上海主城区绿地对鸟类多样性的影响及其边际效应*
2023-09-19王云才卢星昊王若静
王云才 卢星昊 王若静
同济大学建筑与城市规划学院 上海 200092
城市化是导致全球鸟类多样性锐减的重要原因[1-2]。一方面,建设用地扩张导致鸟类栖息地大量丧失或生境破碎化;另一方面,城市绿地面积锐减与外来物种引入加剧城市生物同质化[3-4]。鸟类是城市生态系统重要组成部分,因其易于观测且对栖息环境变化敏感,被视作城市生物多样性的指示物种[5-6]。由于鸟类难以在不透水面与建筑密集区域生存,绿地成为鸟类重要的栖息地与食物源地,研究表明昆虫群落丰富度与鸟类多样性显著相关[7-8]。绿地环境特征同样影响鸟类栖息地选择。绿地面积是影响鸟类多样性最重要变量,并呈显著正相关[8-9]。Wong等[7]研究表明绿地形状规则有助于提升鸟类多样性。植被覆盖面积、组成、树木高度与冠幅同样影响鸟类群落构成[10-12]。此外,也有研究探讨城市建成环境因子如噪音、距市中心距离、建筑高度与密度等对鸟类多样性的干扰[13-14]。
环境因子常具共线性特征,传统分析方法如多元线性回归、结构方程模型难以准确揭示其与鸟类多样性的相关性,而机器学习算法如随机森林(Random forest,RF)、贝叶斯加性回归树(Bayesian additive regression trees,BART)、增强回归树(Boosted regression tree,BRT)在拟合过程中可以有效消除因子间的相互作用,具有较大的应用潜力。BRT模型生成的边际效应曲线可以有效揭示变量潜在影响随其量级变化的特征[15-16],目前已被应用于物种分布与环境因子的相关性分析及物种分布预测模拟,如Meffert等[16]应用BRT模型探究濒危鸟类分布与城市荒野环境特征的相关性。
数据采集技术的进步促进鸟类多样性研究[17]。传统鸟类群落调查以样点或样线计数等结构化方式采集鸟类数据[18-20],数据获取困难限制了鸟类多样性研究,随着公众科学项目兴起,数据采集方式得到进步,鸟类多样性分析时空尺度得以提升。已有学者基于中国鸟类记录中心、eBird、iNaturalist与Burungnesia等鸟类观测网站数据开展鸟类群落分析、景观尺度下鸟类多样性格局成因探测与生境网络构建等相关研究[21-24]。
总体而言,绿地在维持城市鸟类多样性方面发挥重要作用,然而鸟类多样性对于环境因子的响应具有尺度依赖性,受多种因素限制,鸟类多样性对城市绿地景观变化,尤其小尺度景观变化响应不一致,目前该机制成因也尚未清晰[25]。因此,选取上海主城区为研究区域,基于长时序、多样本的鸟类观测记录数据,从绿地景观组分、质量、结构与邻域环境4个维度选取变量,应用BRT模型探测与模拟绿地环境因子对鸟类多样性影响及其因子变化对鸟类多样性产生的边际效应,以期为绿地鸟类生境营造与多样性保护提供科学指标阈值依据。
1 研究区概况
上海地处长江入海口,是“东亚-澳大利西亚”候鸟迁徙通道的重要中转站[26],属亚热带季风气候,地势平坦,平均海拔约4 m。《上海市鸟类名录》显示,截至2019年,上海市共有鸟类22目78科494种,占全国鸟类种数的33.51%。同时,鸟类占上海全市野生动物种数的80%以上[27]。随着上海生态之城建设推进,2021年底上海森林覆盖率达19.42%,公园绿地面积达22 463 hm2,城市绿地作为鸟类栖息地也愈发受到重视。《上海市生态空间建设和市容环境优化“十四五”规划》提出,到2025年,城市生物多样性指数(鸟类)达到0.6以上,2035年基本建成人地和谐共生的生态之城。在此背景下,本研究选取上海城市化水平较高的主城区(1 278.47 km2)作为研究范围,选取207个绿地样本探究环境因子对于鸟类多样性的影响。
2 研究方法
2.1 鸟类调查方法与多样性分析
本研究参考《上海2019鸟类名录》《中国鸟类分类与分布名录》筛选出上海潜在鸟类共计494种,通过eBrid与中国鸟类记录中心网站逐一筛选位于上海主城区的鸟类观测数据,并结合中国鸟类记录中心网站的鸟类观测报告进行数据核对与校正。同时随机抽取20个样本(占总数9.66%),通过样点法观测核对网络与实地数据的一致性,检验结果表明网络数据基本符合实际。最终筛选共计17 461条观测记录,包含311种鸟类(占总类型数62.96%)。其中,国家一级保护野生动物2种,二级保护动物40种,极危鸟类4种,濒危鸟类4种,易危鸟类4种,近危鸟类15种。
本研究选取香农多样性指数、丰度、多度、均匀度及相似度衡量绿地鸟类多样性水平[24,28]。绿地鸟类优势种按照Berger-Parker法确定,其计算公式为[29]:
式(1)中:I为优势度指数;Ni为物种i在该绿地样本中的个体数量;N为该绿地中全部鸟类总个体数。当优势度指数I≥0.10时,定义该物种为优势种。
2.2 绿地环境因子选取
选取7项绿地环境因子,包含绿地景观组分、结构、质量与邻域环境4个维度(表1)。

表1 绿地环境因子选取及计算方法
2.3 增强回归树模型
BRT模型是基于分类回归树模型的自学习算法,该模型消除了因子间交互作用,在自学习与多次随机构建简单回归树过程中考虑预测结果的误差不断迭代建立多重回归树以提升模型预测的精度[30]。模型输出的结果包括各变量的贡献率及其边际效应,当边际效应大于0时表明变量对于因变量具有正向影响,反之为负向影响[31]。本研究应用R中“gbm”包完成BRT模型分析,其中模型参数学习速率设置为0.005,决策树复杂度为5,抽取比例为0.5。
3 结果与分析
3.1 鸟类物种组成与多样性
3.1.1 鸟类物种组成与优势种生境特征
207个绿地样本中共记录到鸟类12 556只,隶属19目63科311种,其中雀型目33科138种,占鸟类种数的44.37%,居于鸟类种数首位;其次为鸻形目8科66种,占21.22%。以居留型而言,冬候鸟106种,占34.08%;旅鸟90种,占28.94%;留鸟65种,占20.90%;夏候鸟50种,占16.08%。为分析不同生活型鸟类对于栖息地生境的偏好及其指标的数值分布,本研究依据鸟类栖息生境与生态习性将鸟类分为游禽、涉禽、陆禽、攀禽、鸣禽和猛禽6种类型。
陆禽优势种如山斑鸠(Streptopelia orientalis)、珠颈斑鸠(Spilopelia chinensis),其主要分布于外环及环内的中小型绿地(共50个,占总数24.15%)。由图1可知,陆禽优势种所在生境的GR、HQ与GLPI水平较高,中位数分别为0.83、0.79和0.46,表明其偏好乔灌木丰富、生境质量较优的绿地环境。
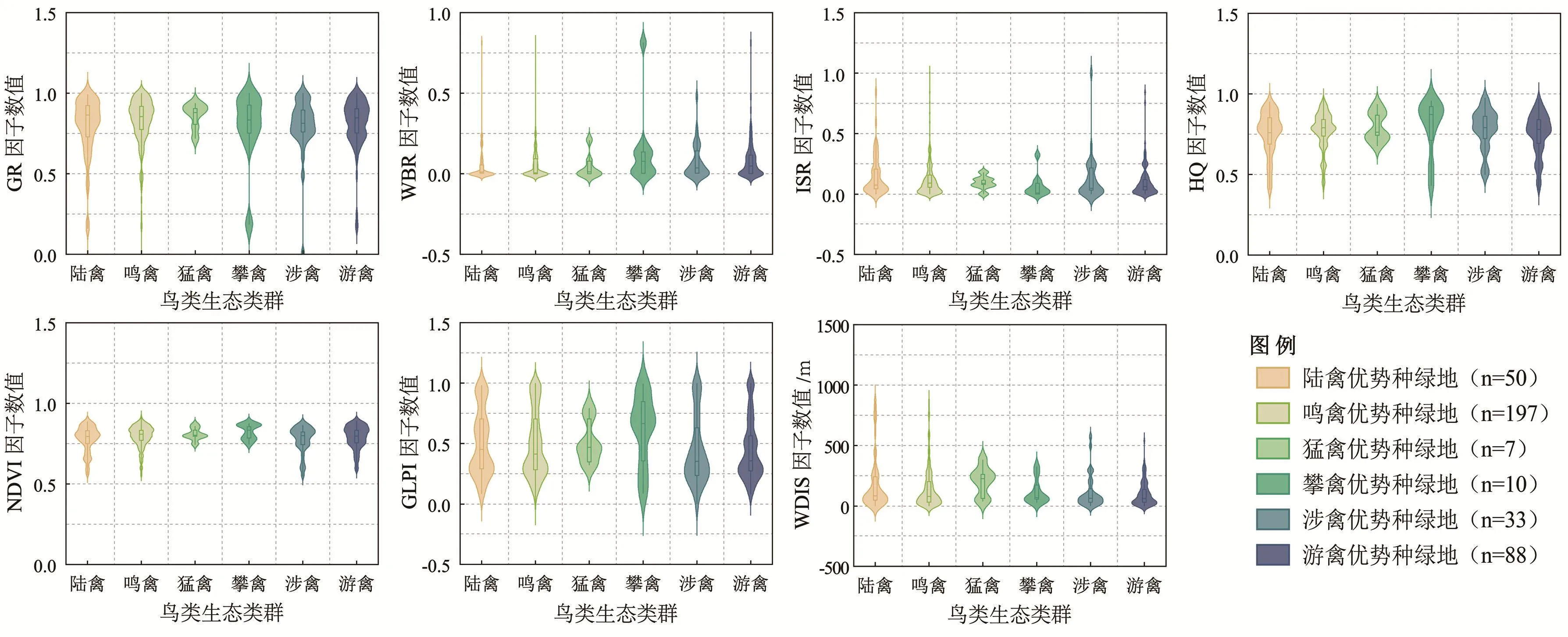
图1 优势种分布绿地的生境特征
鸣禽优势种主要分布于世纪公园、上海植物园与外环绿带等大型绿地(共197个,占总数95.17%),如灰喜鹊(Cyanopica cyanus)、麻雀(Passer montanus)等,通常对城市环境具有较强适应性。其优势种所在绿地GR、HQ、NDVI水平较高(图1),中位数分别为0.85、0.79、0.80。
猛禽优势种主要分布于滨江森林公园、共青森林公园等大型绿地(共7个,占总数3.38%),以凤头鹰 (Accipiter trivirgatus)、普通鵟(Buteojaponicus)等为主,通常对水环境依赖度较低,偏好高大成熟的大面积连续混交林。猛禽优势种所在绿地GR与WDIS水平较高(图1),中位数分别为0.88和226.84。
攀禽优势种主要出现在世纪公园与上海植物园等大型绿地(共10个,占总数4.83%),常见有戴胜(Upupa epops)、普通翠鸟(Alcedo atthis)等,通常偏好植被覆盖度较高且距离水系较远的完整绿地,其优势种分布绿地WBR、HQ、NDVI、GLPI水平较高(图1),各因子中位数分别为0.08、0.87、0.84、0.67。
涉禽优势种多出现于世纪公园、吴淞炮台湿地公园等(共33个,占总数15.94%),常见有黑水鸡(Gallinula chloropus)、骨顶鸡(Fulica atra)等,通常栖息于具有一定人鸟距离的湖塘、河口、芦苇及海滨沼泽等。其优势种所在绿地各指标均处于中等水平(图1),GLPI水平最低(中位数为0.36),其次为WDIS(中位数为65.12)。
游禽优势种常栖息于世纪公园、迪士尼星愿公园、东海沿岸等绿地的各类水域(共88个,占总数 42.51%),常见如斑嘴鸭 (Anas zonorhyncha)、小(Tachybaptus ruficollis)等,偏好生境条件好、水生植物丰富、深度适宜的水域环境,常在岸边乔灌木筑巢,其优势种分布的绿地中(图1),GR水平较高(中位数为0.84),而WDIS最低(中位数为63.50)。
3.1.2 绿地鸟类多样性与空间分布特征
上海市绿地鸟类多样性格局呈现显著的空间异质性(Moran I=0.05,z=2.81,P<0.01)。由图2A可知,浦东滨江森林公园鸟类香农多样性居于首位(4.78),其次为吴淞炮台湿地公园(4.41),再次是上海植物园(4.36)与世纪公园(4.28)。沿海滨江绿地鸟类香农多样性指数均值最高(1.99),其次为远离河流、海域的大型绿地(均值为1.98)。此外,上海部分外环结构性绿地鸟类香农多样性水平也较高(均值为1.78)。
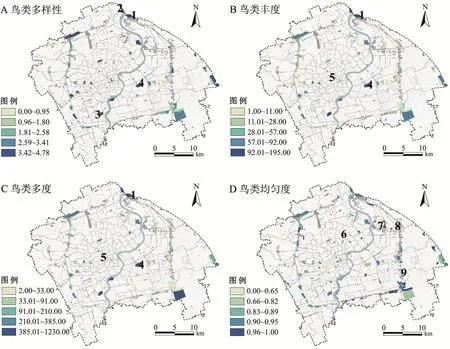
图2 鸟类多样性的空间分布
鸟类丰度与多度的空间分布格局与香农多样性呈相似特征(图2B~C),世纪公园鸟类种类与数量最多(195种,1230只),其次分别是静安雕塑公园(130种)与浦东滨江森林公园(629只)。沿海滨江绿地鸟类丰度与多度均值最高(75.98,21.43),集中建设区绿地 (56.68,17.53)、外环结构性绿地(40.07,13.58)次之。
由图2D可知,均匀度较高的样本主要为位于远离河流、海域的城市内部小型绿地如曲阳公园(1.00)与森兰体育公园(1.00),部分外环绿地的碎片状绿地均匀度同样较高,如高东生态园(0.99)与华夏公园(0.97)。从绿地类型看,外环结构性绿地均匀度均值最高(0.93),其次为沿海滨江(0.91)和集中建设区绿地(0.90)。由表2可知,沿海滨江与集中建设区绿地的鸟类物种相似性较高,而外环结构性绿地分别与集中建设区绿地以及沿海滨江绿地鸟类物质相似性较低。
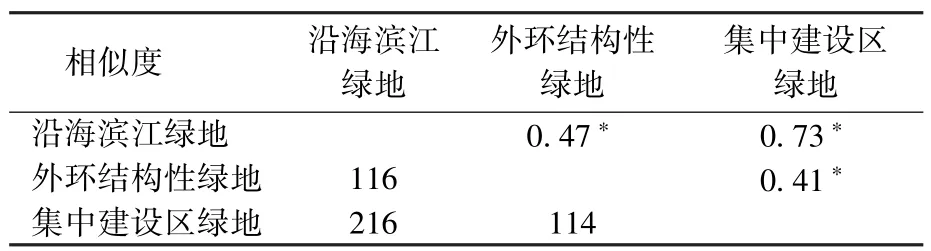
表2 不同类型绿地鸟类相似度
3.2 影响鸟类多样性绿地因子的边际效应
BRT结果显示(图3),GLPI整体贡献率最高(24.20%),HQ、WBR与WDIS接近,NDVI贡献率最小(9.90%)。
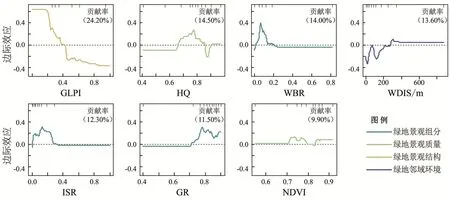
图3 影响因子对鸟类多样性贡献率及边际效应
绿地景观组分因子贡献度依次为WBR、ISR与GR。其中,WBR对鸟类多样性的贡献度为14.00%。当其介于0.00~0.20时,WBR增加可以促进鸟类多样性;超过0.20后,WBR继续增加则对鸟类多样性产生负面影响。ISR对鸟类多样性的贡献度为12.30%,当其处于0.00~0.33时对鸟类多样性呈正面影响,在超过该区间后呈负面影响。GR贡献度为11.50%,其曲线自0.71开始上升,当超过该阈值后,GR提升可以促进鸟类多样性。
绿地景观质量因子贡献度依次为HQ、NDVI,其中HQ贡献度为14.50%,当HQ在0.66~0.92时对鸟类多样性提升具有显著促进作用,在0.88时达到最高值后呈下降趋势,在0.95时达到谷值后开始上升,最终在1.00后不再变化。NDVI对鸟类多样性的影响并不显著,曲线波动较为平缓。整体而言,NVDI提升有利于鸟类多样性提升。
绿地景观结构因子GPLI贡献度为24.20%,是影响鸟类多样性的关键因子。当GLPI在0.00~0.40时,GLPI增大对鸟类多样性呈正面影响,超过0.40后不利于鸟类多样性提升。
绿地邻域环境因子WDIS贡献度为13.60%。当WDIS小于260m时,远离水体将对鸟类多样性产生负面影响,在WDIS达到300 m后对鸟类多样性的影响不再变化。
4 讨论
绿地景观组分对鸟类多样性贡献度整体低于景观结构与质量。较高的GR是提升鸟类多样性的重要因素,这与多项研究结论相似[24,26]。此外,广阔水域能够吸引更多鸟类[32],而过高的WBR会挤压鸟类栖息、隐蔽空间,不利于鸟类多样性提升,这与黄越[33]研究结论一致。值得一提的是,本研究发现当绿地具有一定比例不透水面时,可以在一定程度促进鸟类多样性,Liu等[24]认为ISR对鸟类多样性产生显著的负面影响的结论不同,其原因可能在于城市化地区周围绿地具有更高景观异质性,能为鸟类提供多样栖息空间和食物来源。
Dehuan等[34]验证鸟类多样性与HQ之间未呈现显著相关性,而Matthies等[35]则认为提升栖息地质量能够让原本在面积上占劣势的小型绿地具有较高鸟类多样性。本研究中较高HQ能够一定程度提升鸟类多样性,而当HQ在0.91~1.00时与鸟类多样性呈负相关,结合上海实际情况,该结果与外环绿带存在一定数量拥有较高生境质量、但因规模较小而使鸟类多样性较低的绿地样本有关。
绿地景观结构是本研究影响鸟类多样性最关键因子,当GLPI小于0.40时,鸟类多样性与GLPI呈显著正相关,大面积植被斑块可以有效降低人为因素的干扰[36]。而当GLPI超过阈值后,鸟类多样性则与景观破碎度呈正相关,表明过于单一的生境并不利于鸟类多样性提升,这一特征与Camargo等[37]结果相似。此外,鸟类多样性在一定程度随着栖息地与水体距离的减少而增加,该特征与Liu等[24]和Yang等[26]的研究结论一致。
综上,未来研究应关注以下问题:1)尽管通过公众科学项目平台获取的鸟类观测信息数据能一定程度改善实地调研的局限性,鸟类监测数据也会受观测者个人偏好、地域限制、数据重复等影响,因此可能会与真实数据存在一定偏差,未来可对鸟类数据集进行更精确的筛选与核对。2)未来可以关注景观尺度下高效串联城市鸟类栖息地、改善并提升城市绿地网络的连通性,协调与整合城市生态廊道与鸟类生境廊道,以响应城市生物多样性提升与生态安全格局构建的迫切现实需求。
5 结论
本研究有效揭示了上海主城区鸟类多样性与绿地环境的关系:1)以物种而言,雀形目与鸻形目鸟类占据主城区鸟类总数的6成以上。以生活型而言,上海主城区数量最多为鸣禽,广泛分布于各类城市绿地;陆禽、猛禽与攀禽偏好生境质量较高绿地,对水环境依赖度较低;涉禽与游禽偏好湖泊海域、沿江湿地等,对近水环境有较高要求。2)上海主城区鸟类多样性格局呈现显著空间异质性,近海绿地、沿江绿地、大型公园、部分外环绿地呈现较高鸟类多样性格局;城市综合公园鸟类丰度与多度同样较高。3)最大绿地斑块指数对上海主城区鸟类多样性贡献度最高,其次为生境质量与水体占比。同时,具有一定比例的不透水面绿地能在一定程度促进鸟类多样性,过高水体占比不利于鸟类多样性提升。
