辣椒根系影响下的农地土壤水分空间运动分异特征
2023-10-08陈晓冰秦梓城刘思佳刘俊杰
杨 婷,陈晓冰※,许 昊,秦梓城,刘思佳,刘俊杰
(1.桂林理工大学广西岩溶地区水污染与用水安全保障协同创新中心,桂林 541004;2.桂林理工大学广西环境污染控制理论与技术重点实验室,桂林 541004;3.广西环境污染控制理论与技术重点实验室科教结合科技创新基地,桂林 541004;4.桂林理工大学环境科学与工程学院,桂林 541004)
0 引言
土壤水作为农地水分循环的重要组成部分,也是影响田间作物生长发育的关键因子,对农业生态系统的物质能量流动具有重要意义[1]。因此,如何充分且高效地利用土壤水资源,实现农作物的提质增收是当前农业生产领域关注的研究热点。
目前,对于提高田间土壤水分的相关研究主要集中在探索耕作方式[2]、土壤环境[3]和作物类型[4]等方面上。土壤水分变化是影响作物需用水特性的重要因素之一[5],同时作物根系也对土壤水分特征产生影响[6]。根系生长过程中的穿插、绕结、挤压与腐烂死亡产生的通道引起土壤孔隙结构变化,从而改变土壤水分的分布状况[7]。同时,植物利用其根系分泌物和土壤微生物群落相互作用,从而胶结土壤颗粒以改善土壤微团聚体等结构状况[8],影响水分在土壤空间内的分布变化。当前关于根系影响下的水分状况,以分析其引起的土壤环境改变(诸如土壤理化性质、土壤结构等)进而影响土壤水分状况变化的研究居多。此外,相关研究发现根系生长会通过调节自身状态对土壤水分的吸收利用策略[9],从而影响地上作物生长和产量。根系的提水作用能够在植物生长期与土壤水分保持相对稳定的内在关系[10],通过实现水分再分配,使根土水环境维持平衡状态的同时也提高了根区从深层土壤得到水分补给的能力[11]。土壤水分运动作为反映田间水分状况的重要内容,会随着根土环境的改变发生变化[12],但其如何受到根系的影响作用,目前更多地是从根系影响下形成的土壤环境角度出发开展相关分析,鲜有从作物根系自身(根系形态、结构等)特性来进行水分运动作用关系阐述。而根系形态结构的可塑性[13]是植物在生长发育过程中主动适应土壤水分异质性环境、实现土壤水资源高效利用的体现,这对开展根系自身特性与土壤水分的协同调控机理的研究具有重要作用。因此,进一步阐明根系自身特性对田间土壤水分状况,特别是与其中的水分运动过程之间的作用关系,对于全面认识作物根系对田间土壤水分状况的影响,开展合理的田间作物灌溉具有重要的现实指导作用。
随着乡村振兴的大力发展,广西正逐步成为中国重要的南菜北运基地。其中辣椒作为广西种植面积较大的蔬菜品种之一,已经成为当地农业经济收入的重要来源[14]。近年来受气候变化的影响,广西灾害性天气加剧,高温干旱、暴雨洪涝等气象灾害频发[15],导致田间水热条件相对较差,严重影响辣椒生产。而辣椒作为主根粗、须根密集的一种作物,对土壤环境中水分状况极其敏感[16],不良的土壤水热环境一定程度上影响了广西辣椒种植。因此,本研究从解决当前广西辣椒种植产业的实际问题出发,以田间辣椒为研究对象,基于形态学图像解析技术、灰色关联等分析方法,定量细化田间根区和非根区的土壤水分空间分布、运动变化特征,揭示作物根系对土壤水分运动的影响,为开展田间辣椒精准灌溉,提高辣椒水分利用效率,实现辣椒提质增收提供理论支撑。
1 材料与方法
1.1 辣椒种植概况
研究区位于广西桂林市临桂区(25°17′N,110°01′E),属亚热带季风区,年平均温度为20 ℃,年平均降雨量为1 869 mm,近65%的降雨集中出现在4—7 月,大雨占比约26%。土壤以红壤土、石灰土和水稻土为主要土壤类型,土壤质地主要为粉壤土。选择研究区内长、宽分别为20 m 和10 m 的农用平地作为试验样地,种植前深翻土壤。研究地种植作物为辣椒(桂椒12 号),于2022 年4 月上旬定植,8—10 月收获,采用起垄深埋的种植方式,设置多个垄行,垄间距为15 cm,垄宽为120 cm,垄高为15 cm,采用垄上双株双行定植,株距50 cm,行距70 cm。施肥方式如下:底肥生物有机肥(有机质≥70%)300 kg/hm2,追肥复合肥380 kg/hm2(N 质量分数15%、P2O5质量分数15%、K2O 质量分数15%)。野外试验时间选择在辣椒成熟期,时间为2022年9 月20 日—2022 年9 月23 日,试验期间野外未出现降雨现象。研究样地土壤基本性质如表1 所示。
1.2 染色示踪试验
根据样地种植情况,随机选取3 个种植垄作为染色示踪试验区,清理表面杂草,将长60 cm、宽60 cm、高30 cm 和厚0.3 cm 的金属样方缓慢砸入试验区内(样方内植株具体布设位置见图1),砸入深度为15 cm。
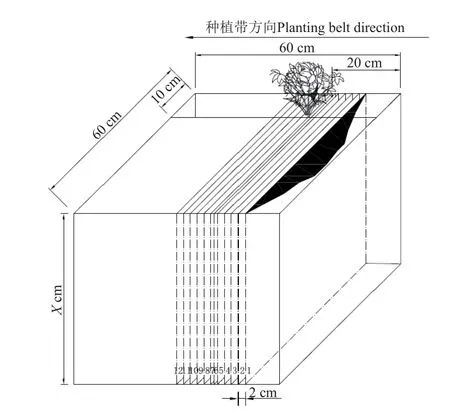
图1 土壤染色剖面示意图Fig.1 Soil dyeing profile diagram
为了防止降雨等外界因素的干扰,试验前使用塑料薄膜对样方进行24 h 覆盖。24 h 后移除金属样方上的薄膜,使用带有稳定恒流泵的供水装置将10 L(含5%的损耗)浓度为4 g/L 的亮蓝溶液(以当地累计24 h 的降雨量25 mm 为配置标准)以150 mL/min 均匀喷洒到样方内,喷洒结束后再用塑料薄膜覆盖样方24 h。24 h 后移除塑料薄膜,选择样方内植株前后10 cm 区域,以2 cm 水平宽度为一层,对试验区进行垂直剖面挖掘至最大染色深度处,当土壤剖面开始出现作物须根时,为减少挖掘过程对根系及其附近土壤染色剖面的影响,以1 cm 水平宽度缓慢向前挖掘,并配合使用毛刷等工具在根系生长区域缓慢刷去根系周围土壤并平整土壤剖面,使用1 510 万像素的单反相机(Canon EOS50D)对剖面图像进行拍摄。土壤垂直剖面挖掘空间示意如图1 所示。
1.3 根系采集与生物量测定
利用铁锹、镊子、毛刷等工具,采用全挖法对观测区辣椒根系进行采集,将根系整体、缓慢地从土壤中取出,尽可能保证根系结构的完整性,并使用皮尺测量记录辣椒最大扎根深度,使用自封袋将采集的根系带回实验室,将根系清洗干净后,采用植物图像分析仪(MICROTEK ScanMaker i800 plus)对根系进行扫描。将得到的根系扫描图导入万深LA-S 系列植物图像分析系统分析根系内部连接数量、外部连接数量、根系长度、根系广度、平均根系直径、总根表面积等根系参数。然后将样本置于80 ℃烘箱中烘干至恒质量,获得辣椒根系干质量,分别计算出根系形态(根系削弱系数、比根长)指标和根系结构(拓扑指数、分形丰度、分枝强度)指标。
1.4 数据处理与分析
1.4.1 染色图像数据处理
利用WGEO 软件对垂直染色图像进行校正,再通过Photoshop 2020 进行裁剪、颜色替换,灰度调节,利用MATLAB R2021b 对处理后的图片进行根系和土壤染色的分割,获得500×500 像素的土壤垂直剖面染色图像。最后利用Image Pro Plus 6.0 图像处理软件进行像素点的分割与数量的统计分析,分别获得0~255 灰度级数值矩阵和仅含有0 和255 的数值矩阵,用于后续土壤水分运动相关参数的计算。
1.4.2 土壤相关指标计算
1)土壤染色形态狭长度
土壤染色形态狭长度[17]是指土壤剖面水平和垂直方向上染色形态外切边界平行线距离的最小值与最大值之比,反映土壤染色形态的细长程度,其值越小,土壤水分竖向分布越狭长。
式中AR为土壤染色形态狭长度;Emin为土壤染色形态最小外切边界平行线距离,cm;Emax为土壤染色形态最大外切边界平行线距离,cm。
2)土壤剖面染色面积比
土壤剖面染色面积比[18]是土壤剖面染色面积占该剖面总面积的百分比,反映土壤中水流运动范围的大小。
式中DC为土壤剖面染色面积比,%;D为土壤剖面染色面积,cm2;ND为土壤剖面未染色面积,cm2。
3)土壤水分湍动强度
土壤水分湍动强度是各土层染色面积沿程累积面积变化幅度。用基于距离变化的沟道面积累计变化率[19]来表征土壤空间内染色面积累计变化率(即速变程度),数值越大说明土壤染色面积波动越大,水分运动速变程度越强,数值越小说明水分运动相对稳定。
式中TK为土壤水分湍动强度;Ki和Ki+1为各土层染色面积变化率,%;n为土层数量;K为相邻土层间汇入距离的变化的土壤染色面积变化率,%;Aa、Ab为2 个相邻土层的染色面积,cm2;Sab为相邻土层中心点之间的距离,cm。
4)最大入渗深度
最大入渗深度是指土壤水分最大入渗量,本研究中指采用部分染色达到的最深染色深度,cm。
1.4.3 根系形态与结构数据处理
1)根系削弱系数
根系削弱系数[20]是根系在不同土壤深度中根系的分布状况。β越小,表明根系在浅层土壤中分布的比例越大。
式中Y为从地表到一定土层深度的根系生物量累计百分比;d为土层深度,cm;β为根系削弱系数。
2)比根长
比根长[21]为根系长度与根干质量的比值。比根长值越大,根越细小。
式中SRL为比根长,m/g;l为根系长度,m;m为根系生物量,g。
3)拓扑指数
对传感器提供的信号评估和处理是生产高质量注塑件的先决条件,当然高精度注塑模具也是必不可少的,例如想要使生产部件公差小于30 µm,就必须使用误差低于10 µm的注塑模具。此外,还需要用于模具补偿的测量传感器,例如Carl Zeiss工业测量仪器公司就为此发开了一种用于表面反馈的创新软件。
拓扑指数[22]反映了不同植物根系的分支模式。植物根系分支介于鱼尾形分支和叉状分支之间,其值越趋于1,根系越接近鱼尾形分支;越趋于0.5,根系越接近叉状分支。
式中TI为拓扑指数;A为根系内部连接总数;M为根系外部连接总数。
4)分形丰度
分形丰度[23]是单位土体中植物根系的体积范围。其值越大,根系主根越发达,运输和吸收土壤水分和养分的能力越强。
式中Nr为根系所截的正方形数目;r为正方形边长;FD为根系分形维数;lgK为分形丰度。
5)分枝强度
分枝强度[24]表示根系分叉数的密集程度,也是衡量根系对土壤资源利用能力的指标。分枝强度越大,侧根数量增多,减少植株间可利用资源的传输距离,提高土壤水分资源利用率。
式中BI为分枝强度;e为根系分叉数;l为根系长度,mm。
1.4.4 数据统计分析
使用WEGO、Photoshop 2020、MATLAB R2021b和Image Pro Plus 6.0 等软件对染色图像进行处理和数据提取;使用Excel 2019 软件对提取的数据进行分析整理;使用SPSS 21.0 软件对数据进行方差分析、差异性分析和相关性分析(α=0.05);土壤水分湍动强度与根系形态与构型的灰色关联使用SPSSAU 进行计算;使用AutoCAD 2020 和Origin Pro 2020 软件对数据分析结果进行图表绘制。
2 结果与分析
2.1 辣椒根系形态及结构特征
2022 年9 月对辣椒根系最大扎根深度进行实地调查发现,根系在土壤中平均最大扎根深度可达25 cm。将野外采集的根系样品带回室内,通过植物图像分析系统计算分析了不同土壤深度下根系根径和比根长的变化,结果见表2。随着土壤深度的增加,根径逐渐减小,比根长也呈增加趋势且在>20~30 cm 达到最大值,说明土壤深度越深,辣椒根系越细小。
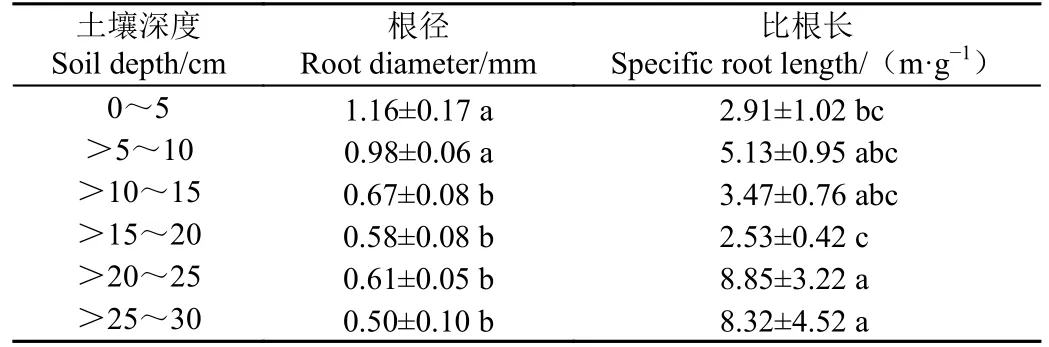
表2 不同土壤深度根径和比根长变化Table 2 Changes of root diameter and specific root length in different soil depths
对辣椒根系整体进行根系形态(根径、根系广度和比根长)和根系结构(拓扑指数、分枝强度和分形丰度)分析可知,根系平均根径为0.47 mm,其生长平均广度为18.46 cm,比根长为14.72 m/g,细根较多。同时辣椒根系平均拓扑指数为0.87,趋于1,根系接近鱼尾形结构。根系平均分形丰度为14.45,分枝强度为0.53,辣椒根系分支较多,侧根密集,在土壤中拓展体积较大。对辣椒根系形态和根系结构进行相关性分析,结果见表3。扎根深度与分枝强度相关性最大(相关系数为0.899),根系广度与分枝强度相关系数为0.883,表现为根系广度越广,分枝强度越大。根系广度与分形丰度存在极显著相关关系(P<0.01),相关系数为0.986。整体而言,反映根系形态的根系扎根深度和根系广度与根系结构的相关程度更高。
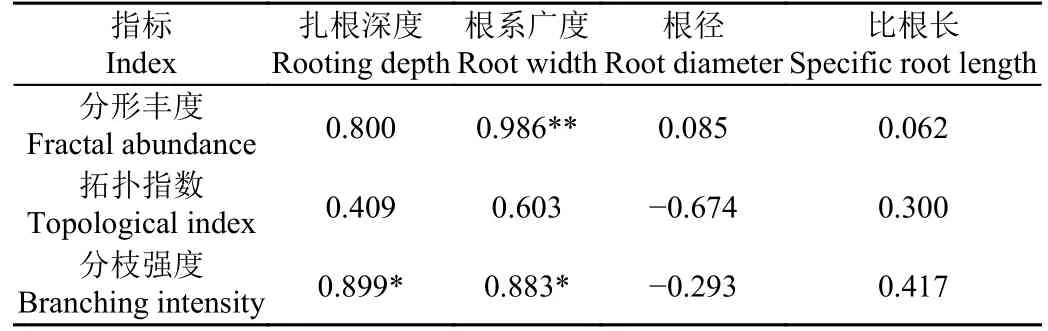
表3 根系形态与结构间相关性Table 3 Correlation between root morphology and structure
2.2 辣椒根系影响下的田间土壤水分空间运动
土壤剖面染色形态可在一定程度上表现土壤水分入渗的运动范围,进而反映根系影响下的土壤水分空间分布状况。对染色示踪试验获得的36 张土壤垂直剖面染色图像数据进行分析,选取最接近染色面积比均值(28.63%)的相对应观测区的染色图像进行土壤水分空间分布状况展示分析,土壤染色相关指标计算采用全部土壤剖面染色数据,结果如图2 所示。
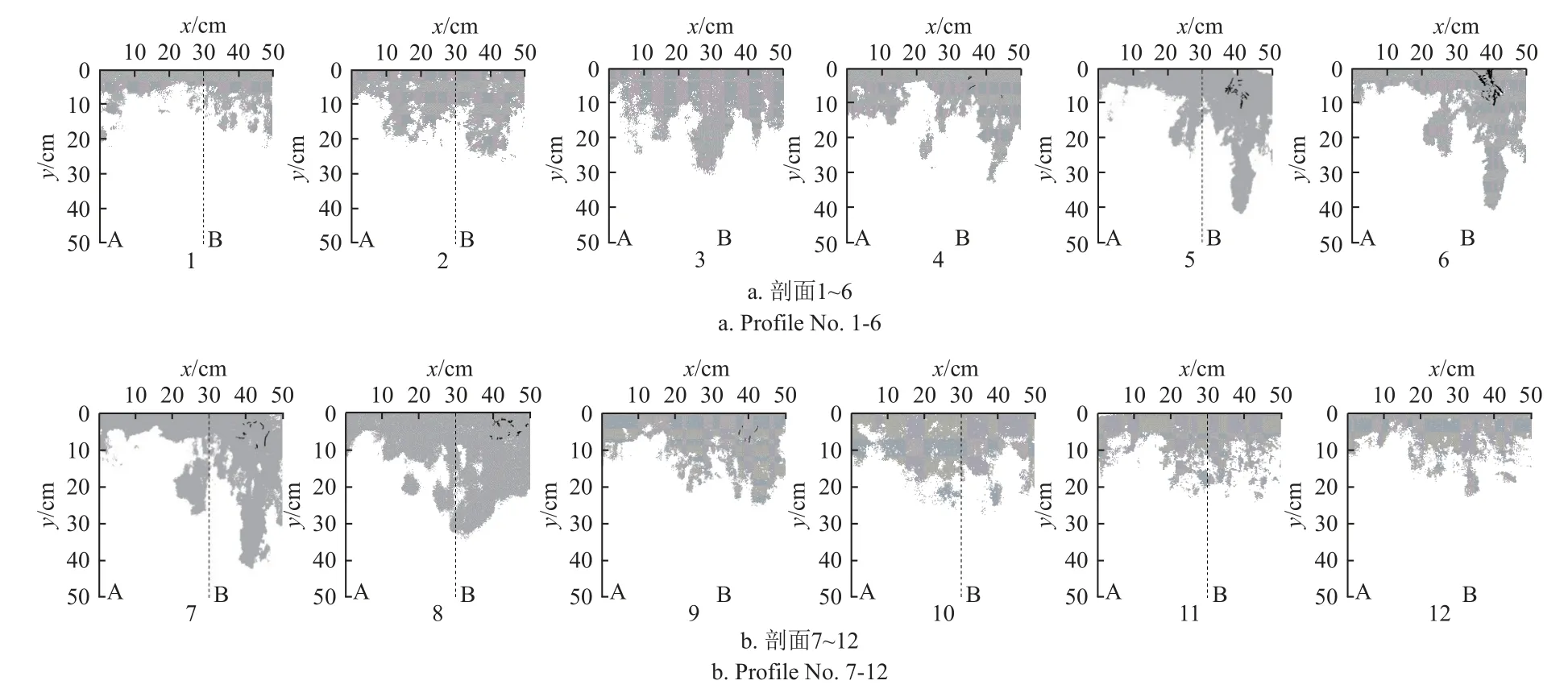
图2 辣椒地土壤垂直剖面染色图像Fig.2 Dyeing image of chili pepper along soil vertical profile
由图2 可知,土壤染色形态在0~5 cm 土层范围内未分化,在5 cm 以下土层沿深度开始出现较明显的团块状分化现象,整体沿土壤深度呈现增加的变化趋势,且染色形态出现侧向偏移,表明土壤剖面中存在侧向水分运移通道,引起水分在下渗过程中发生侧向运动。当逐渐靠近辣椒主根剖面(剖面6)附近时,染色水流向主根附近发生聚集变化,土壤剖面染色形态呈现“指状”分化现象。而特定的根区对根土空间的影响至关重要,尽管根系、土壤、环境等因素影响根土环境到达平衡的时间,但潜在根系影响范围几乎保持不变[25]。因此结合染色实际形态与辣椒根系(平均)生长宽度,将染色剖面沿水平宽度方向分为非根区(0~30 cm)和根区(>30~50 cm)。非根区在0~10 cm 染色多以整体入渗为主,>10~20 cm 由整体入渗向集中入渗过渡,>20~30 cm 土层出现散点状的染色形态,且在>30~50 cm 土层土壤未出现染色。相较于非根区,根区土壤染色图像在0~30 cm 土层呈现集中聚集分布特征,在辣椒主根附近(剖面6)的>30~50 cm 土层出现了“指状”染色形态的水分分布状况,并贯穿土壤剖面。非根区土壤染色形态平均狭长度为0.89,是根区(0.61)的1.5 倍(图3),二者之间差异显著(P<0.05)。说明根区土壤染色形态较狭长,水分空间分布相对集中。结合土壤剖面染色图像分析得出,非根区土壤水分多以整体和侧向分布为主,而根区在浅层土壤中水分分布较集中,但随着土壤深度增加,出现了上下贯通的垂向通道,深层土壤水分分布显著。
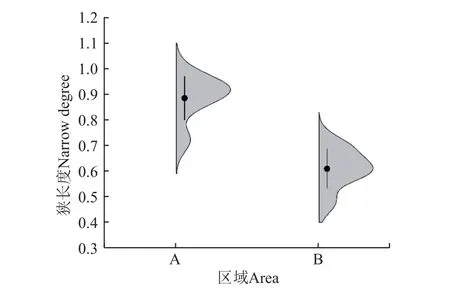
图3 辣椒地土壤染色形态狭长度Fig.3 Pepper soil dyeing form narrow degree
在野外染色示踪试验中,基于根系主导的水流运动范围所形成的染色形态变化可以反映出水流入渗运动的空间分布特征,而土壤染色面积比可定量反映土壤水分的空间运动结果(图4)。在0~10 cm 土层中,非根区染色面积趋于100%,染色面积变化较小,而>10~30 cm 土层中随着土壤深度增加,染色面积比波动较大,在>30~40 cm 土层中染色面积比逐渐趋于0,说明该土层是非根区水分运动能到达的平均最大深度范围。而根区染色面积比呈现随土壤深度增加而减少的趋势,相较于非根区,染色面积比变化较为平缓(>10~20 cm 土壤深度),水流运动范围无较大变化,且在>30~50 cm 土层中发生拖尾现象,根区染色沿土壤深度呈现集中运动。根区在>20~50 cm 土层中,染色面积比快速下降,说明土壤侧向运移通道减少,水分以垂直向下的非均匀流形式快速运动,且根区最大水流到达深度范围(>40~50 cm)显著大于非根区(P<0.05)。根区土壤平均总染色面积比为36.84%,是非根区(23.62%)的1.6 倍(P<0.05),表明在整个土壤空间内,根区土壤染色覆盖变化范围较小,染色分布较广泛,水分空间运动范围相对稳定。
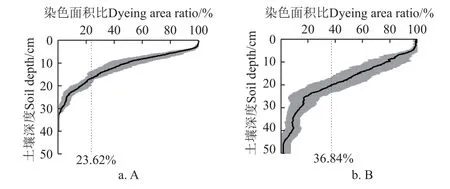
图4 辣椒地土壤染色面积比随土壤深度的变化Fig.4 Changes of soil dyeing area ratio with soil depth in pepper field
为进一步表征辣椒根系影响下不同土层水分空间运动变化程度,对非根区和根区的土壤水分湍动强度进行分析(图5)。非根区不同土层土壤水分湍动强度起伏大,整体呈“S”型曲线波动,而根区整体呈线性递增。在0~20 cm 土层,非根区水分平均湍动强度为14.48,是根区(7.93)的1.6 倍,土壤水分活跃程度更高。随着土层深度(>20~40 cm)增加,非根区和根区的水分湍动强度均逐渐增强,非根区湍动强度达到最大值34.10,远大于根区(22.33),说明非根区水分竖向速变程度较高,水分入渗相对减少。在>40~50 cm 土层,非根区湍动强度减弱,根区土壤水分逐渐到达平均最大入渗深度,湍动强度持续增强,在此深度下非根区达到31.04。结合土壤染色图像,非根区在>20~40 cm 土层的水分空间活跃程度高。辣椒地非根区土壤水分平均湍动强度为116.09,是根区(83.90)的1.4 倍(P<0.05),表明根系影响土壤水分的竖向运动,使土壤水分深层运动能力增强,但深层土壤侧向运移通道相对表层土壤减少,土壤水分速变程度整体较弱。
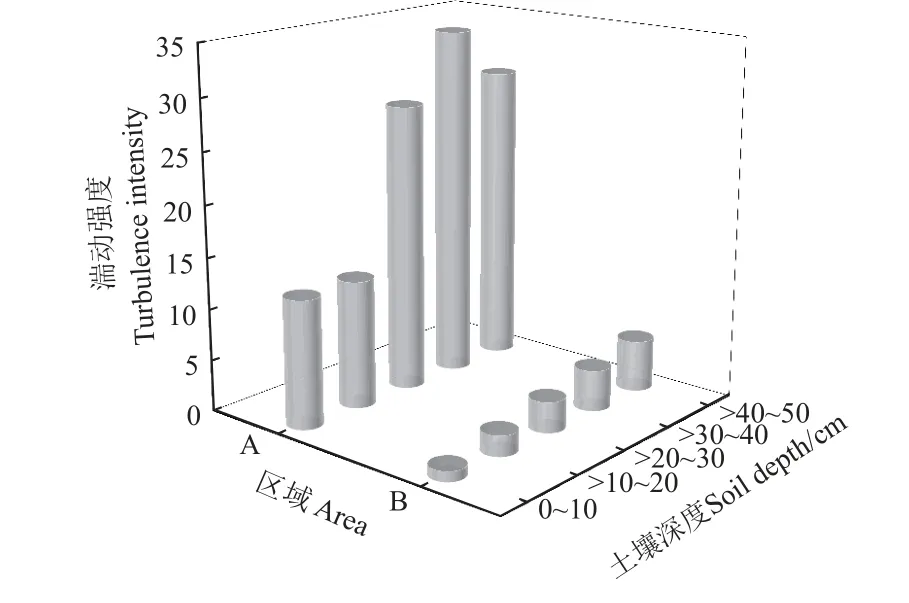
图5 辣椒地土壤水分湍动强度Fig.5 Turbulence intensity of soil moisture in pepper field
2.3 辣椒根系形态结构对土壤水分空间运动变化的影响
湍动强度量化反映了土壤水分空间运动的速变程度,为定量分析根系形态及结构对土壤水分空间运动分异特征的影响,以根区土壤水分湍动强度和根系形态及结构分别作为参考序列和比较序列,对根区土壤水分湍动强度进行灰色关联分析(表4)。反映根系形态状况的根系比根长对土壤水分湍动强度影响最大,关联度为0.984,其次为反映根系结构的根系分枝强度,关联度为0.720,根径(关联度为0.486)影响最小。而根系广度、拓扑指数和分形丰度对土壤水分湍动强度影响相差不大,关联度分别为0.684、0.687 和0.669,均高于根系扎根深度(0.555)。整体而言,根系形态对土壤水分的空间运动异质性影响较大,而根系结构对土壤水分的变化影响程度相对较小,且表征根系结构的特征参数影响程度相似。

表4 辣椒根系形态及结构与土壤水分湍动强度的灰色关联Table 4 Grey correlation between characteristic parameters of root development of chili and turbulence intensity of soil moisture
为进一步量化根系对土壤水分空间运动的影响,对辣椒地非根区和根区土壤染色图像进行灰度值提取,灰度值的大小可以反映土壤染色的深浅变化,进而表征土壤水分含量的大小(即染色越深,灰度值越小,土壤水分含量越高)。而灰度值频率描述了单位空间内某一灰度值出现的次数,不同灰度值出现的频率表征根区和非根区水分含量的高低。由图6 可知,在土壤0~50 cm水平宽度内,土壤染色平均总灰度值整体呈现“下降-上升-下降-上升”的变化趋势,在水平宽度41.6 cm 处平均总灰度值总最小(991 513),说明该位置处土壤颗粒染色最深,土壤水分含量最高。非根区和根区土壤染色平均灰度值频率在灰度级范围内均呈现先增加后降低,随后在一定范围内波动的变化趋势,但在51~153 灰度级范围内,根区平均灰度值频率显著高于非根区。非根区平均总灰度值整体呈下降趋势,其平均总灰度值为1 251 648,平均灰度值频率在51~102 灰度级范围内下降到最小值0.09%后出现小范围的增加趋势(102~153 灰度级范围内),最后保持“锯齿形”波动状态。而根区整体呈“V”字型变化趋势,越靠近辣椒主根定植区域,平均总灰度值越小,表明主根附近土壤含水量越高。根区平均总灰度值为1 099 961,平均灰度值频率下降到0.23%附近(51~102 灰度级范围)后,在102~153 灰度级范围内保持在0.23%附近平稳波动状态。根区平均总灰度值频率(51~153 灰度级范围)为31.34%,是非根区(19.36%)的1.6 倍。因此,在相同外部供水环境下,根区的土壤水分运动分布程度更高,水分运动更活跃。
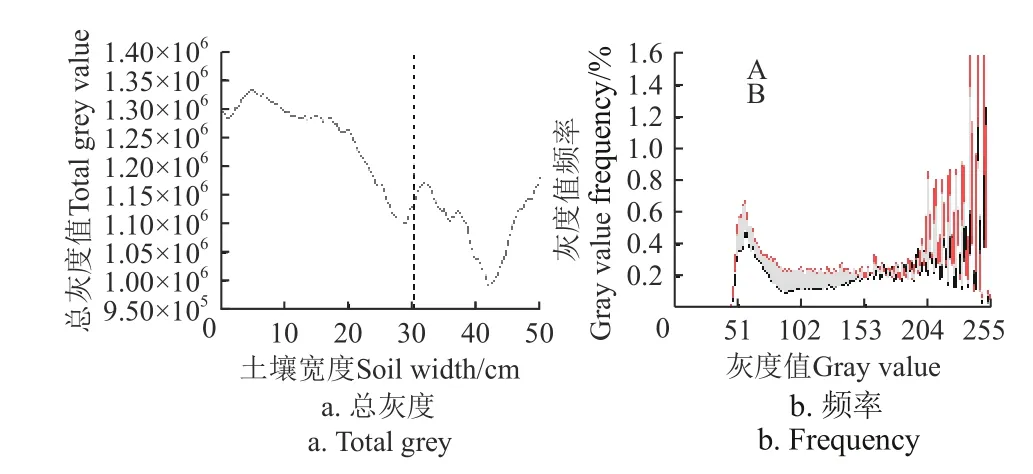
图6 辣椒地土壤染色灰度直方图Fig.6 Gray histogram of soil dyeing in pepper field
综上,根区土壤含水量显著高于非根区土壤含水量。根系削弱系数可表征根系在土壤垂直空间内的分布特征,其与土壤水分间的相互作用量化反映了水分运动状况。从表5 可知,根系削弱系数随着土层深度逐渐增大,在>20~30 cm 最接近1,在0~20 cm 土壤深度相对较小,说明辣椒根系在浅层土壤(0~20 cm)中分布比例较大。同时,在0~20 cm 土层中,土壤含水率呈上升趋势,含水率差异显著(P<0.05),而从15 cm到30 cm 土层土壤含水率降低,说明在辣椒根系浅层分布区,土壤水分状态相对活跃,且土壤水分呈现集中分布状态。
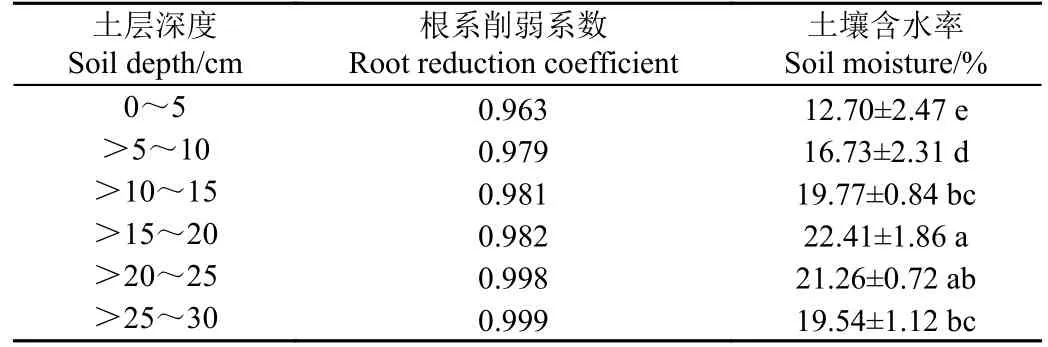
表5 不同土壤深度根系削弱系数及土壤含水率Table 5 Root reduction coefficient and soil moisture content at different soil depths
3 讨论
3.1 辣椒根系分布对土壤染色形态特征的影响
根系通过自身形态及结构来适应土壤空间内的水分环境[26],同时改变了土壤水分分布状况,使得根系生长区域范围内的土壤剖面内出现不同的染色形态分化现象。本研究中的辣椒根系虽然接近鱼尾形分支结构,但随着扎根深度的增加,根系广度变广,根系分形丰度增高,而较高的分形丰度表明辣椒根系细根占比多,分支多且分枝强度更大,侧根更密集,根系在土壤中拓展的体积更大[23],能够在更大的水平空间内获取土壤水分,从而使土壤染色形态在辣椒根系生长区域出现集中聚集分布形态。通过对非根区和根区辣椒地土壤染色形态特征分析发现,根系生长区域染色呈聚集状分布,水分主要分布在0~30 cm 土层内,非根系生长区域表层土壤以整体分布为主,表明根系的生长会对土壤原有结构产生扰动,形成以作物根系为主的根土结构环境,并与现有的土壤水分孔隙路径相连通,增强水流在土壤竖向空间内的运动过程[27],形成聚集的染色形态分布现象。本研究通过对同一土壤非根区和根区的染色形态狭长度分析发现根区水分分布范围更广,水流能到达的土壤深度更深,这与作物根系扎根深度和根系广度有关,研究表明[28],当浅层土壤含水量无法保证作物生理需求,根系会向土壤深层生长,以吸收利用深层土壤水分。而非根区土壤表层富集水分较多,水分深层分布不显著,这可能是因为根区根系的生长对土壤颗粒存在压实作用,使非根区土壤水分呈现表层整体分布特征。同时,在整个土壤空间内,根系的生长极大增强了水分的入渗能力[29]。
3.2 辣椒根系对土壤水分空间运动的影响
本研究通过对0~50 cm 土壤深度染色面积比图像进行分析发现,0~30 cm 土层中非根区染色面积比波动大,根区水分运动范围变化平缓较小,而在深层土壤中,非根区水分入渗较少,说明在同一供水条件下,根土及附近形成的土壤环境可以保持较好的水分入渗条件,使土壤具有高渗透性和良好的垂直连通性[30],这与WU 等[31]对不同植物根系土壤优先流入渗机制的研究结果一致。同时,本研究中根区土壤平均染色面积比是非根区(23.62%)的1.6 倍,且根区土壤水分湍动强度弱,其空间运动范围速变程度平缓,水分运动连通性优于非根区,这可能是因为根区表层土壤水分优先通过根系收缩作用形成的根土间隙运至土壤深层,促进土壤水分快速运动。已有研究表明[32],根土间隙是土壤大孔隙的重要组成部分,在土壤水分运动过程中发挥重要作用。本研究中作物根系分布在0~30 cm 土层中,在此深度下根土间隙促进了水分的垂直入渗,但土壤的紧实性会影响水分的垂直入渗能力[33]。对于整个土壤空间而言,根系提高了表层土壤蓄水能力,同时也在一定程度上促进了水分的竖向入渗运动,这有助于土壤空间内的水分交换,提高作物的水分利用能力[34]。
3.3 辣椒根系与土壤水分空间运动分异的作用关系
通过对根系形态及结构与土壤水分湍动强度进行灰色关联分析发现,表征根系形态的比根长对土壤水分湍动强度影响更大,其次为根系结构中的分枝强度,表明根系的存在增加了水流运动阻力[35]。比根长决定了作物根系利用土壤水分和养分的能力,研究表明[21,36],作物能够通过增加比根长从而提高对水分和养分的吸收。本研究中辣椒作为主根粗、须根密集的作物,即使最大扎根深度无法穿透土壤深层,但根系对土壤产生的扰动使根区与深层土壤建立了竖向连通性,土壤深层水分运动以“指状”形式向下入渗,促进了深层土壤优先流的发育[37],致使水分运动在含有根系的土壤空间内呈现明显的分异特征。
辣椒根系的削弱系数在0~20 cm 土层深度较小,趋于土壤浅层分布,促进了土壤表层蓄水能力。研究发现,相较于非根区,根区土壤含水量更高,且越靠近辣椒主根定植区域,土壤含水量越高,这是根系适应土壤环境从而满足自身吸收利用的体现。有研究发现[38],由于根系在土体内的分布延展及渗透压的影响,将会把土壤水聚集在根系四周的利用区域内,有利于作物根系对土壤资源的吸收利用,且土壤含水率随着土壤深度增加呈现先增大后减小的变化特征。李惟婕等[39]发现土壤水分与果树细根长密度呈负相关关系,这与本研究结果具有一定相似性。
以往关于根系对土壤水分状况的研究较多基于在根系影响下的土壤环境(诸如结构、理化性质等)情况下分析所得,而本研究通过从作物自身特性出发,围绕根系的形态结构对土壤水分空间运动变化进行研究,同时对土壤空间进行非根区与根区的水流染色区域分割,细化揭示了了作物根系对农地土壤层中水分分布变化以及空间运动的影响。但在根土间隙区,根系如何在此微观空间内利用狭窄区域及自身生理特性影响土壤水分状况,且存在的根系分泌物和菌丝际及其他微生物与根系结合形成的根际共同体[40]是否对土壤水分空间分异产生影响,还需进一步探索研究。
4 结论
本研究通过野外染色示踪试验,结合形态学图像解析技术,分析了田间土壤根区和非根区的水分空间分布,探讨了辣椒根系影响下的农地土壤水分空间运动分异特征,得出如下结论:
1)根系形态与结构存在相关关系,进而影响土壤水分空间运动,反映根系形态的根系扎根深度(相关系数为0.899)和根系广度(相关系数为0.883)对分枝强度的影响最大,根系广度与分形丰度极显著相关(P<0.01)。
2)在相同供水条件下,根区土壤水分在0~30 cm土壤深度范围内呈现聚集状态,深层土壤水分以“指状”形式分布。非根区土壤水分以土壤表层0~10 cm 整体分布为主,深层无土壤水分分布,其土壤染色形态平均狭长度是根区的1.5 倍(P<0.05)。非根区土壤水分速变程度(平均土壤水分湍动强度为116.09)强于根区(83.90),差异显著(P<0.05),根区水分空间运动的整体性和连通性均较好,土壤水分竖向入渗程度高。
3)根系形态相比根系结构对土壤水分空间运动分异特征影响程度更大,其中比根长对土壤水分运动影响最大(灰色关联度值为0.984),而根系结构对土壤水分变化的影响程度相接近。在51~153 灰度级范围内,根区平均总灰度值频率为31.34%,是非根区的1.6 倍,土壤含水量更高,根区土壤水分运动更活跃。
