红松自由授粉子代测定及优良家系选择*
2023-10-09潘凤刚孙佰飞张含国潘志刚高艳容
潘凤刚 孙佰飞 张含国 潘志刚 高艳容
(1.苇河林业局青山林木种子园,黑龙江 尚志 150623;2.东北林业大学,黑龙江 哈尔滨 150040;3.方正县腰岭子林场,黑龙江 哈尔滨 150812)
红松(Pinus koraiensis)为松科(Pinaceae)松属(Pinus)乔木,是寒温带针阔叶混交林的主要建群树种,其树干通直、材质良好、出材率高,为中国东北地区珍贵用材树种,主要分布于长白山、张广才岭、老爷岭、完达山和小兴安岭[1-2]。树干圆满且高大,加工出材率高,是生产优质大径材的首选树种[3]。红松成材时间较长,天然林红松幼龄材与成熟材的界限为35 年,人工林红松幼龄材与成熟材的界限为15 年,大径级用材林平均培育年限需70 年左右[4-5]。由于红松生长速度较慢,优良材料选育迟缓,严重制约了红松用材林培育。对红松生长性状的遗传改良是缩短育种周期、提高红松生产力水平的有效途径。黑龙江省苇河林业局1970 年代开始红松一代种子园的营建工作,1980 年代开展红松二代种子园筹备工作,1988 年营建红松子代测定林,为种子园提供遗传基因资源。随着林木遗传育种研究的深入,建立高世代种子园已成为发展的需要[6-8]。在生长成熟之前,子代测定林的树龄越大,选择结果越准确,生长性状的遗传变异是筛选优良群体的重要内容[9-10]。
本研究以黑龙江省苇河林业局红松子代测定林117 个红松自由授粉子代家系为材料,对其27 年生时树高、胸径、材积性状进行家系间差异性比较与遗传变异分析,基于材积性状育种值估计,筛选出优良家系,为红松初级种子园的去劣疏伐提供理论依据和参考数据,旨在为红松高世代种子园营建提供基础材料。
1 材料与方法
1.1 试验林概况
试验林位于黑龙江省苇河林业局青山种子园,当地海拔为300 m,年降水量为666.1 mm,年日照时数为2 552.3 h,年均温2.3℃。年蒸发量1 084.4 mm,>5℃积温为2 753.2℃,7 月均温21.6℃。试验林采用完全随机区组设计,单行小区8 株,5 次重复,株行距1.5 m×2.0 m。
1.2 试验材料
试验材料为1988 年黑龙江省苇河林业局营建的红松自由授粉子代家系测定林,共有117 个家系(包括1 个对照,种子来源于当地生产对照,其余116 个家系来源为黑龙江省鹤北优树嫁接营建的无性系种子园优良单株)。
1.3 数据分析方法
在林龄27 年时对红松自由授粉子代家系树高、胸径性状进行测量调查。
采用实验形数法计算立木材积:
式中,g1.3为胸高断面积;H为树高;fε为试验形数(红松为0.42)。
变异系数
式中,SD为性状观测值标准差;为性状群体均值。
方差分析采用以下线性模型:
式中,Yijk为第i个区组第j个家系第k观测值;Bi为区组效应;μ 为家系平均值;Fj为家系效应;BFij为家系与区组互作效应;eijk为随机误差。
家系遗传力
单株遗传力
式中,σf2为家系的方差分量;σfb2为家系与区组互作方差分量;σe2为环境方差分量;n为小区内株数;b为区组数。
采用最佳线性无偏估计(BLUP)法进行育种值估算[11-12]遗传增益
式中,xi为入选家系平均值;x为性状群体均值;hf2为家系遗传力。
以上数据分析主要采用SPSS26.0、R 4.1.2 和ASReml 程序包处理与分析。
2 结果与分析
2.1 红松自由授粉家系生长性状描述性统计与变异分析
对红松自由授粉家系树高、胸径和材积性状进行描述性统计分析,结果(表1)显示:树高、胸径、材积平均值分别为11.096 m、16.725 cm、0.054 1 m3,树高变异幅度为6.8~19.9 m,胸径变异幅度为10.56~26.53 cm,材积变异幅度为0.007 65~0.153 34 m3,树高、胸径、材积变异系数分别为11.82%、21.01%、43.99%,其中材积变异系数最大,胸径变异系数其次,树高变异系数最小。
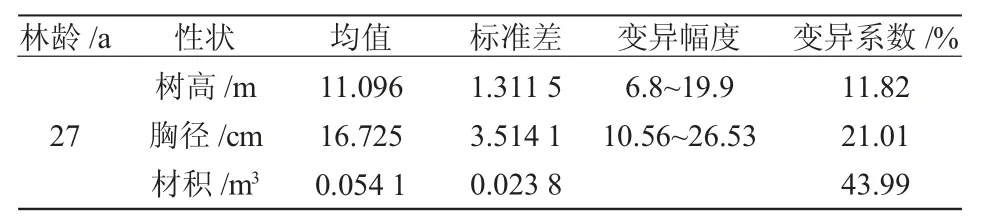
表1 红松自由授粉家系生长性状描述性统计
对各家系红松自由授粉家系树高、胸径和材积性状进行变异分析,结果(表2)显示:树高变异系数最大的家系为072,达23.07%;变异系数最小的家系为086,达5.28%;CK 变异系数达11.83%,变异系数较大的前10 名家系变异系数平均值为16.81%;变异系数居中的10 名家系变异系数平均值为9.92%;变异系数较小的后10 名家系变异系数平均值为6.31%。胸径变异系数最大的家系为049,达28.34%;变异系数最小的家系为166,达9.11%;CK 变异系数达21.27%,变异系数较大的前10 名家系变异系数平均值为26.40%;变异系数居中的10 名家系变异系数平均值为20.69%;变异系数较小的后10 名家系变异系数平均值为12.16%。材积变异系数最大的家系为125,达57.92%;变异系数最小的家系为166,达21.65%;CK 变异系数达44.08%,变异系数较大的前10 名家系变异系数平均值为54.55%;变异系数居中的10 名家系变异系数平均值为43.19%;变异系数较小的10 名家系变异系数平均值为26.33%。
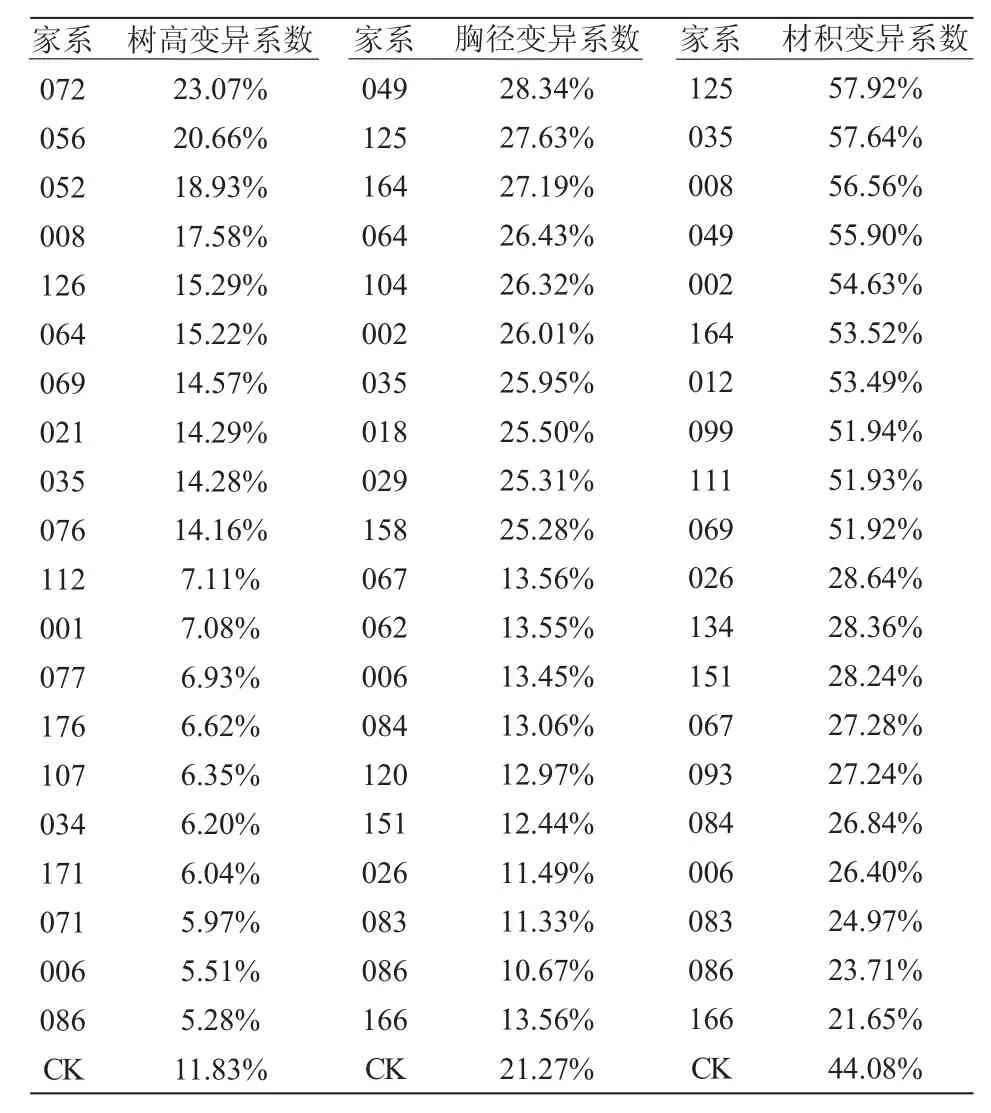
表2 红松自由授粉家系家系生长性状变异分析
2.2 红松自由授粉家系生长性状方差分析与多重比较
为克服环境影响,将调查数据均除以该区CK值后进行方差分析,对红松自由授粉家系树高、胸径和材积性状进行方差分析的结果(表3)显示:各性状家系间差异均达到极显著水平。引起家系间变异的主要因素,即家系效应、区组效应、家系×区组效应,各自的方差分量在不同性状中所占比例不同。在各性状中,家系效应方差分量占比均最高,反映出树高、胸径和材积性状在家系间的差异主要来源于遗传因素。3 个性状中树高性状变异相对较小,表型变异与遗传变异系数分别为0.110 和0.049,胸径性状变异相对居中,表型变异与遗传变异系数分别为0.206 和0.047,材积性状变异相对较大,表型变异和遗传变异系数分别为0.419 和0.106。材积性状表型变异和遗传变异系数分别为树高性状的3.81 倍和2.16 倍,为胸径性状的2.03 倍和2.26 倍。树高、胸径、材积性状的家系遗传力分别为0.857、0.658、0.695。单株遗传力表现与家系遗传表现相近,树高、胸径、材积性状的单株遗传力分别为0.683、0.203、0.243

表3 红松自由授粉家系生长性状方差分析
对红松自由授粉家系树高、胸径和材积性状Duncan 多重比较结果(表4)显示:群体树高均值11.096 m 高于CK 树高均值10.97 m,其中树高均值大于CK 的家系有162、006、197 等65 个家系,65个家系树高均值为11.558m,高于CK 树高均值4.83%;树高均值排名前10 的家系之间差异不显著,10 个家系树高均值为12.376m,高出群体树高均值10.34%,高于CK 树高均值11.36%;其中树高均值最大的家系为162,达12.653m,高于CK 树高均值13.30%。群体胸径均值16.725 cm 低于CK 胸径均值16.774 cm,其中胸径均值大于CK 的家系有034、122、166 等54 个家系,54 个家系胸径均值为17.683 cm,高于CK胸径均值5.14%,胸径均值排名前10 的家系之间差异不显著,10 个家系胸径均值为18.936 cm,高出群体胸径均值11.68%,高于CK 胸径均值11.42%,其中胸径均值最大的家系为034,达20.513 cm,高于CK 胸径均值18.23%。群体材积均值0.054 1 m3高于CK 材积均值0.053 8 m3,其中材积均值大于CK的家系有034、006、122 等57 个家系,57 个家系材积均值为0.060 3 m3,高于CK 材积均值10.78%,材积均值排名前10 的家系中,034、006、122 家系与其他家系间差异显著,10 个家系材积均值为0.0702m3,高出群体均值0.022.91m3,高于CK材积均值23.36%,材积均值最大的家系为034,达0.088 9 m3,高于CK材积均值39.48%。

表4 红松自由授粉家系生长性状Duncan 多重比较
2.3 红松自由授粉家系生长性状育种值分析
育种值的估算在常规育种过程中具有重要意义[13]。对红松自由授粉家系树高、胸径和材积性状育种值进行分析的结果(表5)显示:树高育种值排名在CK 之前的家系有162、006、197 等67 个家系,其中排名第1 的家系为162,排名最后的为023;胸径育种值排名在CK 之前的家系有034、122、062等52 个家系,其中排名第1 的为034,排名最后的为074;材积育种值排名在CK 之前的家系有034、006、036 等57 个家系,其中排名第1 的为034,排名最后的为074。各性状家系育种值排名靠前的家系其性状均值也较大,育种值排名与均值排名间存在不同,排名上下波动不明显。
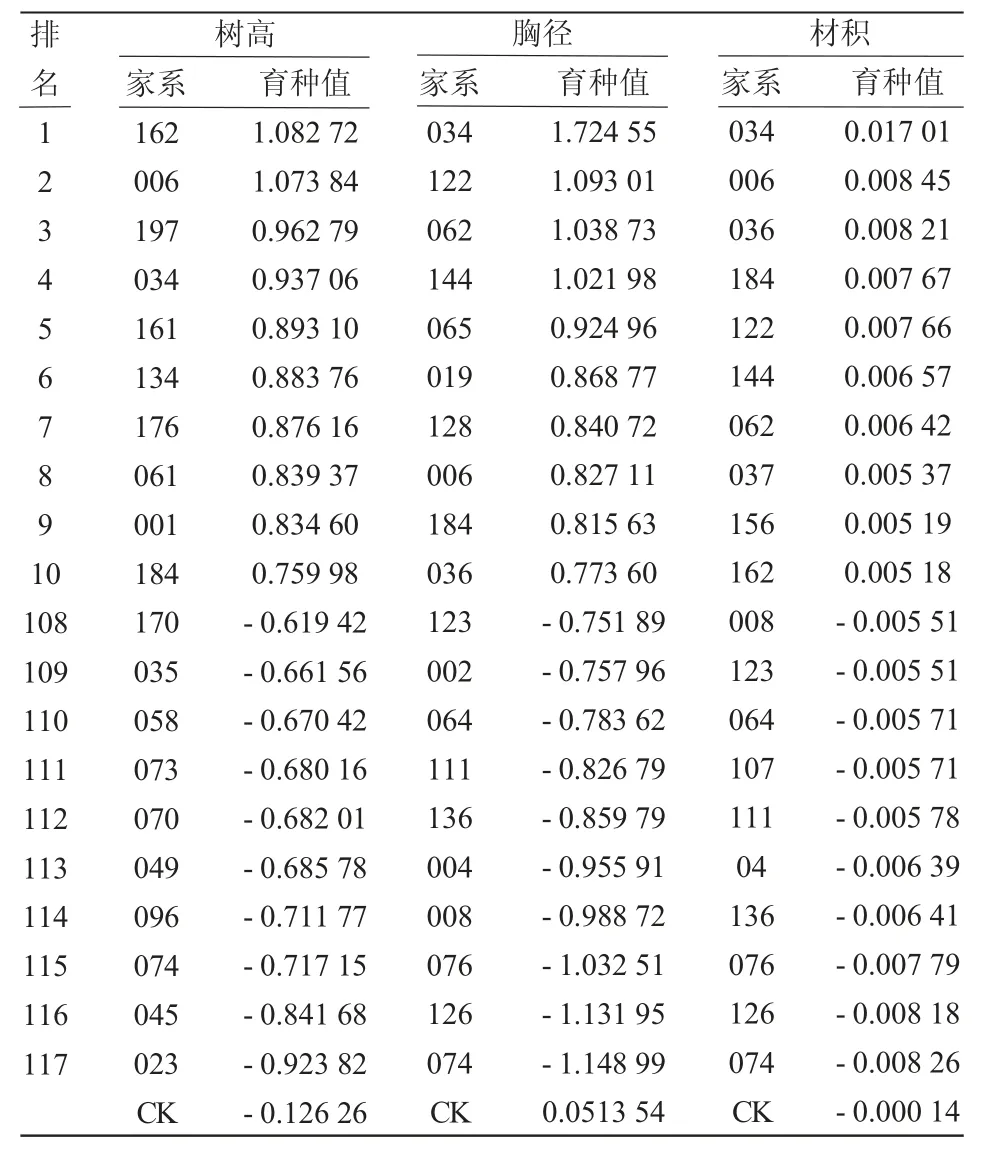
表5 红松自由授粉家系生长性状育种值估计
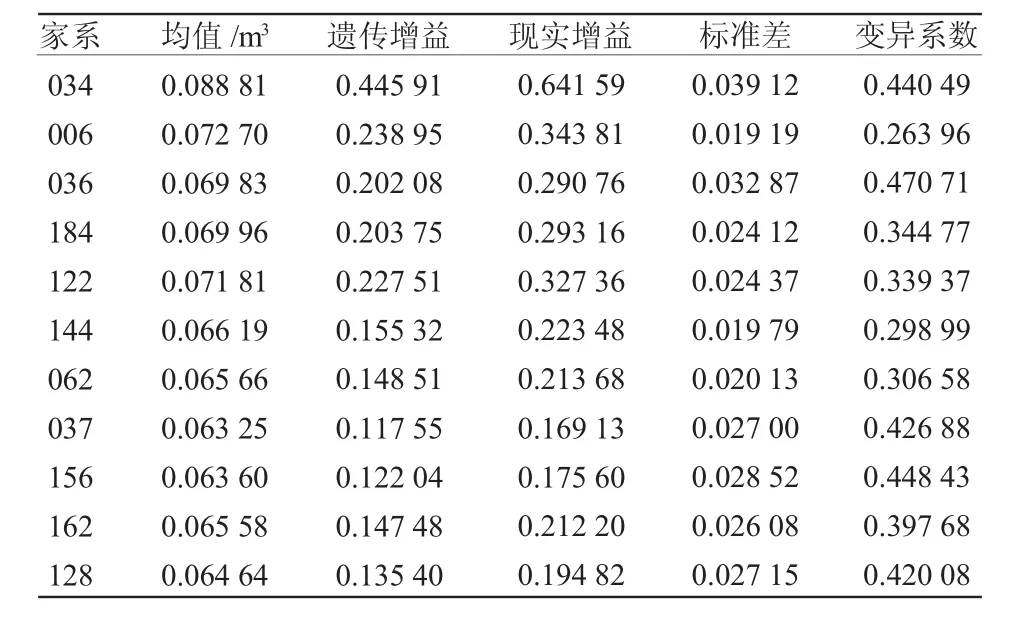
表6 优良家系材积性状遗传增益
以材积作为优良家系选择的主要指标,基于家系材积均值及育种值表现,选取家系材积均值大于CK 材积均值的家系034、006、122 等57 个家系作为初选群体,选取其中育种值排名在CK 之前的家系作为复选群体,以10%入选率对材积进行选择,入选的11 个优良家系分别为034、006、036、184、122、144、062、037、156、162、128,所选优良家系群体材积性状均值为0.069 28 m3,高出群体均值21.91%,高于CK 22.26%。
3 讨 论
遗传和变异是林木育种研究的主要内容,对变异来源、特点和规律的认识是进行树种改良的重要基础[14]。变异系数包括表型变异系数和遗传变异系数,是衡量群体变异程度大小,反应性状遗传变异能力的重要指标,变异程度高,有利于优良家系的选择[15-16]。本研究中27 年生红松自由授粉子代家系的树高、胸径材积性状均存在不同程度的变异,这与李艳霞等[17]在研究红松优良家系选择时得出的结论相符,变异程度大的家系可进行单株选择进行利用,变异程度小的家系可进行有性繁殖利用。其中材积性状的变异系数最大,说明以材积为选择指标对家系进行选择更具有潜力。遗传力可用来判断性状变异传递给后代的可能程度,是预估遗传增益的一个参考指标[18-19]。本研究中各生长性状的遗传力均处于较高水平,说明所选家系的各性状能较为稳定遗传,各家系生长性状家系遗传力均大于单株遗传力,这与油松[20](Pinus tabulaeformis)、杉木[21](Cunninghamia lanceolata)和华山松[22](Pinus armandi)的研究结果一致。最佳线性无偏预测(BLUP)方法是对遗传材料在不平衡环境下利用混合线性模型方程对育种值进行无偏预测[23-25]。育种值剔除了环境效应,反映真实遗传效应,有较高精确性[26-27]。本研究中不同家系性状均值表现与育种值表现存在一定程度不同,家系均值排名与育种值排名间存在5 名以内波动,这与欧阳芳群等[28]在青海云杉自由授粉家系遗传变异与基于BLUP 的改良代亲本选择研究中表现一致,反映出家系表型与基因型中加性效应部分存在一定程度差异。本研究以黑龙江省苇河林业局红松代测定林117 个红松自由授粉子代家系为材料,对家系树高、胸径和材积性状的遗传变异进行评价,所选的11 个优良家系优势明显。通过测定结果可对亲本无性系进行重新选择,同时可在优良家系内选出优良单株,为二代种子园的营建提供材料基础。
