中层鱼昼夜垂直迁移研究进展
2023-09-11江艳娥徐姗楠陈作志朱江峰
田 翰,江艳娥,张 俊,徐姗楠,陈作志*,朱江峰
(1.中国水产科学研究院南海水产研究所 农业农村部外海渔业可持续利用重点实验室,广东 广州 510301;2.上海海洋大学 海洋科学学院,上海 201306;3.南方海洋科学与工程广东省实验室,广东 广州 511548)
引 言
中层鱼(mesopelagic fish)是指栖息在深海中上层(200~1 000 m)的小型鱼类,游泳能力较弱,广泛分布于世界各大洋(图1)[1-2]。通过拖网捕捞和水声学勘测对中层鱼资源的评估值约为5~19.5×109t[3-4]。此外,中层鱼被认为是深海散射层(Deep Scattering Layers)的主要游泳生物,深海散射层的时空分布与海洋生态系统动力及环境关系密切,可用于海洋生态环境监测[5-6]。中层鱼的营养级水平较低,主要摄食浮游动物和包括同类在内的其他体型小的鱼类,同时作为高营养级掠食动物的重要食物来源,连接起海洋食物网的上、下行食物关系,是物质转换和能量传递的枢纽,对维持海洋生态系统的稳定至关重要[1,7]。
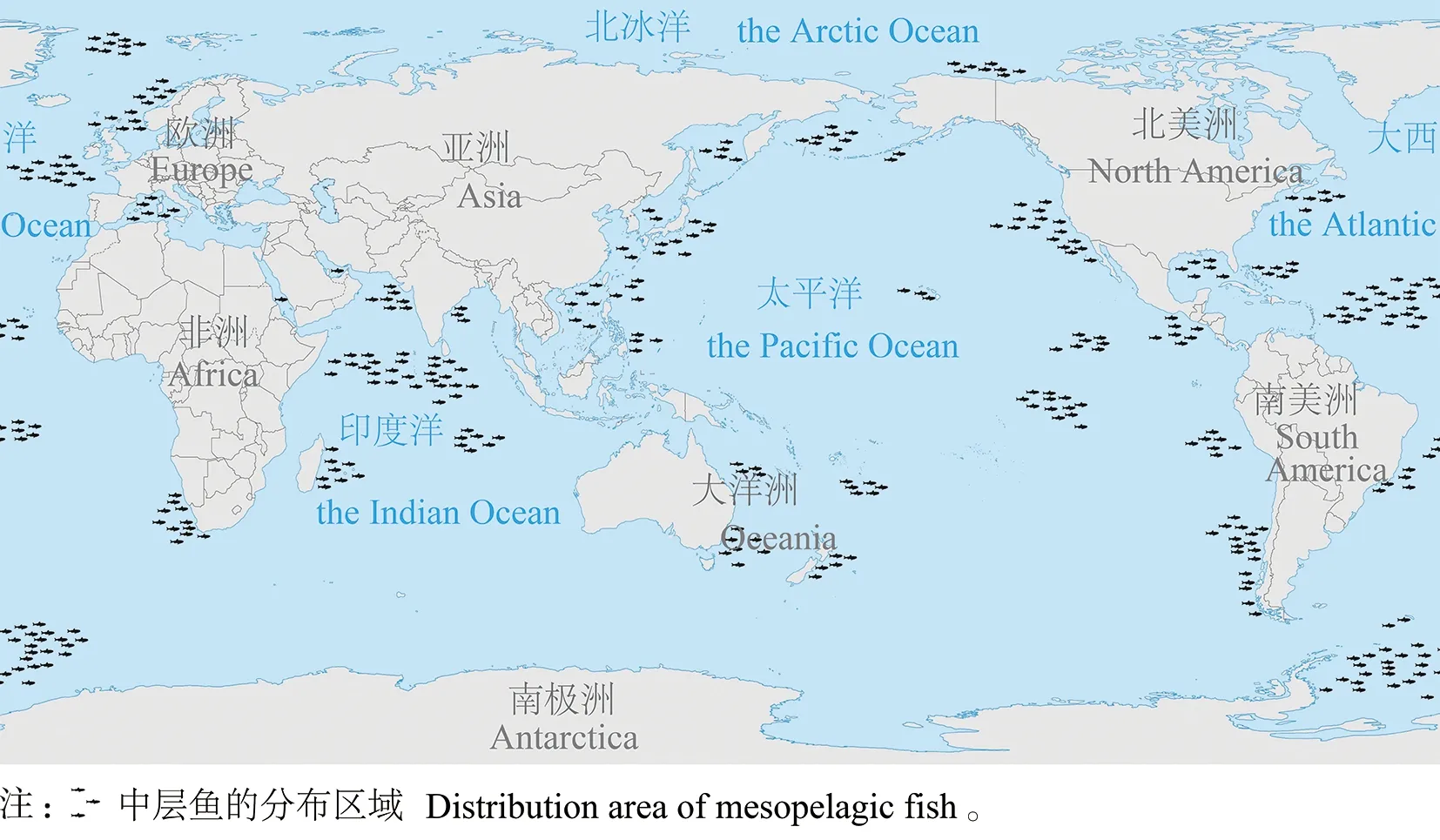
图1 中层鱼全球分布格局
昼夜垂直迁移(diel vertical migration,DVM)是大部分中层鱼种类的生存模式,是一种以日周期进行的昼沉夜浮垂直方向的长距离运动[8-9]。通过DVM,中层鱼以代谢的方式或以自身作饵料为深海带入碳源,对全球海洋生态系统的碳循环和氧气的动态平衡作出重要贡献[10]。
在全球近海传统渔业资源衰退的背景下,中层鱼资源逐步得到关注,有关中层鱼的研究逐步开展,涉及其生物学、资源分布、生态作用、食用价值等[6-8,10-15]。中国中层鱼的研究尚处于初始阶段且主要集中在南中国海,少数研究对南中国海北部陆坡海域、中部海域以及西沙海域中层鱼群落组成和生物学特点进行了报道[9,11-19],而有关中层鱼DVM的研究仅限于南中国海北部陆坡海域[19]。对中层鱼DVM的研究将有利于增加对中层鱼类群落和中上层海洋系统生态功能的认识。
1 中层鱼的昼夜垂直迁移
中层鱼的DVM可通过对深海散射层的连续声学映像进行直接观察,对深海散射层而言,约有95%的生物具有DVM行为,其中以中层鱼和浮游动物为主[6,8]。PEARCY等[8]依据中层鱼是否进行DVM,将中层鱼划分为非迁移物种和迁移物种。一般认为非迁移物种普遍栖息于深海,在近表层几乎无法发现他们的踪迹,例如长丝角鮟鱇科(Gigantactinidae)、鼠尾鳕科(Macrouridae)鱼类,但它们只占中层鱼中较少的比例。迁移物种涵盖了大部分种类的中层鱼,迁移者以灯笼鱼科(Myctophidae)鱼类占比最高[2,19,26]。
中层鱼的DVM行为影响物种在昼夜垂直空间上栖息水层的分布差异。多数迁移种类白天栖息在深海,于黄昏时间(约为日落前后1 h),凭借其主动游泳能力,冲破几百米垂直路径上存在的盐跃层和温跃层的屏障作用,以10~200 m/h不等的速度进行持续数小时的垂直迁移运动达到较浅水层(深度范围主要集中在0~200 m)。迁移种类在近表层的活动会持续整个夜晚,并在黎明时间(约为日出前后1 h)完成到深海的下潜[20,21,26-28]。
对迁移群体而言,依据迁移前后栖息水层是否重合,可为不完全迁移种类和完全迁移种类[4]。不完全迁移的物种迁移前后栖息水层存在重合,如钻光鱼科(Gonostomatidae)、褶胸鱼科(Sternoptychidae)、孔头鲷科(Melamphaidae)的大部分种类,这些种类也称为弱迁移鱼类,通常生活在深水层[4,28-30]。完全迁移物种是具有强迁移能力的鱼类,迁移前后栖息地完全分离,例如多数灯笼鱼科鱼类[4,20,26,28-30]。完全迁移的鱼类也存在垂直迁移路径长短的差异,以多数灯笼鱼为例,其迁移的垂直路径从深水层一直到海表,而蝰鱼科(Chauliodontidae)鱼类的迁移并不会上浮至近表层[28-31]。除了昼沉夜浮的迁移种类之外,少数种类则是反向垂直迁移,它们在黄昏沉降,黎明上浮[1,32](图2)。DVM行为的起止时间在不同水深、季节和不同群体之间均存在差异[1,18,31,33]。迁移群体在进行DVM时具有较大的主动选择性,种间差异导致DVM起始和结束的时间、深度不同;即使同一群体,前后两次迁移的深度和时间也会存在差异[1];此外,某些种类中层鱼可改变迁移强度,以适应自身生理需求及海洋环境的变化,例如智利串光鱼(Vinciguerrianimbaria),在性腺发育期间会滞留在海洋表层数天[25]。
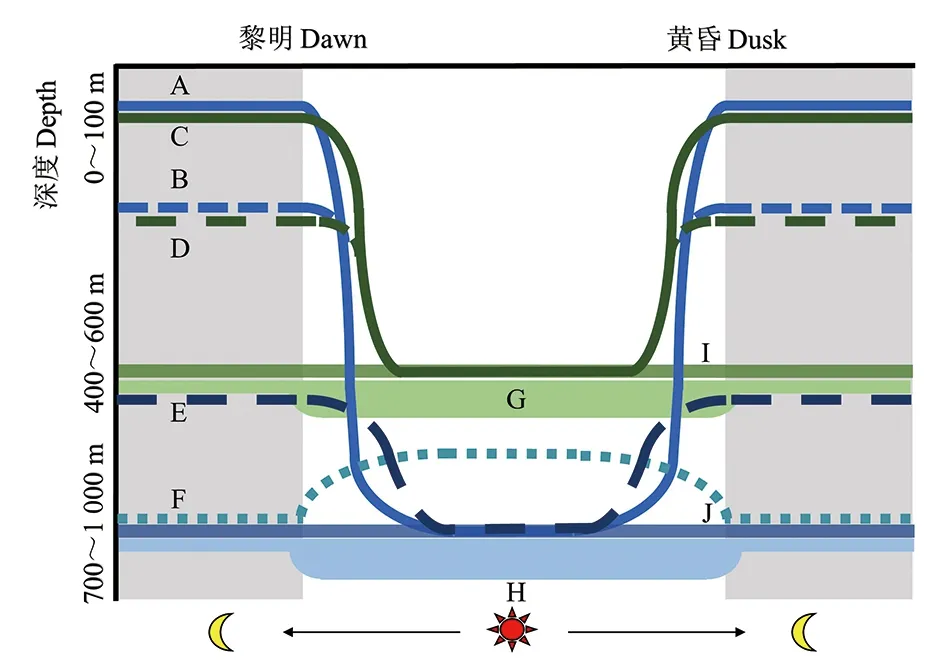
图2 中层鱼的昼夜垂直迁移行为
中层鱼的DVM在全球各海域皆有记录,最早的观测记录可追溯到1965年,近年来,有关中层鱼DVM的研究已在全球不同海域陆续展开,200~1 000 m水层,是国内外研究DVM的重点水层[1-2,4,6,8,18-21,29-30]。目前,对中层鱼DVM行为的观测有声学观察和拖网捕捞两种主要方式[1,4,11-12,17,29,34]。表面散射层和深海散射层在昼夜存在着剧烈的波动,具体表现为表面散射层在白天密度小、厚度薄,夜晚密度大、厚度宽,深海散射层的变化情况与表面散射层相反,在黎明和黄昏,能明显看到深海散射层和表面散射层之间形成连接的通路。拖网捕捞是证明DVM的间接方法,即根据样本获得时的作业水层及时间间接推算中层鱼的DVM[7,10,15,19,32]。然而,声学映像无法区分鱼类和其他生物,拖网捕捞可以确认具体的迁移种类,但存在网目选择和鱼类对网具的逃逸而导致调查结果偏小的问题。此外,水下光学设备观察、稳定同位素技术、有机碳浓度测定、不同水层捕食生物的胃含物分析、寄生虫检测等方法也直接或间接地用于中层鱼的DVM的观测[18,33,35]。
2 昼夜垂直迁移影响因素
DVM是中层鱼种群的生存策略,是一种完全主动的行为[35-40]。中层鱼DVM的动机是获取食物和躲避敌害,环境因素也会影响DVM进程[38-40]。中层鱼迁移的垂直迁移受生物因素和环境因素的影响,在其垂直迁移通路上不同深度水层的捕食者,以及不同性质水团的生境,均给中层鱼的DVM带来极大的风险[1,36-39]。
2.1 生物因素
2.1.1 饵料生物
中层鱼营养水平较低,约在2~4之间,主要捕食浮游动物,但同时也会捕食较大的鱼类,一般认为,大部分种类中层鱼倾向捕食小于其口径的猎物,是依靠视觉搜寻猎物的机会主义捕食生物[2,20,28,41]。中层鱼的浮游动物食性的推论主要来自对捕捞样本的胃含物的解剖观察和浮游动物以及中层鱼DVM间极高的同步性。OLIVAR等[2]和SASSA等[42]的研究认为存在迁移行为的中层鱼在夜间表层和白天深层均会捕食,二者相较而言,在夜间表层从深海迁移上来的中层鱼会进行更大规模觅食,此外,PEARCY等[8]认为DVM所消耗的能量占到中层鱼每日日获得能量总量的67%。从种群生存的角度来看,如果中层鱼每天在深层海域获得的能量足以维系其生长和发育,DVM所需要付出的巨大代价对其来说并非是必要的。这表明对有DVM习性的种类而言,仅靠在深海捕食获取的能量不足以支持其日生长所需,发生在迁移过后夜间海表的捕食活动为这些鱼类提供了远超过DVM消耗的能量,支持了索饵是诱发中层鱼进行DVM的关键的推论。
2.1.2 敌害生物
中层鱼面临的捕食压力主要来自包括同类在内的其他中层鱼、捕食性鱼类、头足类、海洋哺乳动物和鸟类,大都为视觉捕食生物,多项研究表明这些捕食生物的昼夜活动和中层鱼的DVM存在相关性,例如部分头足类和金枪鱼类在夜间的栖息水层会临近海表,白天则在较深海域出没[20,28,40,43-44]。在白天,海洋表层光照充足,视野开阔,大型捕食生物活动频繁,浮游动物下潜至深层海域。中层鱼在此时普遍栖息于海洋深层,深海低温低氧的环境限制了对温度及溶氧需求较高的高营养级捕食生物的活动,从而使中层鱼在有食物保障的同时,极大地降低了被天敌捕食的风险。夜间海表,大型捕食生物活动减少,来自不同深度的浮游动物大量聚集,食物充沛,中层鱼会在此进行8~10 h的摄食活动。此外,夜间表层被认为是多数种类中层鱼进行产卵的水层[45]。DVM使中层鱼在满足自身食物需求的条件下最大程度地减少被敌害生物捕食的可能从而保障了种群的生存。
2.2 环境因素
2.2.1 光
光是对中层鱼DVM影响最大的环境因素,起到控制、启动和定向的驱动作用,决定了DVM起末的时间、深度及迁移速度[26,46-48]。中层鱼被认为具有趋弱光、避强光的特点[26]。在白天海表,光照强烈,紫外线的辐射量大,不利于中层鱼在海表的聚集。中层鱼主要分布在散射层及附近水层,而在阴天或光线较弱时,海表零星出现中层鱼的分布,此时中层鱼在深海的垂直分布水层比晴天更浅,分布数量更大,这一现象在多项研究均有报道[4,20,26,39,48-54]。这说明光线强弱对中层鱼DVM的起止深度有直接影响,控制着中层鱼在深海的垂直分布。夜间从海表进入海洋的自然光主要来自月球对太阳光线的反射,相较于白天阳光的直射,夜间的光线较弱。在夜间、阴天时中层鱼在海表聚集得更加密集,深层的中层鱼上浮水层更浅,这一特点在浮游动物中同样被发现[51-52]。此外,月相同样会影响夜间光线的亮度,现阶段有关中层鱼DVM和月相的研究较少,作用原理尚存在争议。虽然部分研究观察到在新月期间,夜间海表物种丰度较高,深层的种类保持在较深的水层,在满月期间夜间海表物种丰度较低,深层的种类保持在较浅的水层,表明月相和中层鱼DVM存在相关性,但在这些研究中,月相只是诸多环境因子中的一个,似乎并没有对中层鱼DVM起到决定性的影响[53-57]。总体而言,月相变化对多数种类中层鱼DVM的影响不显著,只有少数中层鱼的DVM和月相存在明显的相关性。
光驱动的另一重要原因是中层鱼的上下营养级生物活动均受光照强弱的影响。多数中层鱼的捕食者依靠视觉捕食,光线强弱直接影响捕食者的活跃程度和捕猎成功率[58-59]。此外,光照强弱间接影响到海域营养水平的高低,低营养海域通常伴有水域清澈的特点,高营养海域则浊度较大,因此相较于高营养海域,低营养海域的中层鱼会迁移至更深水层[1]。
2.2.2 温度和盐度等因素
除了光照的影响之外,温度和盐度对中层鱼DVM影响较大。海洋中广泛存在不同理化性质的水团,决定了海洋环境的高度空间异质性,并直接影响不同空间分布的中层鱼的物种组成[25,60]。
基于不同种类中层鱼对环境温度的耐受差异,BUTLER等[31]认为温度是造成DVM差异的重要环境因素,CAVAN等等[40]的研究同样认为海洋环境中剧烈的温度变化会限制中层鱼的垂直迁移。拥有更大温度耐受区间的鱼类拥有更广泛的迁徙范围,10℃的温差耐受区间足以支撑某些种类进行几百米甚至数千米垂直距离的移动[19,61-62],WANG等等[19]的研究进一步发现某些种类迁移群体迁移起止所处不同水层的最大温差可以达到20 ℃。蝰鱼属(Chauliodus)鱼类在热带和温带的种群有着不同的垂直迁移模式,热带群体几乎不会迁移至500 m以浅的海域,而在温带海域可以在200 m左右的深度发现它们的踪迹[31]。NETBURN等等[61]对极地锋面两侧中层鱼DVM的研究结果表明,锋面较冷的一侧迁移的种类更多;而例如圆罩鱼属(Cyclothone)等不进行垂直移动的鱼类则对温度变化不敏感[40]。盐度则从两个方面影响中层鱼的迁移:1)不同水体的盐度动态变化与中层鱼的盐度耐受;2)盐度通过影响水体的营养水平进而影响水体浑浊程度。研究认为,盐度耐受范围更大的中层鱼种类能进行路径更长的迁移,低盐度区域的中层鱼DVM的幅度更大[36-37,60]。
HAURY[28]认为中层鱼DVM行为有明显的季节相关性,迁移群体数目在夏季明显多于秋季,与WANG等[19]在南中国海的结论一致。鱼鳔的有无同样被认为会影响中层鱼的DVM,目前的研究认为有鳔类的迁移幅度通常小于无鳔类[63]。此外,不同种类中层鱼对溶解氧的耐受范围差异可能也是造成DVM差异的原因之一,但限于调查的广度,溶解氧对中层鱼DVM的影响仍需进一步验证[50]。
3 昼夜垂直迁移对海洋生态系统的影响
3.1 对海洋碳汇的影响
在DVM过程中,中层鱼充当了“生物碳泵”的角色,促进了能量的流动,改变了海洋中有机碳的空间分布,促进了全球碳汇进程[10,23,41,64]。在中层鱼介导的碳汇中,碳以溶解性有机碳(Dissolved Organic Carbon)和颗粒性有机碳(Particulate Organic Carbon)的形式进入中层海洋(图3)。溶解性有机碳的产生主要来自表层浮游动物和细菌对有机碳的矿化作用,颗粒性有机碳则主要由中层鱼排泄的固体颗粒物以及尸骸构成[40,66]。中层鱼DVM对有机碳的搬运作用,减少了颗粒性有机碳在表层海洋的分解,给中层海洋提供了可以利用的碳源,是中层及更深层海域颗粒性有机碳的主要来源之一[40-41]。ZHANG等[18]观察到同一海域夜间海表和白天深层海域溶解性有机碳浓度的同幅度升高,表明深海可能发生了颗粒性有机碳向溶解性有机碳的转化,也说明有机碳的垂直变化可以验证中层鱼的DVM。部分研究表明,中层鱼通过DVM进入深海后的呼吸消耗达到了表层海域生产力的10%。“深海呼吸”是深海中存在巨大的微生物呼吸作用,但产生这一现象的原因尚不清楚,中层鱼在深海的代谢可能是这一现象形成的原因之一[41,65]。中层鱼碳汇模型的建立使中层鱼对碳汇的贡献可以被有效评估,其碳传递效率被认为主要与中层鱼个体大小、游泳能力、丰富度以及环境因素有关,相关研究表明,世界各地中层鱼传导的生物碳占总碳量的8%~40%不等[18,41,66-67]。研究表明,中层鱼DVM对碳的搬运作用促进了海底碳床的积累,稀释了空气中的二氧化碳浓度,对缓解温室效应具有一定贡献[28,65]。
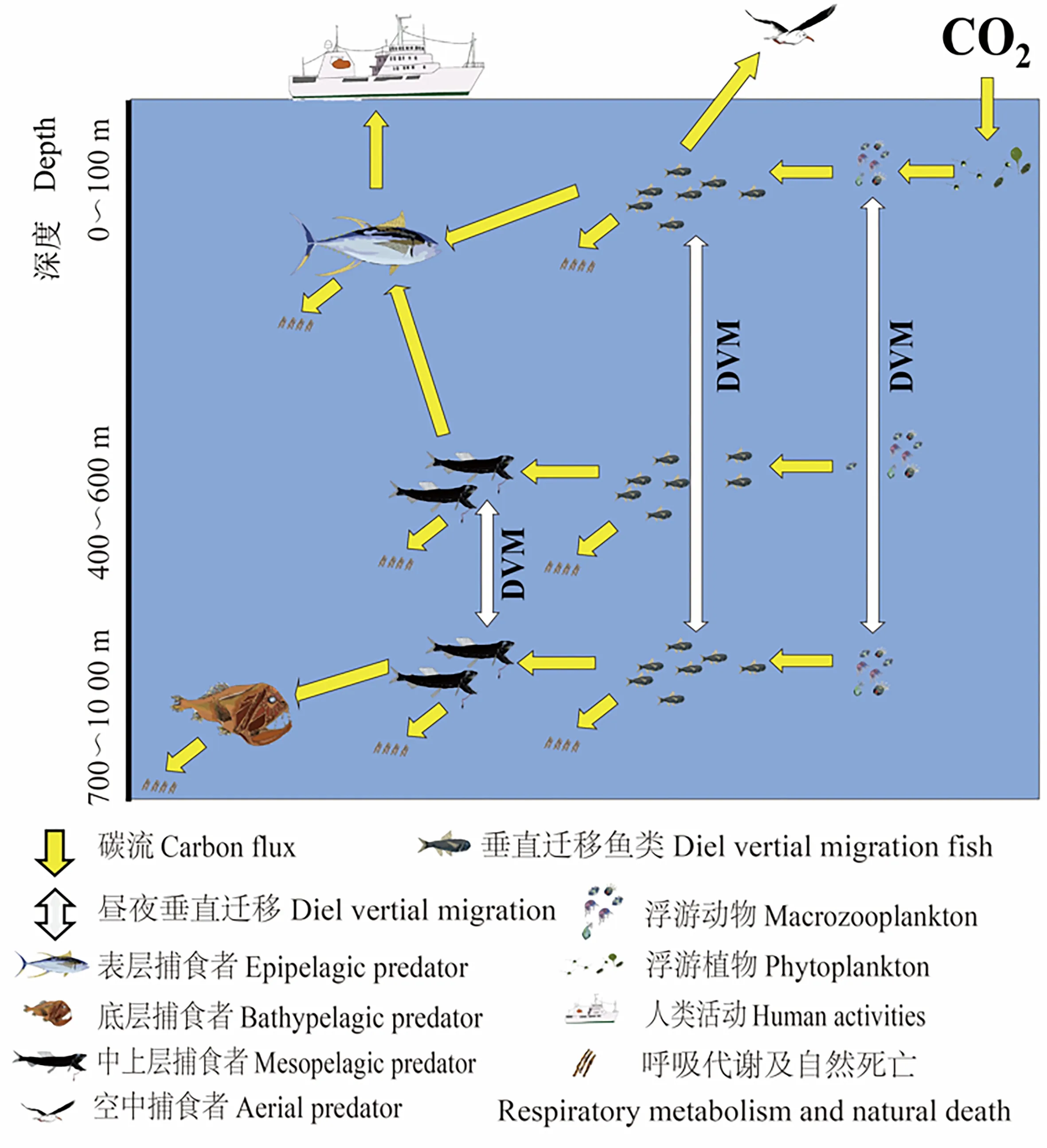
图3 中层鱼类的垂直迁移和营养相互作用(据[21]修改)
3.2 生物空间联系
中层鱼通过DVM建立了多个水层在时间、空间和生物方面的联系,是海洋食物网的核心枢纽,维持了深海生态系统的稳定[6-7,39]。CAVAN等[40]研究认为,相较于上层海洋,中层鱼对中层海洋各生态因素空间联系的贡献,尤其对中层海洋生物群落结构的影响更为重要。阳光的匮乏使中层深海生物生长所需的能量大量依赖上层海洋的注入,中层鱼则是最重要的能量运输者[23,41,64]。
中层鱼白天在不同深度水层垂直分布,存在种群的空间隔离,夜间海洋上层狭窄空间的大量聚集会打破这一限制,KLEVJER等[6]认为,原本因白天栖息水层不同而没有上下级营养关系的鱼类在夜间海表会构建新的捕食关系,除此之外,整合效应将会极大地丰富种间交流。有报道指出,中层鱼的DVM使自身成为部分深海寄生虫的理想中间宿主,寄生虫最终通过食物网进入上层海域的鲸类和其他高营养级动物的体内[35]。
4 展望
中层鱼资源具有重要的开发潜力,在全球渔业资源衰退的背景下,逐渐受到广泛关注[1,4,20]。现阶段,对中层鱼资源的开采尚没有形成完整体系,存在着能源消耗大、产品价值低等诸多问题,未来,开采及加工方式的优化升级将提升中层鱼资源的利用价值[1,20,62]。目前进行中层鱼研究的方式主要为中层拖网调查和声学勘测,然而两种主要的调查方式均存在弊端;拖网捕捞中鱼类的逃逸问题、网目的选择性、如何剔除其他游泳鱼类、采样深度与鱼类群落栖息深度的动态变化等[2,13,42]。在声学观测中,如何有效区分有鳔类鱼类和无鳔类鱼类的差异,排除其他游泳生物,例如浮游动物、水母(jelly fish)等其他生物对音频信号反射的干扰等,是未来中层鱼资源勘测需要解决的问题。由于中层鱼个体相对较小以及现阶段捕捞方式几乎无法保证样本的存活[1,20],极难应用鱼类监测跟踪技术,使中层鱼个体的DVM难以被直接观测。中层鱼的DVM行为主要通过对比拖网样本的水层分布差异并结合声学映像的整体趋势推断得出,水下光学设备提供的影像资料虽可以提供中层鱼DVM最直接的证据,但因成本高昂和深度的限制难以普及推广。现阶段常规的拖网捕捞作业水平和声学勘测技术限制了继续向更深水层的调查,因此,DVM的深度极限目前尚未有定论,中层鱼的DVM是否会影响1 000 m以下的深海生态系统是未来深海研究值得考虑的问题。
了解迁移种类中层鱼的生物学信息是了解中层鱼DVM及其生态作用的前提,现阶段对中层鱼生物学的研究主要涉及年龄发育、食性组成和身体构造等,仍有大量空白亟待补充[1,4,17,20]。部分生物学特征被认为与中层鱼DVM存在相关性,例如,迁移群体中有鳔类比无鳔类占比更大;从肌肉组成的比例来看,红肌/白肌比例更高的种类更倾向于进行DVM[1,19-20]。基于对中层鱼DVM发生时间的掌握,BADCOCK[64]认为黄昏和黎明是研究中层鱼迁移的最佳时段,而对中层鱼垂直分布的研究应尽量集中在白天。中层鱼DVM过程的连续性至今仍未有定论,即便是长距离完全迁移种类,没有直接证据证明他们在迁移过程中是否会短暂滞留某一水层进行捕食,这些因素限制着对DVM行为的认识和理解。若捕食行为存在于迁移过程中,我们需要重新考虑中层鱼的迁移行为如何影响中层海洋的食物联系。中层鱼在迁移过程中存在的捕食、寄生关系、繁殖、代谢行为,以及作为“生物碳泵”和“深海呼吸”的关键生物对深海生态系统乃至生物圈的影响将是生态学研究者们进一步研究的问题[10,23,41,55,64]。例如DAVISON等[68]发现,由中层鱼深海呼吸消耗推算得出的区域海域初级生产力的理论值比实际结果更高,这一发现可能催化人类对海洋能量生产与消耗新的认识。
近年来,南中国海发现了资源量巨大的中层鱼资源,具有潜在的开发利用价值[9,11-15]。目前,南中国海中层鱼研究基础较为薄弱,只掌握了少数部分海域中层鱼群落特征和DVM习性,由于南海深度相对较浅,南中国海中层鱼的栖息水层主要为200~800 m,因此南海中层鱼DVM的研究也将集中在这一深度[19]。中层鱼DVM生态效益的研究是一个庞大而繁琐的工程,鱼类资源与物理、化学、地理、大气等多学科相互渗透将会推动海洋领域的研究走向整体化和合作模式,除了需要进一步加强对中层鱼资源的科学勘测及开发,更需要借助政府政策支持以及国际养护管理和区域协同合作。未来,我国可从中层鱼的生态习性、数量及分布等基础研究起步,再到聚鱼探捕、渔情预报等关键技术进行深入研究,提升我国对这一海洋生物资源的认识,培育海洋生物新兴产业,同时为该资源的合理开发利用和保护提供科技支撑。
