达的bHLH基因,并 参 照Ensembl plants(http://plants.ensembl.org/index.html)和MaizeGDB(https://www.maizegdb.org/)数据库进行命名。
1.3 玉米bHLH基因的生物信息学分析
1.3.1 蛋白质分子质量和等电点 使用Protparam(https://web.expasy.org/protparam)预测并分析玉米bHLH 基因编码氨基酸的数目、蛋白质分子质量及等电点。
1.3.2 系统进化 从玉米数据库MaizeGDB 中下载差异表达的bHLH 基因编码的蛋白质序列,从数据库TAIR(https://www.arabi-dopsis.org/)中下载拟南芥同源bHLH 蛋白序列。利用DNAMAN 软件对玉米和拟南芥bHLH 蛋白的氨基酸序列进行比对,采用MEGA 7.0 软 件(http://www.megaso ftware.net/history.php)以邻接法构建系统进化树,步长值设为1 000。
1.3.3 基因在染色体上的定位和复制关系 从NCBI 下载玉米基因组注释信息,使用TBtools 软件分析差异表达的bHLH 基因在染色体上的分布和复制关系。
1.3.4 基因结构和蛋白质保守基序(Motif) 利用MEME Suite 5.4.1(https://meme-suite.org/meme/)[28]在线预测玉米差异表达bHLH 蛋白的保守基序,保守基序数目设为10 个,其他参数均为默认值;从玉米基因组数据库MaizeGDB 中提取差异表达的bHLH 基因的编码序列(CDS),利用GSDS(http://gsds.gao-lab.org/)分析bHLH 基因的外显子-内含子结构,利用TBtools 软件对上述结果进行可视化分析。
1.3.5 启动子区域顺式作用元件 从数据库Ensembl plants 中获取玉米bHLH 基因的启动子序列(ATG 上游2 000 bp),并通过Plant CARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对bHLH基因启动子区域顺式作用元件进行分析。
1.4 玉米bHLH基因的表达分析
对玉米bHLH 基因的表达量数据进行均一化处理,利用Tbtools 软件对基因的表达量进行热图分析。
1.5 RNA 的提取及实时荧光定量PCR(qRTPCR)分析
根据Ensembl plants 中检索到的候选bHLH 基因的cDNA 序列,使用Primer Premier 5.0 进行qRTPCR 引物设计(表1)。利用Trizol 试剂提取叶片总RNA,使用PrimeScriptTMRT reagent Kit with gDNA Eraser(TaKaRa)反转录试剂盒合成cDNA。根据TB Green Premix ExTaqTMⅡ(TaKaRa)试剂盒说明书进行qRT-PCR,采 用 玉 米18S RNA(GenBank No.AF168884.1)作为内参基因。qRT-PCR 反应体系为20 μL:cDNA 1 μL,TB Green Premix ExTaqTMⅡ10 μL,上、下游引物(10 μmol/L)各0.8 μL,ddH2O 7.4 μL。qRT-PCR 反应程序:95 ℃30 s;95 ℃5 s,60 ℃30 s,40 个循环。试验进行3 次生物学重复,采用2-ΔΔCt方法计算基因的相对表达量。

表1 玉米bHLH基因qRT-PCR引物序列Tab.1 qRT-PCR primer sequences for bHLH genes in maize
1.6 数据分析
使用SPSS 26 软件单因素方差分析对基因的相对表达量进行差异显著性分析。
2 结果与分析
2.1 干旱-复水处理下玉米差异表达bHLH基因的鉴定及理化性质分析
本研究鉴定出51 个具有完整bHLH 结构域并在干旱-复水处理下玉米中差异表达的bHLH 基因(表2),其编码氨基酸数目在80(ZmbHLH25)~705个(ZmbHLH12),蛋白质分子质量在21.26(ZmbHLH41)~92.17 ku(ZmbHLH25),等电点在4.54(ZmbHLH81)~12.41(ZmbHLH25)。
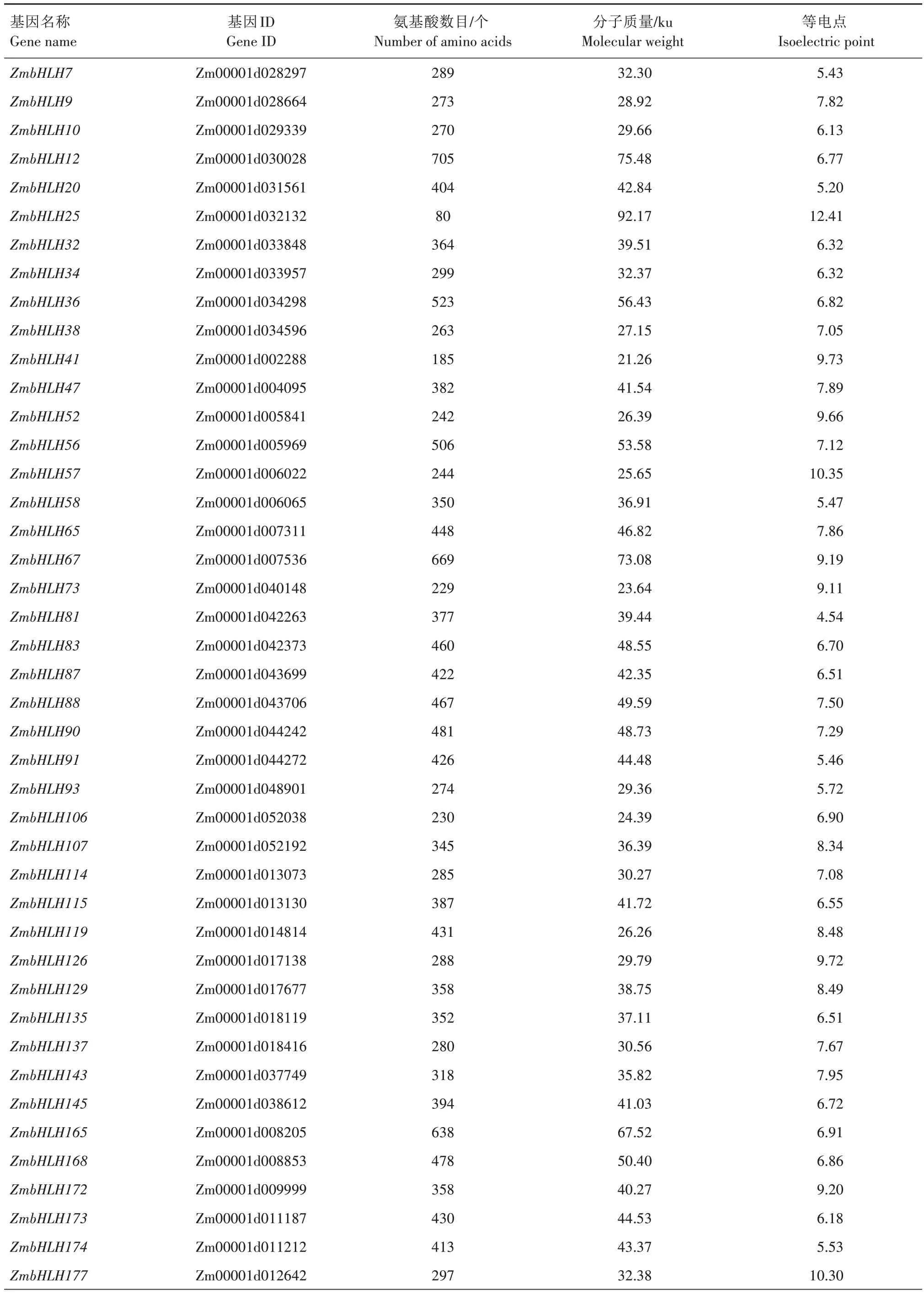
表2 干旱-复水处理下玉米差异表达bHLH基因的基本信息Tab.2 The basic information of differentially expressed bHLH genes in maize under drought-rewatering treatment
2.2 干旱-复水处理下玉米差异表达bHLH基因系统进化分析
本研究筛选出38个干旱-复水处理下玉米差异表达bHLH 基因的拟南芥同源基因,构建玉米、拟南芥bHLH 蛋白系统进化树(图1)。由图1可知,51个玉米bHLH 蛋白分为16 个亚族,其中,最大的亚族为Ⅺ,含有9 个bHLH 蛋白;最小的亚族为Ⅵ、Ⅷ、Ⅸ和ⅩⅢ,均只有1个bHLH 蛋白。每个亚族均含有至少1 个拟南芥bHLH 蛋白,说明玉米和拟南芥的bHLH蛋白之间存在平行进化关系。
2.3 干旱-复水处理下玉米差异表达bHLH基因染色体定位及复制关系分析
由图2 可知,51 个干旱-复水处理下玉米差异表达bHLH 基因在10 条染色体上呈不均匀分布。其中,1 号染色体上有10 个bHLH 基因,2 号染色体上有8 个bHLH 基因,而7 号染色体上没有bHLH 基因。发生了7 对复制事件,包括3 号染色体上的ZmbHLH91基因和8 号染色体上的ZmbHLH174基因,6 号染色体上的ZmbHLH143基因和8 号染色体上的ZmbHLH168、ZmbHLH172基因,1 号染色体上的ZmbHLH12基因和9 号染色体上的ZmbHLH185基因,1 号染色体上的ZmbHLH20基因和2号染色体上的ZmbHLH58基因,3号染色体上的ZmbHLH90基因和8 号染色体上的ZmbHLH173基因,1 号染色体上的ZmbHLH20基因和3 号染色体上的ZmbHLH83基因。
2.4 干旱-复水处理下玉米差异表达bHLH基因结构和蛋白质保守基序分析
由图3 可知,51 个干旱-复水处理下玉米差异表达bHLH 基因的外显子数目差别很大,9 个bHLH基因只有1 个外显子,如ZmbHLH25、ZmbHLH52、ZmbHLH87、ZmbHLH90和ZmbHLH173等;27 个bHLH基因含有2~5个外显子,15个bHLH基因含有6个及以上外显子。
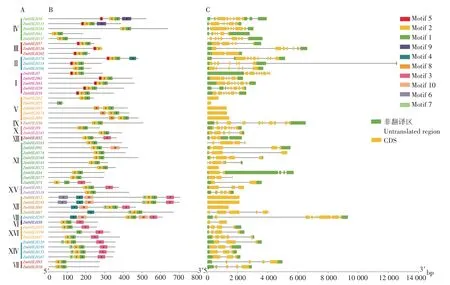
图3 干旱-复水处理下玉米差异表达bHLH基因结构及蛋白质保守基序分析Fig.3 Analysis of differentially expressed bHLH gene structure and encoded protein’s conserved motif in maize under drought-rewatering treatment
对51 个干旱-复水处理下玉米差异表达bHLH蛋白的保守基序进行分析(图3),发现Motif 1 和Motif 2 在51 个bHLH 蛋白中出现的频率较高,48 个bHLH蛋白同时含有Motif 1和Motif 2;其次是Motif 3和Motif 5,分别出现在15 个和12 个bHLH 蛋白中;出现频率最少的是Motif 6 和Motif 9,分别出现在2个bHLH蛋白中。
2.5 干旱-复水处理下玉米差异表达bHLH基因启动子区域顺式作用元件分析
为进一步预测玉米bHLH 基因的功能,对51 个干旱-复水处理下玉米差异表达bHLH 基因启动子区域的顺式作用元件进行分析。由图4 可以看出,干旱-复水处理下玉米差异表达bHLH 基因启动子区域含有多个响应植物激素和非生物胁迫的顺式作用元件。其中,49 个bHLH 基因含有脱落酸响应元 件 ABRE, 如ZmbHLH10、ZmbHLH106、ZmbHLH107、ZmbHLH114、ZmbHLH119等,其中含有5 个以上ABRE 元件的基因有25 个,而ZmbHLH185、ZmbHLH199、ZmbHLH34基因分别含有 14、12、10 个 ;ZmbHLH12、ZmbHLH126、ZmbHLH129、ZmbHLH135、ZmbHLH137等47 个bHLH 基因均含有茉莉酸响应元件CGTCA-motif 和TGACG-motif;另外,大部分bHLH 基因的启动子区域中都存在水杨酸响应元件(TCA-element)、脱水响应元件(MBS)、生长素响应元件(AuxRR-core)、赤 霉 素 响 应 元 件(GARE-motif 和P-box),表 明bHLH 基因参与植物激素合成和逆境胁迫响应等生物学过程。

图4 干旱-复水处理下玉米差异表达bHLH基因启动子区域顺式作用元件分析Fig.4 Analysis of cis-acting elements in the promoter region of differentially expressed bHLH genes in maize under drought-rewatering treatment
2.6 干旱-复水处理下玉米差异表达bHLH基因的表达分析
依据转录组结果,在干旱-复水处理下,51个玉米差异表达bHLH 基因呈现不同的表达模式。由图5可知,在干旱处理60 h、96 h和复水3 d 3个处理时间点,表达量提高的基因分别有23、18、17 个,表达量下降的基因分别有28、33、34个。干旱-复水处理下,ZmbHLH20、ZmbHLH25、ZmbHLH9、ZmbHLH137、ZmbHLH178等14 个基因在干旱处理60、96 h,表达量高于对照,复水3 d,表达量低于对照,说明这些基因正向响应干旱胁迫;ZmbHLH58、ZmbHLH87、ZmbHLH36、ZmbHLH106等14 个基因在干旱处理60、96 h,表达量低于对照,复水3 d,表达量高于对照,呈负向响应模式;而ZmbHLH90、ZmbHLH165、ZmbHLH83、ZmbHLH177、ZmbHLH168等基因的表达呈无规律变化。
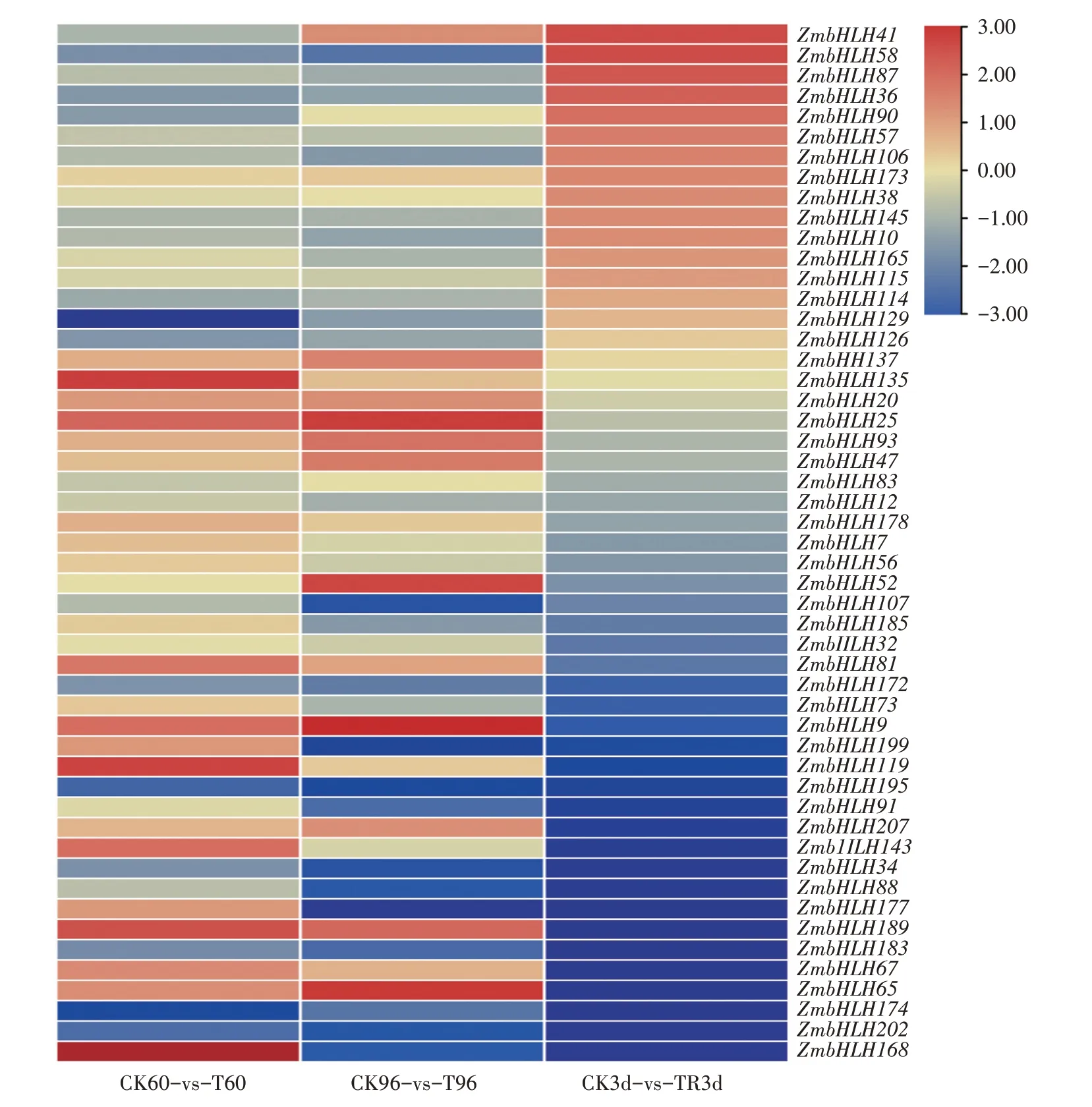
图5 干旱-复水处理下玉米差异表达bHLH基因的表达分析Fig.5 Analysis of expression of differentially expressed bHLH genes in maize under drought-rewatering treatment
为验证转录组数据的准确性,随机挑选ZmbHLH9、ZmbHLH25、ZmbHLH65、ZmbHLH93、ZmbHLH58和ZmbHLH106基因进行qRT-PCR 分析(图6),结果表明,在干旱处理60、96 h,ZmbHLH9、ZmbHLH25、ZmbHLH65、ZmbHLH93基因的表达量高于对照,而ZmbHLH58和ZmbHLH106的表达量低于 对 照 ;复 水 3 d,ZmbHLH9、ZmbHLH25、ZmbHLH65andZmbHLH93基因的表达量低于对照,而ZmbHLH58和ZmbHLH106基因的表达量高于对照。这些基因的qRT-PCR 结果与转录组分析结果一致,说明转录组分析结果可靠。

图6 干旱-复水处理下部分玉米差异表达bHLH基因的表达验证Fig.6 Expression validation of partial differentially expressed bHLH genes in maize under drought-rewatering treatment
3 结论与讨论
玉米是高耗水作物,目前我国已有超过50%的玉米种植区受到严重的干旱,对玉米生产造成了严重威胁,干旱已成为限制玉米生产发展和产量提高的第一要素。在现有水资源状况下,如何最大限度地提高玉米抗旱能力,实现玉米高产稳产,是破解我国农业用水短缺和维持玉米高产稳产的关键问题。山仑院士最早提出生物节水的概念,即在一定干旱胁迫范围内,进行干旱-复水处理,巧用生物自身的补偿或超补偿效应实现节水,但其核心内容是生物本身要有抗旱性[29]。利用作物自身的生理和基因潜力来抵御干旱是提高作物水分利用效率的根本途径。因此,鉴定和筛选抗旱基因是增强玉米抗旱性的基础。
诸多研究结果表明,bHLH 转录因子在植物对干旱胁迫的应答中起着积极的作用,可提高植物对非生物胁迫的耐受性[30-31]。本研究从前期干旱-复水处理的玉米转录组数据中鉴定出51 个差异表达的bHLH 基因,参考ZHANG 等[32]玉米bHLH 蛋白的分类结果,51 个bHLH 蛋白分为16 个亚族。第Ⅳ亚族 的 3 个 成 员(ZmbHLH36、ZmbHLH115、ZmbHLH165)均包含保守的活性光敏色素A(APA)或活性光敏色素B(APB)结构域,推测其可能调控光调节基因。与AtbHLH27、AtbHLH35 蛋白[33]相邻且聚集在第ⅩⅥ亚族上的ZmbHLH47、ZmbHLH95 和ZmbHLH99 蛋白,推测也具有抗非生物胁迫方面的作用。保守基序分析结果表明,Motif 1 和Motif 2 在51 个bHLH 蛋白中出现的频率较高,部分基序表现出亚家族的特异性。对51 个bHLH 基因结构进行分析,发现不同亚族bHLH 基因外显子数目差别较大,但同一亚族bHLH 转录因子的保守基序与基因结构比较相近,说明同一亚族成员之间可能存在着密切的进化关系。
51 个玉米bHLH 基因的启动子区域存在许多顺式作用元件,包括脱落酸响应元件(ABRE)、生长素响应元件(AuxRR-core)、水杨酸响应元件(TCAelement)、茉 莉 酸 响 应 元 件(TGACG-motif 和CGTCA-motif)、赤霉素响应元件(GARE-motif 和Pbox)等。ABRE 参与脱落酸和干旱胁迫反应[34],49个玉米bHLH 基因启动子区包含多个ABRE 元件,推测这些基因可能与植物的逆境胁迫响应有关。
本研究分析51 个玉米bHLH 基因在干旱处理60 h、96 h 和复水3 d 条件下的表达模式发现,ZmbHLH9、ZmbHLH25、ZmbHLH93、ZmbHLH47等14 个基因正向响应干旱胁迫,ZmbHLH58、ZmbHLH87、ZmbHLH36等14 个基因负向响应干旱胁迫,其余23 个基因的表达呈不规律变化,研究结果为进一步解析bHLH基因的功能奠定了基础。
