31份小麦材料中抗旱基因的KASP检测
2023-10-11孔淑鑫宋国琦李玉莲张淑娟张荣志李吉虎樊庆琦李根英
李 玮,孔淑鑫,宋国琦,李玉莲,张淑娟,张荣志,高 洁,李吉虎,樊庆琦,李根英
(1.山东省农业科学院作物研究所/小麦玉米国家工程研究中心/农业农村部黄淮北部小麦生物学与遗传育种重点实验室/山东省小麦技术创新中心,山东济南 250100; 2.山东农业工程学院农业科技学院,山东德州 251100)
干旱是影响小麦产量的重要因素之一。选育抗旱节水品种是保障旱地小麦稳产最经济、有效的方法。传统小麦抗旱育种是在种质创新的基础上,通过杂交后代的抗旱鉴定和产量筛选培育抗旱品种,通常采用高产×抗旱或抗旱×抗旱的组合模式,目前仍是品种选育的主流方法。但传统小麦抗旱育种方法周期较长、育种效率低,因此缩短育种年限、加快育种进程已成为目前小麦新品种选育的客观要求。
随着小麦基因组的测序完成,越来越多与小麦抗旱相关的基因被定位和克隆,为小麦抗旱分子标记育种奠定了基础。鞠丽萍等[1]根据不同抗旱性小麦品种中铁结合蛋白基因(TaFer-A1)的序列差异开发了分子标记FerA1-intrl,并进行了有效性验证,发现该标记FerA1-intr1可用于小麦抗旱性的鉴定和筛选,也表明TaFer-A1基因与小麦抗旱性有关。Shoaib等[2]利用16个小麦抗旱基因的KASP标记对153份小麦品种进行了检测,发现优异等位基因与产量性状显著相关。于铭等[3]通过分析吡咯啉-5-羧酸合成酶基因(TaP5CS)在不同抗旱性小麦品种中的序列差异,发现第5内含子上存在SNP,据此开发了分子标记TaP5CS-1A-CAPS,可用于小麦萌发期抗旱性的筛选与鉴定。Mao等[4]研究发现,TaPYL1-1B基因启动子区的20 bp插入与抗旱性显著关联,20 bp的插入中含有的MYB元件可以增强TaPYL1-1B的表达,从而提高小麦的抗旱性。以上研究均为小麦抗旱品种选育提供了有效标记。随着大量与抗旱相关分子标记的开发与应用,分子标记在抗旱小麦品种辅助选育过程中已成为重要的筛选方法。
为提高抗旱小麦育种过程中亲本材料选择的目的性和准确性,本研究利用29个KASP标记,对收集到的16份抗旱小麦品种和15份高产小麦品种进行检测,明确这些材料中抗旱基因的分布情况,以期为分子标记辅助选择创制聚合优异基因的新种质提供依据。
1 材料与方法
1.1 材料
供试小麦品种共31份,其中抗旱小麦品种16份,包括济麦379、山农27、和尚头、青麦6号、临麦9号、济麦262、济麦60、存麦1号、晋麦47、晋麦92、晋麦101、青麦7号、鲁麦21、品育8161、鹤麦801和洛旱7号;高产小麦品种15份,包括MFW1、鲁研128、周麦27、山农20、烟农19、烟农999、中麦578、济麦5198、济麦38、山农30、济麦22、山农29、百农207、鲁原502和矮抗58,均由本课题组收集保存。
1.2 DNA提取和分子标记检测
取5粒均匀饱满的小麦种子置于一次性培养皿中,加水浸泡24 h左右,待种子露白后选择生长一致的3粒种子种植在装有育苗基质的塑料盆中,在人工气候室(23 ℃光照/18 ℃黑暗,16 h光照/18 h黑暗)继续培养7 d后,每株取3个叶片,按照高 洁等[5]的方法提取DNA。
1.3 分子标记类型转换及KASP检测
利用已报道的29个标记[2-4,6-12](表1)对供试材料进行检测,其中18个标记类型为KASP,所用引物序列参考相应文献。对其他11个标记进行KASP转换,利用文献中报道的原始标记引物序列,通过WheatOmics[13]网站进行序列比对,获得参考基因组的分子标记扩增序列,按照文献描述的SNP或Indel信息修改参考基因组序列,获得等位变异序列。用DNAMAN对2个序列进行比对,设计KASP标记引物。
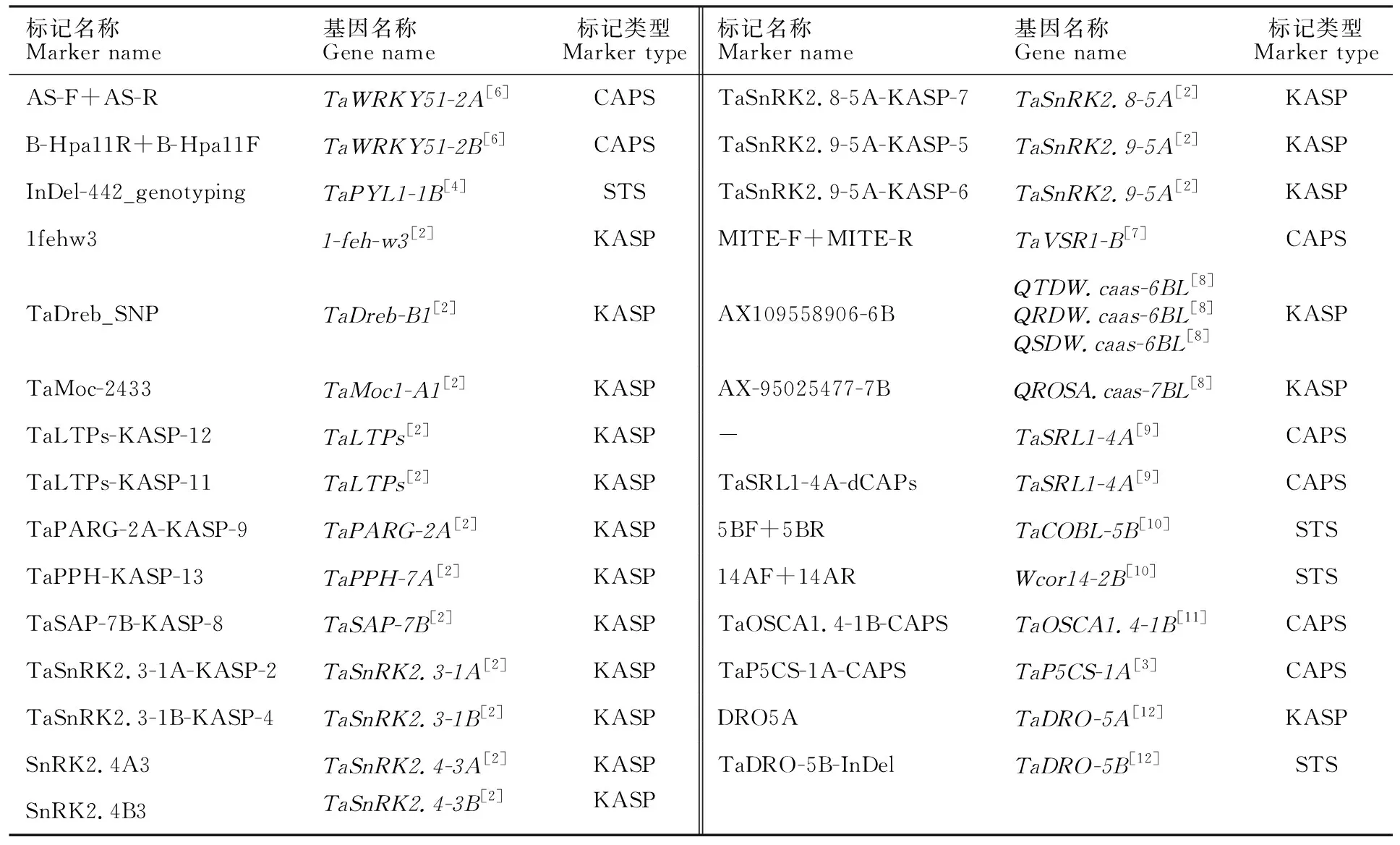
表1 已报道的29个标记的类型Table 1 Types of 29 markers reported
进一步对KASP标记引物进行检测,选取分型效果好的引物组合进行检测。试验所需的KSAP标记引物均由北京擎科生物科技有限公司合成,共有三组混合引物,每一组混合引物都含有两条末端碱基不同的等位基因正向引物(100 μmol·L-1)各12 μL、一条共同的反向引物(100 μmol·L-1)30 μL以及ddH2O 46 μL。PCR反应体系为5 μL,包括2.0 μL DNA ( 50 ng·μL-1),2.5 μL KASP Master Mix ( LGC Genomics, Hoddeston, UK),0.07 μL混合引物和0.43 μL ddH2O。为避免对试验结果的误判,设置空白对照,其DNA模板用ddH2O代替。PCR反应在ABI veriti PCR仪上进行,PCR反应程序:95 ℃预变性15 min;95 ℃变性20 s,65 ℃退火和延伸25 s,10个循环;57 ℃退火和延伸60 s,30个循环。反应结束后利用PHERA star多功能酶标仪读取荧光数据﹐通过Kluster Caller软件(LGC Genomics, Hoddeston, UK)生成基因分型图。
2 结果与分析
2.1 分子标记类型的转换结果
由于部分基因(表2第1列)的原始标记为STS或CAPS,为了提高检测效率,需将其转化成KASP标记。利用中国春参考基因组通过引物的序列比对,获得引物的物理位置以及11个分子标记扩增产物的原始参考基因组序列,根据SNP或Indel位点设计11对KASP引物,并对31份小麦品种的DNA进行检测,表明这些STS或CAPS标记已成功转化为KASP标记(表2)。其中,TaSRL1-4A基因的2个SNP转化为2个KASP标记。
2.2 KASP标记检测结果
利用29个KASP标记对31份小麦品种进行检测,部分标记的检测结果如图1所示。根据31份小麦品种的检测结果,可将KASP标记分成四类,第一类是优异等位基因占比较高的标记(图2),包括DRO5A(90.32%)、TaDreb_SNP(77.42%)、TaPPH-KASP-13(83.87%)、PYL1B(83.87%)、VSR1-2BMITE(87.10%)、1fehw3(74.19%)、SnRK2.4B3(90.32%)、TaSnRK2.8-5A-KASP-7(93.55%)、TaLTPs-KASP-11+TaLTPs-KASP-12(87.10%)和Wcor142B(77.42%),表明这些标记检测到的优异等位基因在供试材料中利用程度较高;第二类是两种等位基因占比相当的标记,包括P5CS1A(54.84%)、SRL14A929(54.84%)、SRL14A96(54.84%)和TaSnRK2.3-1A-KASP-2(41.94%),表明这些标记检测到的优异等位基因在供试材料中利用程度中等;第三类是优异等位基因占比较低的标记,包括WRKY51-2B(9.68%)、WRKY51-2A(32.26%)、TaSnRK2.3-1B-KASP-4(22.58%)、SnRK2.4A3(3.23%)、TaSnRK2.9-5A-KASP-5(23.33%)、TaSnRK2.9-5A-KASP-6(23.33%)、COBL5B(22.58%)、AX109558906-6B(29.03%)和OSCA1.4(12.90%),表明这些标记检测到的优异等位基因在供试材料中利用程度较低;第四类是在所检测材料中只检测到一种等位基因的标记,包括TaSAP-7B-KASP-8、AX-95025477-7B、TaMoc-2433、DRO5B和TaPARG-2A-KASP-9,其中AX-95025477-7B和TaMoc-2433检测到的基因为非优异等位基因,其余为优异等位基因的功能标记。
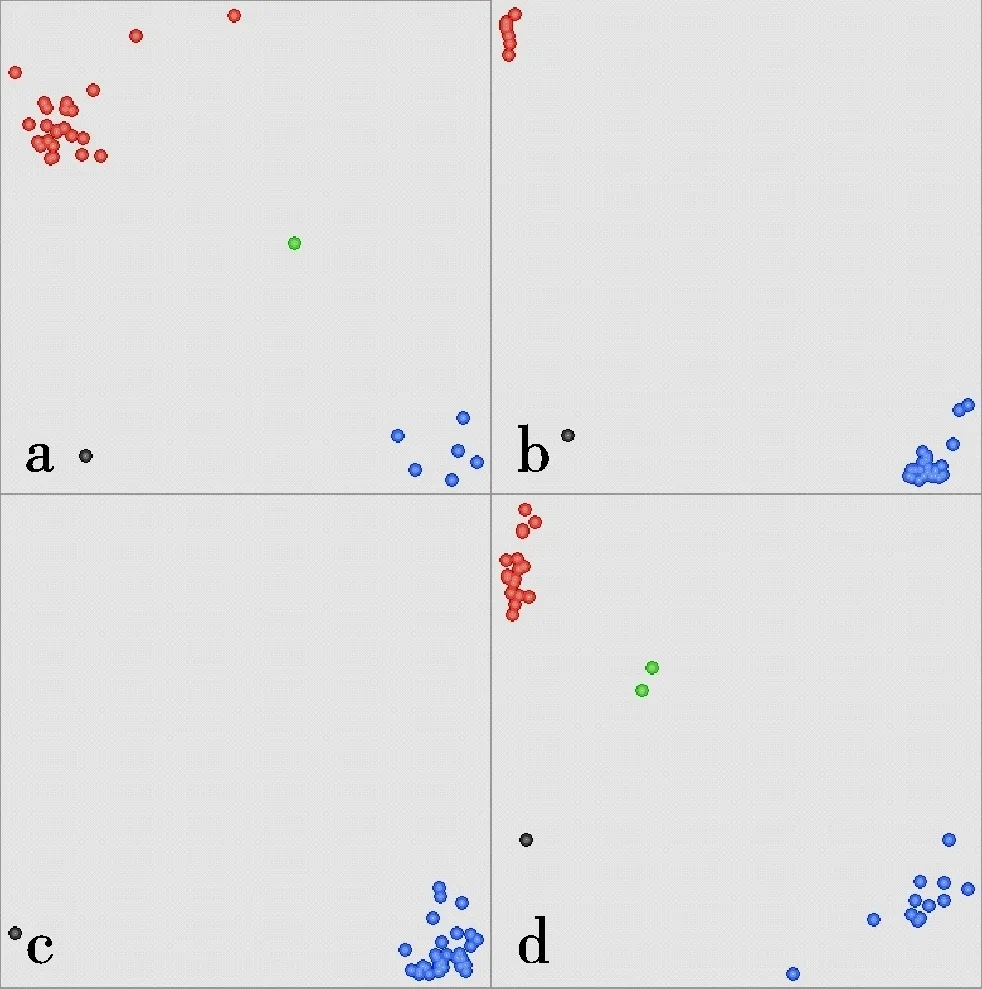
a:WRKY51-2A;b:TaDreb_SNP;c:AX-95025477-7B,d:P5CS1A。红色点为Hex等位基因;蓝色点为Fam等位基因;绿色点为杂合型;黑色点为阴性对照。a: WRKY51-2A; b: TaDreb_SNP; c: AX-95025477-7B; d: P5CS1A. Red dots represent Hex allele; Blue dots represent Fam allele; Green dots represent heterozygous; Black dots represent no template control.图1 供试材料中部分KASP标记检测分型图(散点图)Fig.1 Detection of the tested wheat materials by KASP markers(scatter plots)

图中白色、浅灰色、黑色和深灰色柱子分别代表Fam等位基因、Hex等位基因、杂合基因型和未知基因型。The white, light gray, black and dark gray bars represent Fam, Hex, heterozygous and unknown allele, respectively.图2 29个KASP标记所检测到的等位基因分布频率Fig.2 Allele frequency detected by 29 KASP markers
2.3 双标记对抗旱相关基因的检测结果
分别用2个KASP标记对3个抗旱基因TaLTPs、TaSRL1-4A和TaSnRK2.9-5A进行检测,TaLTPs基因的2个标记TaLTPs-KASP-11和TaLTPs-KASP-12是根据TaLTPs启动子区的2个SNP开发而来,2个SNP共形成3种单倍型[14],本研究中共检测到HapⅡ和HapⅢ两种单倍型,其中HapⅢ是主要单倍型,与株高相关。TaSRL1-4A基因虽然在启动子和编码区共有3个SNP[9],但仅有2种单倍型Hap-4A-1和Hap-4A-2,2个标记SRL14A929和SRL14A96的检测结果完全一致,2种单倍型各占50%左右,Hap-4A-2单倍型与高千粒重相关。TaSnRK2.9-5A基因的2个标记TaSnRK2.9-5A-KASP-5和TaSnRK2.9-5A-KASP-6形成4种单倍型[15],本研究共检测到Hap-5A-1、Hap-5A-3和Hap-5A-4三种单倍型,2个标记的检测结果完全一致。其中Hap-5A-1与高千粒重相关,Hap-5A-4与高穗粒数相关,分别占比23.33%和3.23%。
2.4 高产小麦品种和抗旱小麦品种之间优异等位基因的数目差异
将供试小麦品种分成高产组和抗旱组分别进行统计,发现大部分标记所检测到的优异等位基因数目在组间差异较小(小于等于3),差异优异等位基因数目超过3个的标记包括PYL1B、Wcor142B、SRL14A96、SRL14A929、TaSnRK2.3-1A-KASP-2和AX109558906-6B(图3),其中仅Wcor142B在高产组中检测到的优异等位基因数目比抗旱组多,其余均是抗旱组较多,表明这些标记在抗旱育种中值得关注。此外,WRKY51-2B和SnRK2.4A3标记所检测到的优异等位基因数目虽然在组间差异较小,但仅在抗旱组中能检测到优异等位基因。携带TaWRKY51-2B位点优异等位基因 Hap-2B-Ⅱ(与根长相关[6])的品种有晋麦47、晋麦92和晋麦101,携带TaSnRK2.4-3A位点优异等位基因T allele(与高千粒重相关[16])的品种仅有济麦379。
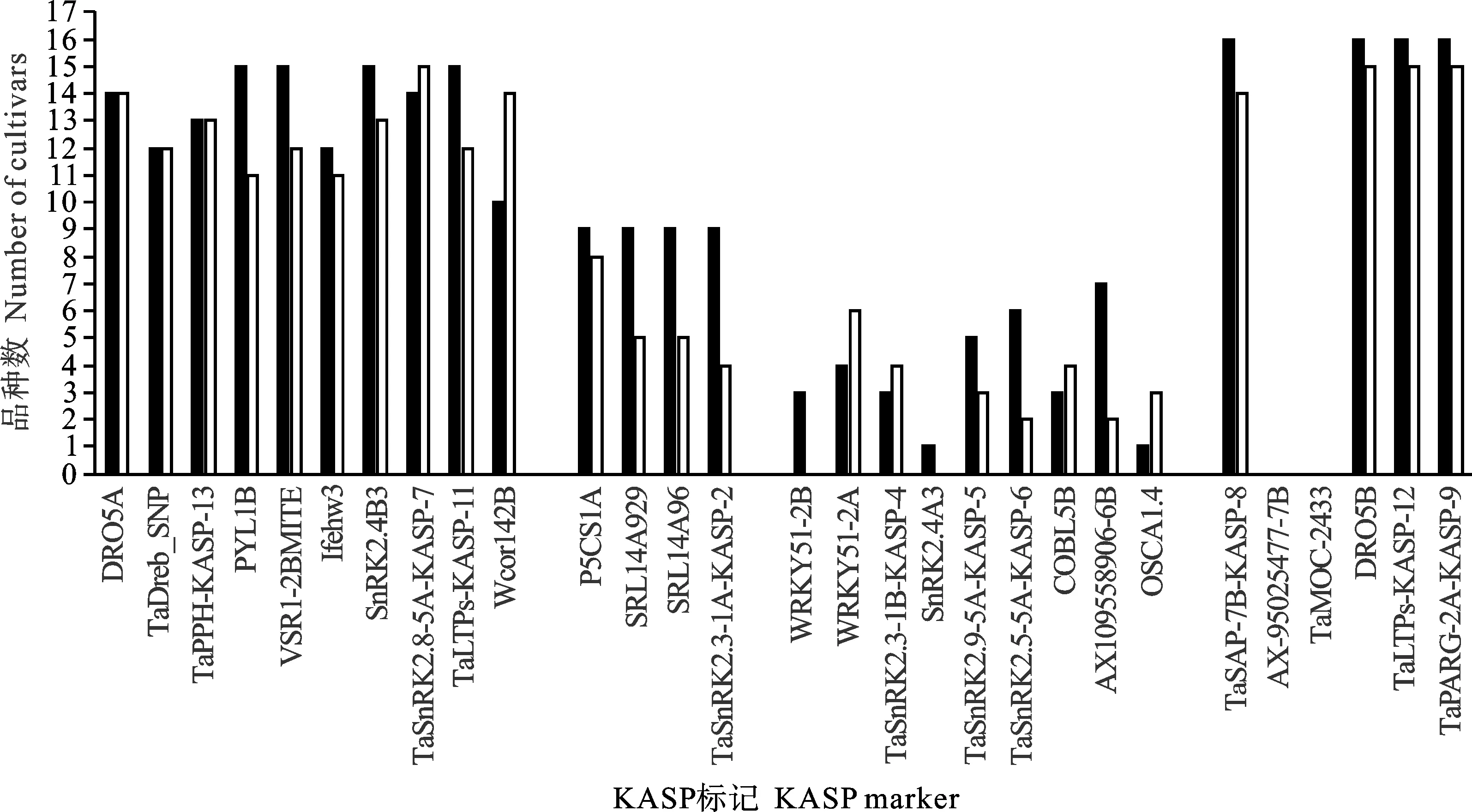
黑色和白色分别代表抗旱小麦品种和高产小麦品种。Black and white represent drought resistant wheat cultivars and high-yield wheat cultivars, respectively.图3 抗旱小麦品种和高产小麦品种优异等位基因数目差异比较Fig.3 Comparison of allele number between drought resistant wheat cultivars and high-yield wheat cultivars
3 讨论
抗旱育种在我国开展较早,通过常规育种方法已选育出大量抗旱节水小麦品种。进入21世纪以来,抗旱小麦新品种选育进展缓慢,新审定的旱地品种虽然在产量和品质上有所提高,但在生产上抗旱性的表现并不佳。随着现代生物技术的发展,越来越多的功能基因或QTL被挖掘,为分子标记辅助选择抗旱育种奠定了基础[17-18]。明确亲本的等位基因组成,根据基因互补原则,选择抗旱基因供体亲本和高产受体亲本进行杂交组配,结合农艺性状选择和分子标记辅助选择的方法可提高育种效率。本研究利用29个KASP标记对收集的16个抗旱小麦品种和15个高产小麦品种进行了分子检测,通过对比发现,Wcor142B标记在高产小麦品种中检测到的优异等位基因占比较高,而PYL1B、SRL14A96、SRL14A929、TaSnRK2.3-1A-KASP-2、AX109558906-6B标记在抗旱小麦品种中检测到的优异等位基因占比较高。Mao等[4]研究发现,TaPYL1-1B位点的优异等位基因与小麦的高ABA(脱落酸)敏感性、高光合能力和高水分利用效率相关。Zhuang等[9]研究发现,TaSRL1-4A位点的优异等位基因Hap-4A-2与浅根、矮秆和高千粒重相关。Miao等[19]研究发现,TaSnRK2.3-1A位点的优异等位基因Hap-1A-1与矮秆和高千粒重相关。Yang等[8]研究发现,QTDW.caas-6BL、QRDW.caas-6BL和QSDW.caas-6BL位点的优异等位基因与较高的根干重、根表面积和地上部干重相关。本研究中TaPYL1-1B、TaSRL1-4A、TaSnRK2.3-1A以及QTDW.caas-6BL/QRDW.caas-6BL/QSDW.caas-6BL的优异等位基因占比分别为83.87%、45.16%、41.94%和29.03%。说明上述位点的优异等位基因在育种中已得到利用,但是利用率还有待提升。本研究仅在抗旱品种中检测到优异等位基因TaWRKY51-2B和TaSnRK2.4-3A,并筛选到TaWRKY51-2B的供体亲本为晋麦47、晋麦92、晋麦101,TaSnRK2.4-3A的供体亲本为济麦379。济麦379是新审定的节水多抗优质早熟小麦新品种,携带TaSnRK2.4-3A位点优异等位基因;携带TaWRKY51-2B位点的优异等位基因的三个品种中,晋麦101与济麦379的优异等位基因互补性更好。因此,开展以晋麦101为优异等位基因供体,以济麦379为受体的遗传改良工作,在保持济麦379优异性状的基础上利用分子育种技术进一步聚合抗旱和高产优异等位基因,有望培育出更好的抗旱品种。
