藻类水华聚散迁移机制及其数值模拟研究进展
2023-10-12杨正健刘德富颜碧娥
杨正健 章 睿 刘德富 马 骏 颜碧娥
(1.三峡大学 三峡水库生态系统湖北省野外科学观测研究站, 湖北 宜昌 443002;2.湖北工业大学 土木建筑与环境学院, 武汉 430068)
氮磷污染导致的水体富营养化及藻类水华严重威胁饮用水安全和水生态安全,已成为世界共同面临的水生态环境问题[1].大量研究已阐明了生源要素、光照、水温、生物捕食、水动力等因素与藻类水华生消的关系[2],但“水华在短期内(约1~2 d)突然暴发或消失”这一现象的发生机制目前仍不明确[3-4],成为突破“水华预测预报和防控”这一难题的关键.这种水面藻类浓度骤增骤减现象用经典的藻类生长动力学理论很难解释,近年来被逐步认为其与不同环境下藻类颗粒聚散迁移有关,成为水华生消机理和水华预测预报新的研究方向[3],也是相关研究的难点.本文通过整理国内外有关水华的研究论文,分析藻类聚散迁移的关键机制并阐述藻类水华预测预报现状,以此提出目前难以精准模拟水华的主要原因并为后期研究提出建议,以期为提高藻类水华的预测预报精度并改进水华防控措施提供新的借鉴.
1 有关藻类水华论文发表情况及研究趋势
水华(algal bloom)是因水体氮、磷等营养盐过高导致浮游藻类迅速繁殖聚集并显著改变水色的一种自然现象.藻类不同,水华表观和危害程度也不同,其中以微囊藻水华为代表的部分蓝藻水华因能产生神经毒素并在水面形成一层绿色漂浮物[2],且死后大量消耗溶氧,进而对水生态系统产生严重威胁,被称为“生态癌症”,已成为全世界共同面临的严重生态环境问题之一[1].截至2022年底,在Web of Science中以“algal bloom”或“algae bloom”为主题检索的科技论文已超过23 000篇,且目前仍以超过2 000篇/a的速度快速增长(如图1所示),说明“水华”问题仍是国际科技界关注的热点和重点.中国从改革开放后逐步重视生态环境保护工作,统计“中国知网”相关数据发现,有关“水华”的中文论文发表量从2000 年的49篇/a,十年内迅速增长到498篇/a,之后一直维持在较高水平,这说明正处于水生态环境治理“黄金时期”的我国仍高度关注“水华”问题.
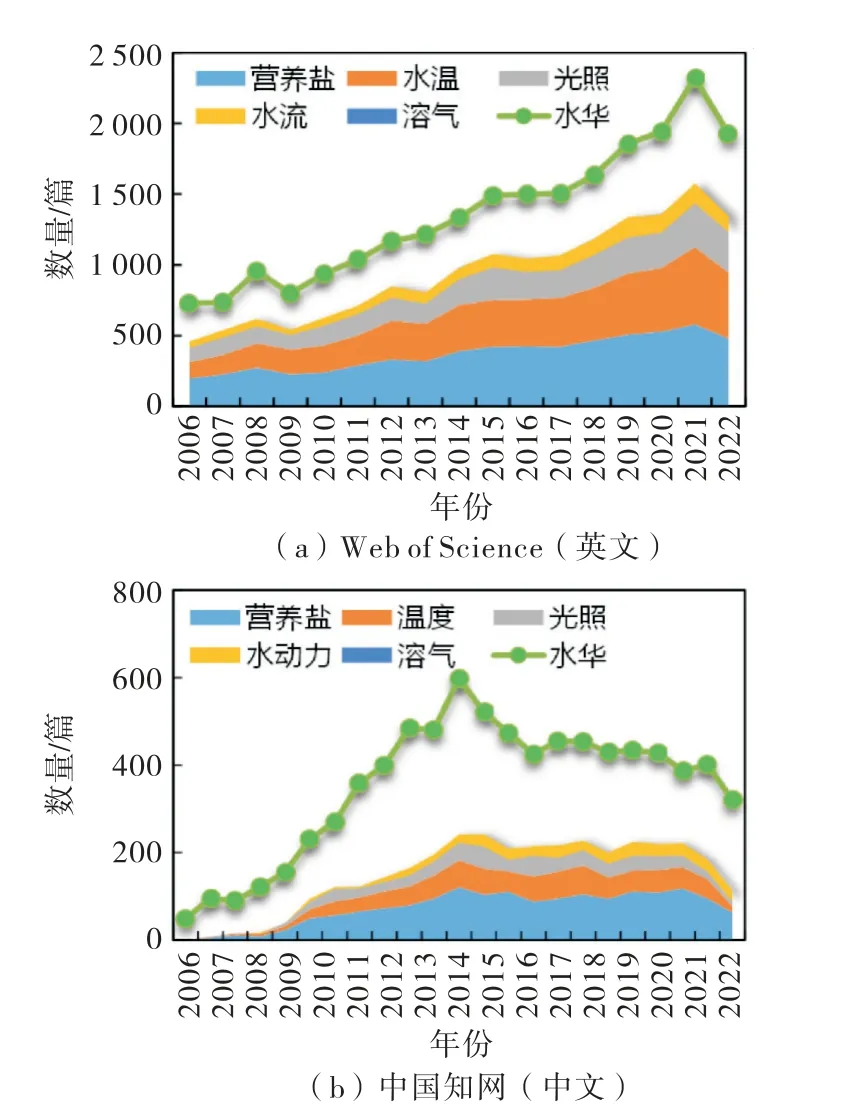
图1 有关“水华”研究论文统计分析图
通过关键词检索发现,国内外关于水华的研究多聚焦于探究营养盐、水温、光照、水流等环境因子与藻类生长或水华生消的关系[2].营养盐是藻类生长的物质基础,水体富营养化一般是水华发生的前提条件.因藻类生长所需的碳:氮:磷之比接近106∶16∶1[5],所以在碳较为充足的开放水体中,氮、磷等元素的贫丰或相对组成可以决定浮游植物群落结构的演变,进而决定不同藻类水华发生程度[6].水温决定了藻类进行光合作用和呼吸作用过程中酶的活性,因而不同藻类的适宜生长温度也不相同,进而导致了不同季节水华优势种的不同,例如蓝藻水华一般发生在高温水体,而硅、甲藻水华则在相对低温水体中发生[7].光照因能影响浮游藻类的光合速率而成为藻类生长的重要环境因子[8],一般将有效光合强度大到能进行藻类光合作用的水层称之为“真光层(euphotic zone)”,并将“真光层”深度大小作为水华发生潜势的判定指标[9].大量水流与水华的关系研究聚焦于宏观水流指标下的藻类生长过程分析,例如“临界流速”概念[10],“水体滞留时间”概念[11],临界层假设[12],水体冲淡假设[13]等.此外,浮游动物与藻类的食物链关系[14],微量元素对藻类生长的刺激作用[15],微生物与藻类的耦合互馈关系[16]等也被认为可影响水华的生消.
虽然国内外相关研究阐明了很多决定水华生消的关键机制问题,但世界范围内的水华特别是蓝藻水华问题并未得到根本解决[1],说明水华发生的某个关键科学问题目前仍未探明,亟待寻求研究水华生消机制的新途径.早期研究多关注于环境因子与水华生消的宏观关系,但最新研究发现从藻类个体大小角度观测到的藻类聚散迁移等微观过程与水华生消密切相关[17],已成为水华研究的重要发展方向.
2 藻类聚散迁移关键机制研究进展
大量观测表明,在水华发生期间水体表面藻类密度往往在短期内(1~2 d)内由102~103cell/L 迅速达到105~109cell/L,部分藻类(如微囊藻)还会在水面形成藻类漂浮物[18],即为水华的典型特征;在遇到降雨、降温等特殊环境改变时有些藻类形成的水面漂浮物又会骤然消失[19].这种水面藻类浓度骤增骤减现象用经典的藻类生长动力学理论很难解释,近年来被逐步认为其与不同环境下藻类颗粒聚散迁移有关[3],是目前相关研究的难点.
部分藻类具备的浮力调节能力被认为是驱动藻类垂向迁移的主要因素.蓝藻可利用伪空泡(gas vesicle)实现浮力调节[2],伪空泡是蓝藻或细菌体内形成的由蛋白质构成的两端为锥形中间为圆柱体的中空、充有气体的细胞内结构[20],如图2所示.
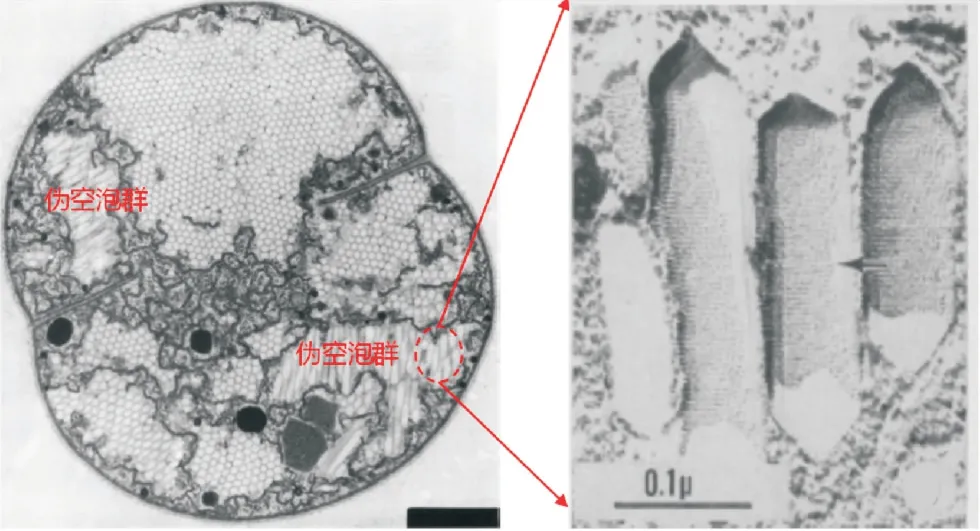
图2 伪空泡在蓝藻细胞内的分布示意图[20]
蓝藻能够通过自身新陈代谢调节伪空泡的密度,形成上浮或下沉,从而在不均匀环境下更容易获得光照、营养盐等资源[2],目前也被认为是蓝藻能够上浮至水面形成水华漂浮物的主要原因.部分具有鞭毛结构的藻类也可利用鞭毛在水体中实现主动迁移,如甲藻具有鞭毛,能够在水中上下移动以选择适宜的生长环境[21];部分衣藻、裸藻等也能够利用鞭毛运动至水面形成水华优势藻类[22].此外,有些藻类也可通过分泌胞外多糖(extracellular polysaccharides,EPS)调节浮力大小,如微囊藻可通过胞外多糖实现藻细胞的聚合,使蓝藻形成数十个至数万细胞不等的蓝藻颗粒群体以获取更大的浮力[23],这些胞外多糖还具有保护蓝藻细胞免于病毒、捕食或脱水等危害的作用[24].此外,藻类细胞内的糖类物质累积作用也可通过改变细胞密度并与伪空泡一起实现藻类的浮力调节,如某些束丝藻白天光合作用合成的糖类以淀粉的形式积累而使丝状体密度增加,晚上呼吸作用消耗淀粉使细胞密度下降[25].
实际上,藻类虽然可通过自身生理结构实现浮力调节,但其能否最终上浮至水面形成水华还与水动力密切相关.例如风浪作用可使蓝藻细胞(细胞团)受到湍流向下的裹挟作用,只有当湍流裹挟作用小于蓝藻细胞浮力作用时才会形成蓝藻水华;也正因为波浪与湖流发育具有较强的时空异质性,才使一般蓝藻水华呈现多点暴发的特点[3].同时,不同藻类对水体紊动的抵抗差异决定了不同水动力条件下的水华优势种不同,如甲藻因鞭毛运动能力较弱,只能在水体弱分层时占优[26];但蓝藻因伪空泡和胞外多糖的作用具有更强的浮力调节能力,既能在高温且强分层水体中存活[27],又能在一定的风扰作用下漂浮至水面形成水华[3].基于此,大批学者针对不同水体紊动模式,形成了多种模型来预测藻类生长和水华生消过程,其中“临界层理论”是目前公认的预测水华生消的经典模型,被广泛应用并解释了多种水华生消过程[13].此外,近年的最新研究还表明,有些藻类还可通过演化形成不同生理结构来抵御不同的水流紊动,如为了躲避“紊流涡旋”作用,裸藻演化形成了非对称结构[28]、衣藻演化形成鞭毛对称运动特性[22]等,如图3所示,这种“微观水流与藻类生理的关系”可能是决定藻类聚散迁移和水华生消的关键因素.

图3 “旋涡陷阱”与衣藻运移的关系[22]
虽然藻类可通过浮力调节作用实现水体垂向迁移,但相关研究估算发现以藻类生理结构改变可形成的运移速率不超过mm/s量级,但藻类水华短期“暴发”并形成漂浮物的现象至少需要dm/s量级以上的运移速率才能实现[3].且有些不具备伪空泡等生理结构的藻类在短期内也能形成水华漂浮物现象[29].因此,截至目前关于藻类聚散迁移导致水华“暴发”或突然“消失”的现象仍未有公认的合理解释.秦伯强等[3]结合太湖蓝藻水华发生过程提出一种“藻类聚团”假说,但藻类为何能在短期内相对定向聚团,以及这种聚团藻类仅靠藻团浮力就能实现快速上浮的机制仍不清楚.
3 藻类水华预测预报方法研究进展
目前关于藻类水华预测预报方法大致可分为两类:一是基于物理过程的数值模型,是研究富营养化状态及预测预报水华发生的有效工具,其研究、预测精度取决于对事物规律的认识;二是基于数据挖掘的方法,其精度取决于已有数据的数量及质量.
基于过程的数值模型方法从水动力计算出发,通过连续性方程、动量方程和能量方程等,得到水流场,再结合输移扩散方程等求解温度场、营养盐浓度场、水华生消过程等.水流场准确模拟是水华精准预测预报的前提,最早可追溯到Hansen和Rattay利用一维垂向模型对Navier-Stokes方程进行求解,并应用于盐度和垂向速度的计算[30].为了研究水下地形摩擦系数对水动力场的影响,建立了北美安大略湖的二维湖泊水动力模型,奠定了平面二维水动力模拟的基础[31].后续学者在此基础上,逐渐模拟研究了北美的艾利湖[32]和日本的琵琶湖[33]内部水动力要素之间的相互关系,逐步发展了浅水湖泊平面二维水动力模型,并较为详细地研究了水动力模型内部各个影响因子的相互作用关系,为后人提供了较为重要的参考价值.近几十年来,国内外学者针对河流、水库、河口等水域的水动力过程及水质变化机理等研究,开发了一系列功能完善的水动力-水质-水生生物(水华)耦合的数值模型和软件,具有代表性的如美国Virginia海洋研究所开发的EFDC 模型[34-35],丹麦水资源与环境研究所开发的MIKE 软件[35],荷兰水力学实验室开发的Delft3D 模型软件[36],美国陆军工程兵团与波特兰州立大学联合开发的CE-QUAL-W2 模型[37]等.
基于过程的模型将人们对生物地球化学过程的理解,形成一个数学公式框架,但这些都会产生不可避免的简化,导致对复杂生物地球化学相互作用的描述不完整,导致模型有时可信度低[38].基于创新的数据挖掘和统计技术,数据驱动的机器学习模型已被应用于识别观测数据中的模式[39],并且随着湖泊监测数据的最近激增,机器学习模型已被用作基于过程模型的替代物,用于水华预测[40].早先应用的机器学习模型包括随机森林[41]、支持向量机[42]和人工神经网络模型等[43],可以改进藻类叶绿素的时间和季节性预测,显然可以避免复杂过程模型的编码困难问题.然而,这种模型的缺点是它们缺乏基于过程模型的显式结构中的可解释性和通用性.近年来,过程引导的深度学习模型已经出现,并已应用于水温[44]和水质模拟[45],该模型将明确定义的物理理论结合到机器学习模型的训练中,增强了其可解释性.虽然这种方法取得了可喜的成果,但由于生物地球化学循环中的许多非线性相互作用,以及可指导决策的物理参数的测量过程或质量平衡难以定义,因此很难将其应用于浮游植物生长动力学模型.此外,湖泊水质(如营养盐和叶绿素浓度)观测的稀疏性可能会限制机器学习模型在藻类水华模型中的应用[40].
从水流、到水质、再到水华的数值模拟,需要基于对物理过程的清晰认识和准确表达.已有模型运用研究表明,还没有一个模型能适合各种水体,且从水流、到水质、再到水华的数值模拟精度呈递减趋势,其原因就在于其过程越来越复杂.如三峡水库属于典型的河道型水库,研究发现支流库湾普遍存在分层异重流及振荡波现象[46],水流场的模拟要能精准反映这个水动力特征,才有水质、水华模拟精度的基本保障;此外,目前藻类生态动力学模型将藻类作为固有密度的颗粒进行简化,未考虑藻类的生理行为和环境因素导致藻类密度改变对藻类迁移的影响,这是无法准确模拟藻类垂向分布的重要原因.只有搞清楚这种特殊水动力过程带来的环境生态效应,才能做到水质、水华模拟的准确.
4 进一步研究建议
总体来看,藻类水华引起的生态环境问题已成为世界共同面临的课题,特别在我国,太湖、巢湖、三峡水库等湖库蓝藻水华仍然严重,并呈现突然发生、多点暴发、难以预测的特点[47],水华的预测预报技术和有效的水华防控措施在我国河湖水生态修复与保护工作中仍有重大需求,亟待研究解决.水华生消过程特别是表层水体藻类浓度“骤增”或“突减”的关键机制不明,是导致难以“准确预测预报和精准防控水华”的关键所在.为进一步提升藻类水华的预测预报精度,改进水华的防控措施,建议进一步在如下方面开展更为深入的研究工作.
1)藻类聚散迁移及其影响因素研究:基于不同水生态系统和藻类群体,开展实验以研究藻类颗粒聚散迁移过程与环境因子的关系,重点探究藻类鞭毛、伪空泡、胞外多糖等生理性浮力调节机制和水体溶解性气体、水流扰动等环境浮力调节机制.
2)微流场与藻类生理行为关系研究:开展微流场下藻类迁移聚散行为跟踪观测,研究不同藻类颗粒在微观流场结构下的游泳特征和运动策略,探明微流场对藻类定向行为(如趋光性)、浮力调节、聚散迁移、形态改变(如聚团、离散等)等过程的影响机制.
3)微流场与宏观水动力指标关系研究:研究微流场综合表征指标(如漩涡密度、紊动能、脉冲水流、瞬时浮力频率等),构建宏观流场指标(如水流、流量、水体分层、雷诺数、弗劳德数等)与微流场指标的关系,探究宏观流场指标对微流场特征的定性指示方法.
4)藻类聚散迁移数值模型构建方法研究:综合分析藻类迁移聚散的影响因素,构建藻类迁移聚散数学模型,包括藻类生理性浮力调节模型、环境影响下浮力调节模型、藻类颗粒聚团与分散模型等,修正当前水动力模型中藻类运动迁移项,实现考虑藻类聚散迁移的水华数值模拟.
