灰枣扩展蛋白基因ZjEXPA8的克隆及序列分析
2023-10-11苏比娜肖克来提阿布都卡尤木阿依麦提樊丁宇
靳 娟,苏比娜·肖克来提,阿布都卡尤木·阿依麦提,杨 磊,郝 庆,樊丁宇
(新疆农业科学院园艺作物研究所/农业部新疆地区果树科学观测实验站,乌鲁木齐 830091)
0 引 言
【研究意义】枣(ZiziphusjujubaMill.)为鼠李科(Rhamnaceae)枣属(Ziziphus)植物,是我国特有的经济林树种[1]。枣营养丰富,抗逆性强。新疆是我国最大的优质干枣产区,其中灰枣是最重要的主栽品种,但其自然坐果率较低[2]。研究灰枣的生长发育机理对改良灰枣品种和提高产量具有重要意义。对灰枣花进行转录组学研究,鉴定到一个扩展蛋白基因(Expansin-A8,ZjEXPA8)在花发育不同时期均上调表达[3],研究其序列特征,为分析ZjEXPA8基因在灰枣果实生长发育中的作用和功能奠定基础。【前人研究进展】扩展蛋白是植物中一类重要的细胞壁蛋白,能够打断细胞壁多糖之间之间的非共价键,使细胞壁松弛,促进细胞伸展与分裂[4]。根据结构的不同,扩展蛋白基因家族被分为EXPA(Expansin A)、EXPB(Expansin B)、EXLA(Expansin-like A)和EXLB(Expansin-like B)四个亚家族[5],其中EXPA和EXPB是2个最大的亚家族[6]。蔡兆琴等[7]以甘薯为材料,对扩展蛋白基因IbEXPA4 进行克隆和表达分析,结果表明甘薯IbEXPA4基因的表达能促进块根纵向和横向生长,参与了甘薯块根的生长调节。Bajwa等[8]在当地棉花品种NIAB 846上过表达GhEXPA8基因可以明显促进棉纤维的伸长。赵湾湾等[9]研究发现,随着果实成熟软化程度提高,番木瓜CpEXPA2基因表达量也相应提高,该基因可能参与了番木瓜果实成熟软化过程。扩展蛋白在植物的生长和发育中发挥着重要作用。【本研究切入点】目前,关于灰枣扩展蛋白基因调控果实发育的分子机理研究较少,前期在灰枣转录组学分析中发现ZjEXPA8基因在不同发育时期均上调表达,需该基因与灰枣果实生长发育相关性。【拟解决的关键问题】对ZjEXPA8基因进行克隆,并对其进行生物信息学分析,并采用实时荧光定量PCR技术检测其在灰枣不同发育时期的表达情况,研究ZjEXPA8基因在灰枣果实生长发育中的作用,为灰枣遗传改良和新品种培育提供理论基础和基因资源。
1 材料与方法
1.1 材 料
1.1.1 灰 枣
供试品种为灰枣,采集地点为新疆阿克苏地区阿克苏市试验林场,树龄10 年生,株行距2 m×4 m,树形为自然开心形,枣园水肥投入到位,管理水平较好。分别采集蕾黄期、萼片展平期、花丝萎蔫期、子房膨大期和幼果期5个时期的枣花和幼果样本。样品采集后立即在液氮中冷冻,并保存在-80℃冰箱中备用。
1.1.2 主要试剂
植物多糖多酚总RNA提取试剂盒Quick RNA isolation Kit、反转录试剂盒Fasting RT Kit(With gDNA)、琼脂糖凝胶回收试剂盒、质粒小量提取试剂盒和DL2000 DNA Marker购自北京天根生化科技有限公司;南京诺唯赞公司的2×TaqMaster Mix;pEASY-Blunt克隆载体和DH5α感受态细胞购自北京全式金公司;抗生素卡那霉素(Kan)购自上海生工生物工程有限公司;实时荧光定量PCR SuperReal PreMix Plus(SYBR Green)试剂盒购自TAKARA公司;所用引物均在上海生工公司合成。其他试剂为国产分析纯试剂。
1.2 方 法
1.2.1 总RNA提取和cDNA合成
采用植物多糖多酚总RNA提取试剂盒,按照说明书中的操作流程进行,提取不同发育时期灰枣花的总RNA,提取完成后,使用Nanophotometer-N60核酸检测仪检测所有样品的RNA浓度与纯度,再用1%的琼脂糖凝胶电泳验证RNA的完整性,筛选出质量提取质量较高的RNA样品,保存于-80℃冰箱中。使用Fasting RT Kit(With gDNase)试剂盒将得到的总RNA模板反转录合成灰枣花的cDNA。
1.2.2 灰枣ZjEXPA8基因克隆
根据前期研究发布的灰枣转录组数据库(PRJNA588582)筛选获得目的基因ZjEXPA8,并在NCBI查找获得ZjEXPA8基因的CDS序列,利用Primer Premier 5 软件设计克隆引物,上游引物为5’-GTACCATGGCTAATATTTCTGCATA-3’(ZjEXPA8-F),下游引物为5’- GCGTCGACGAATTGGCTACCCTC-3’(ZjEXPA8-R)。以反转录得到蕾黄期的灰枣花cDNA为模板,利用2×TaqMaster Mix进行PCR扩增。PCR反应体系:cDNA 0.5 μL,ZjEXPA8-F和ZjEXPA8-R引物各1 μL,2×TaqMaster Mix 10 μL,ddH2O 补充反应体系至20 μL。反应条件为95℃ 3 min,95℃ 15 s,57℃ 20 s,72℃ 1 min,35个循环。利用DNA胶回收试剂盒对PCR产物进行回收,并将其连接至pEASY-Blunt载体上,并转化到DH5α感受态细胞中,挑选阳性克隆送至上海生工生物工程有限公司测序。
1.2.3 生物信息学
通过在线软件ProtParam(https://web.expasy.org/protparam/)对灰枣ZjEXPA8编码的氨基酸进行分析,包括蛋白分子量和等电点等;利用ProtScale(https://web.expasy.org/protscale/)分析ZjEXPA8编码蛋白的氨基酸序列的疏水性/亲水性;利用NCBI的Conserved domains database数据库(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)分析ZjEXPA8的结构域;用SOPMA软件(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html)预测ZjEXPA8蛋白的二级结构;利用SWISS-Model(https://swissmodel.expasy.org/interactive)预测ZjEXPA8蛋白的三级结构;利用TMHMM 2.0(http://www.cbs.dtu.dk/services/TMHMM/)预测跨膜域;利用DNAMAN软件比对分析ZjEXPA8序列;用MEGA7.0软件对不同物种的ZjEXPA8基因编码的氨基酸序列以邻接法来构建系统进化树。
1.2.3 实时荧光定量PCR
根据已获得的ZjEXPA8基因序列,设计实时荧光定量PCR引物,上游引物为5’- ATGCCGGATCTAAGCCCTCT -3’(QZjEXPA8-F),下游引物为5’-CATACCCACAAGCACCTCCCAT’ (QZjEXPA8-R)。以枣ZjActin为内参基因,将不同发育时期灰枣花和幼果期的cDNA为模板,采用qTOWER3G荧光定量PCR仪检测ZjEXPA8基因在不同发育时期灰枣花和幼果中的表达水平,用内参基因检测扩增效率一致性。荧光定量反应体系为cDNA 1 μL,QZjEXPA8-F和QZjEXPA8-R引物各0.5 μL,2×SuperReal PreMix Plus 10 μL,ddH2O 补充反应体系至20 μL。反应条件为95℃ 15 min,95℃ 10 s,55℃ 30 s,72℃ 20 s,40个循环,重复3次。实时荧光定量数据采用2-ΔΔCt法进行分析计算。用GraphPad Prism 7.0软件进行数据处理和作图。
2 结果与分析
2.1 灰枣ZjEXPA8基因克隆
研究表明,灰枣ZjEXPA8基因包含一个长度为762 bp的开放阅读框,编码253个氨基酸,以ATG 为起始密码子,TAG为终止密码子。图1,图2
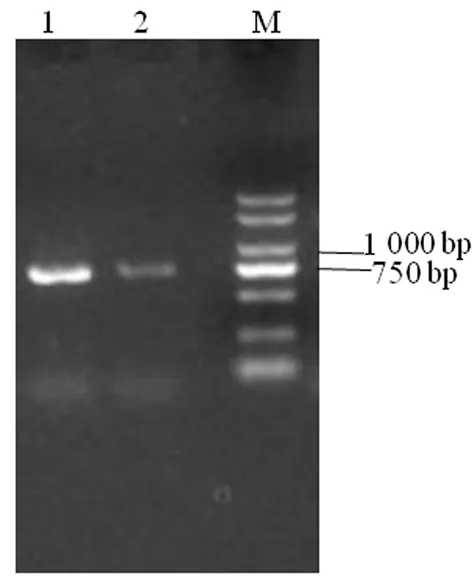
注:M.DL2000 DNA Marker;1, 2: ZjEXPA8
2.2 灰枣ZjEXPA8基因的生物信息学
研究表明,ZjEXPA8蛋白中甘氨酸Gly含量最高,占全部氨基酸的13.8%;而谷氨酸Glu,组氨酸His和甲硫氨酸Met含量相对较少,分别占全部氨基酸的0.8%,1.6%和1.6%。灰枣ZjEXPA8带负电荷氨基酸残基(Asp+Glu)总数为10,带正电荷氨基酸残基(Arg+Lys)总数为13;不稳定系数为30.09,属于稳定蛋白。预测ZjEXPA8基因编码的氨基酸序列的分子式为C1192H1787N327O360S14,分子量为 26.91 kDa,等电点为8.32。表1
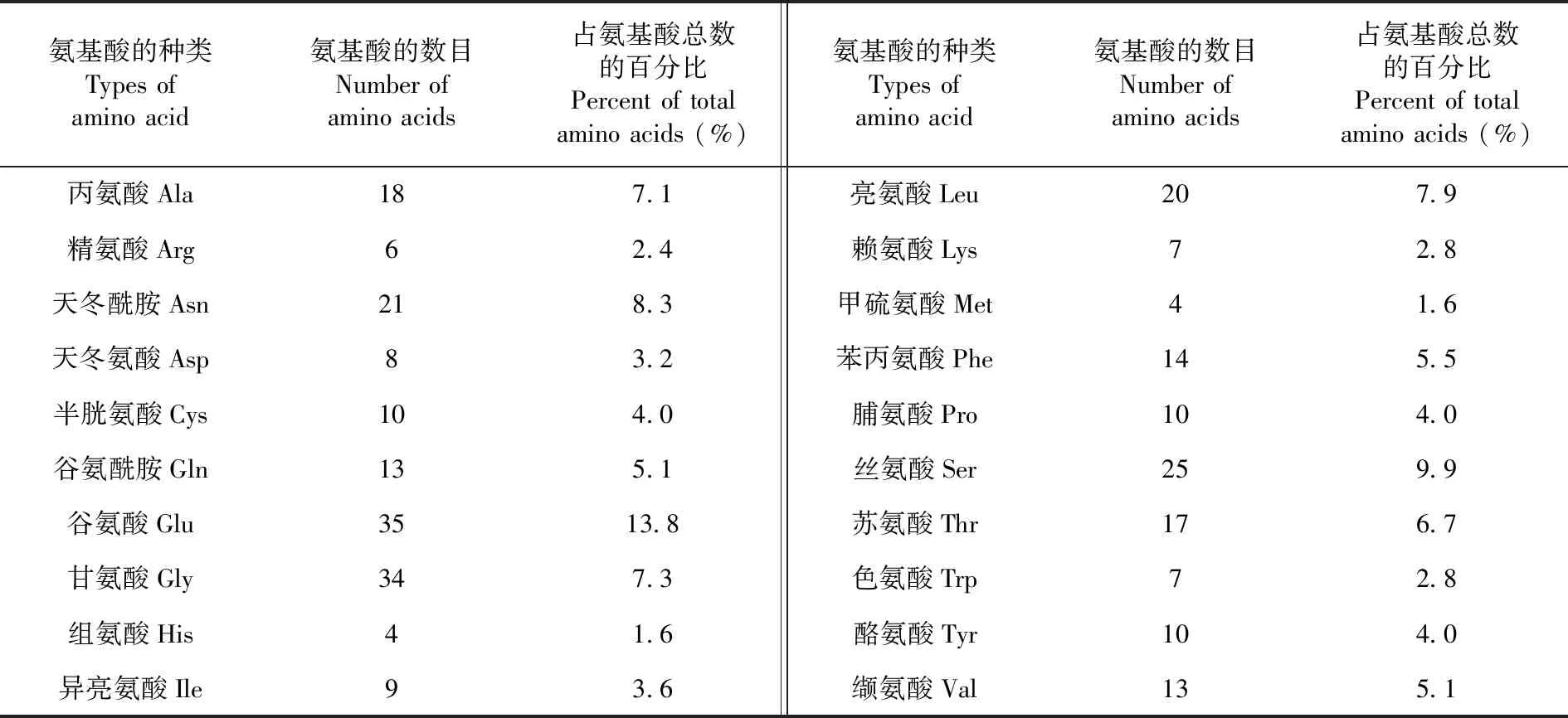
表1 ZjEXPA8编码蛋白的氨基酸组成
灰枣花ZjEXPA8基因编码的氨基酸中,第13位的氨基酸残基的疏水性均最强,为2.933,第207位的氨基酸残基亲水性最强,为-2.322。该蛋白脂肪指数为66.72,总亲水性平均数为-0.107,预测ZjEXPA8属于亲水蛋白。图3
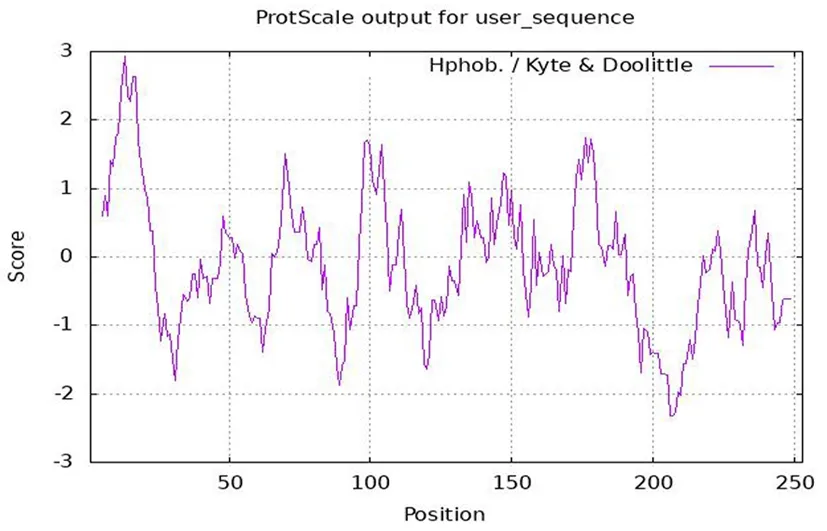
图3 ZjEXPA8编码蛋白亲水/疏水性的预测
ZjEXPA8基因编码的253个氨基酸中,无规则卷曲占比最高,为51.78%,α螺旋占15.81%,延伸链占23.32%,β-转角占9.09%,与SWISS-MODEL三维建模的结果相吻合。该蛋白含有DPBB_1和Pollen_allerg_1 2个保守结构域。该蛋白没有跨膜域。图4~6
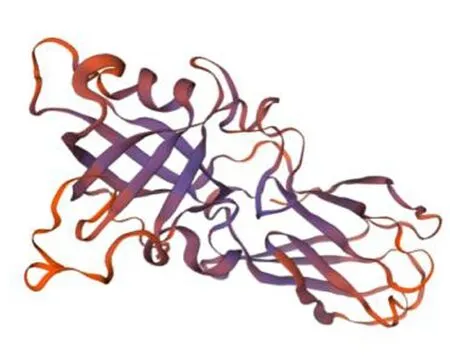
图5 ZjEXPA8蛋白的三级结构预测
图6 ZjEXPA8蛋白质保守结构域预测
2.3 灰枣ZjEXPA8基因的同源序列比对及系统进化树构建
研究表明,灰枣ZjEXPA8基因编码的氨基酸序列与大豆(Glycinemax,XP_003523276.1,GmEXPA8)的相似性最高,为93.13%;与蔓花生(Arachisduranensis,XP_015945084.1,AdEXPA8)、木薯(Manihotesculenta,XP_021617395.1,MeEXPA8)、柱花草(Stylosanthesguianensis,QHQ74416.1,SgEXPA8)、橡胶(Heveabrasiliensis,XP_021684783.1,HbEXPA8)、蒺藜苜蓿(Medicagotruncatula,XP_013461515.1,MtEXPA8)、麻风树(Jatrophacurcas,XP_012091147.1,JcEXPA8)、巨桉(Eucalyptusgrandis,XP_018732353.2,EgEXPA8)、番樱桃(Syzygiumoleosum,XP_030441410.1,SoEXPA8)、阿月浑子(Pistacia vera,XP_031271261.1,PvEXPA8)、甜橙(Citrus sinensis,KAH9702866.1,CsEXPA8)、杨梅(Morellarubra,KAB1217588.1,MrEXPA8)、番木瓜(Caricapapaya,XP_021904225.1,CpEXPA8)和棉花(Gossypiumhirsutum,XP_016686328.1,GhEXPA8)的相似性分别为92.03%、90.76%、90.44%、90.12%、90.04%、89.60%、89.24%、88.93%、88.80%、88.58%、88.40%、87.80%和7.80%。与模式植物拟南芥(Arabidopsisthaliana,NP_181593.1,AtEXPA8)的相似性最低,为78.74%。图7
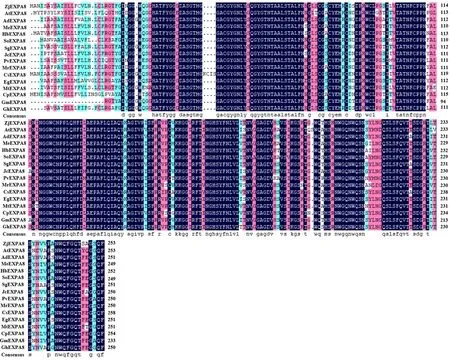
图7 ZjEXPA8的氨基酸序列与其他植物EXPA8氨基酸序列比对
灰枣ZjEXPA8基因与橡胶(Heveabrasiliensis,XP_021684783.1,HbEXPA8)和拟南芥(Arabidopsisthaliana,NP_181593.1,AtEXPA8)的亲缘关系最近。图8
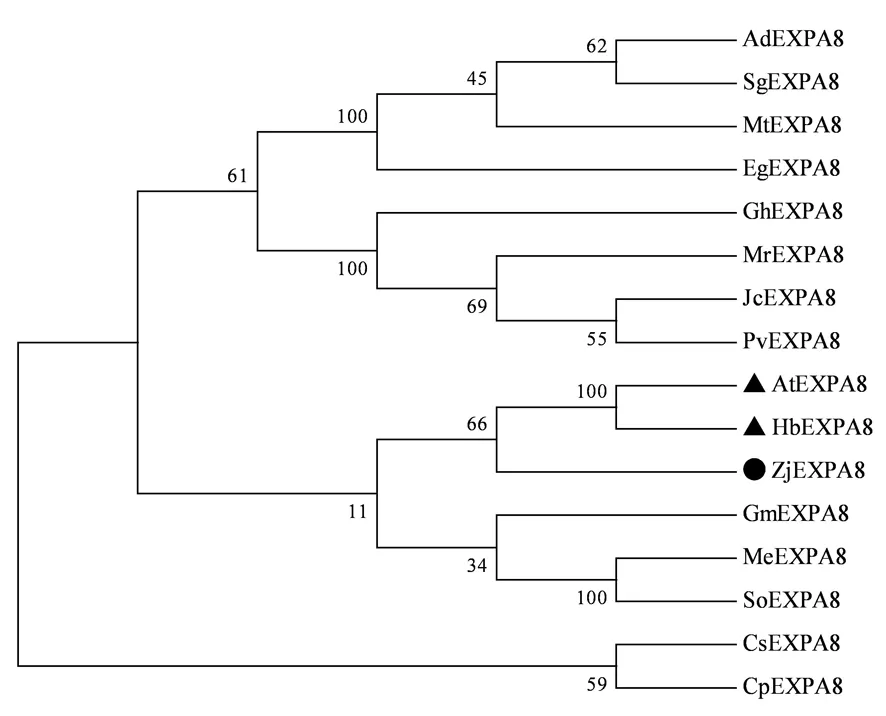
图8 ZjEXPA8蛋白序列的系统进化
2.4 灰枣ZjEXPA8基因表达
研究表明,灰枣ZjEXPA8基因在枣花发育不同时期(蕾黄期、萼片展平期、花丝萎蔫期、子房膨大期)和幼果期均有表达,蕾黄期表达量最高,萼片展平期次之,该基因可能主要在蕾黄期和萼片展平期发挥作用。图9
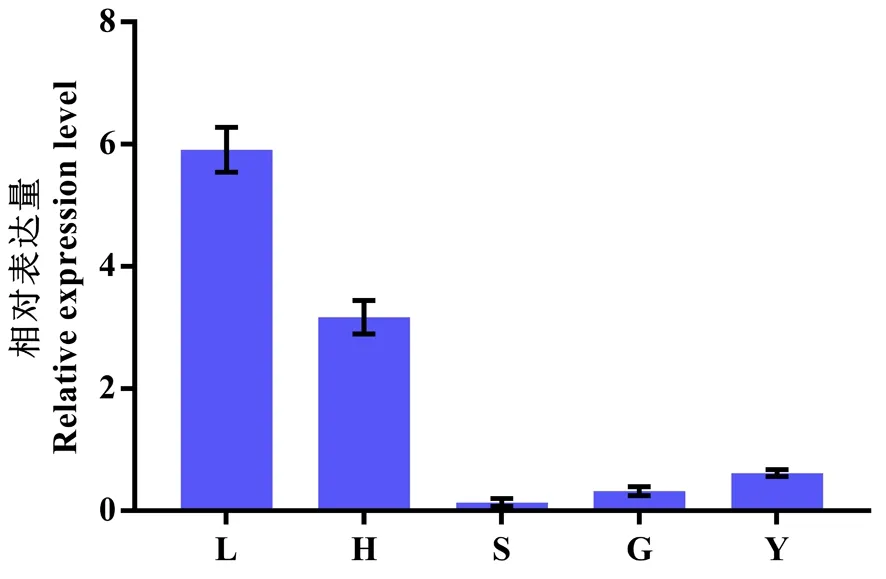
注:L.蕾黄期;H.萼片展平期;S.花丝萎蔫期;G.子房膨大期;Y.幼果期
3 讨 论
3.1枣(ZiziphusjujubaMill.)是我国重要栽培果树和生态树种[10]。目前在全疆59个县市区广泛种植。2019年底[11]新疆红枣种植面积44.52 ×104hm2(含兵团10.04×104hm2)、产量372.77×104t(含兵团200.33×104t)。灰枣是新疆栽培面积最大的主栽品种,其花量大,但落花落果严重,自然坐果率极低[2]。
3.2ZjEXPA8基因在花发育不同时期均上调表达,其可能在灰枣果实生长调控中发挥重要作用[3]。研究利用RT-PCR方法克隆获得灰枣扩展蛋白ZjEXPA8基因,该基因全长为762 bp,编码253个氨基酸,蛋白分子量为 26.91 kDa,不稳定系数为30.09,总亲水性平均数为-0.107,属于稳定亲水蛋白。ZjEXPA8蛋白的结构进行分析含有DPBB_1和Pollen_allerg_1两个保守结构域,与甘薯IbEXPA4蛋白的结构相似[7],其属于EXPA 亚家族。同源序列比对结果显示,ZjEXPA8蛋白与大豆GmEXPA8的相似度最高,达93.13%,蔓花生AdEXPA8次之,相似度达92.03%。系统进化树分析表明,ZjEXPA8基因与橡胶HbEXPA8和拟南芥AtEXPA8基因的亲缘关系最近。
3.3扩展蛋白在植物的生长和发育过程中扮演着重要角色[12]。扩展蛋白参与种子萌发[13]、叶片生长[14]、根系发育[15]、花芽发育[16]、果实成熟[17]等多个生物学过程。研究中,灰枣ZjEXPA8基因在枣花发育不同时期均有表达,蕾黄期表达量最高,萼片展平期次之,该基因可能主要在蕾黄期和萼片展平期发挥作用。研究探明了灰枣ZjEXPA8的基因序列和结构特征,但是该基因在灰枣果实发育过程中的调控作用和功能还有待于进一步的深入研究。
4 结 论
克隆获得了灰枣扩展蛋白ZjEXPA8基因,开放阅读框为762 bp,编码253个氨基酸,蛋白分子量为 26.91 kDa。ZjEXPA8基因编码的蛋白与大豆GmEXPA8的同源性最高,亲缘关系上与橡胶HbEXPA8和拟南芥AtEXPA8最近。ZjEXPA8基因在灰枣花蕾黄期表达量最高。
