盐胁迫下乌桕无性系叶片的比较蛋白组学研究
2023-09-09隋德宗王保松
隋德宗 王保松
(江苏省林业科学研究院,南京 211153)
土壤盐渍化是我国农林生产面临的主要生态环境问题之一,严重影响种子萌发,限制植物生长发育,制约现代农林业发展[1-2]。据统计,江苏省沿海滩涂总面积高达68.7万hm2,其中潮上带29.5万hm2,并以每年1 267 hm2的速度淤长[3]。沿海滩涂作为江苏省重要的土地后备资源,是林业发展极具潜力的区域。沿海滩涂地区土壤含盐量高,立地条件差,适生树种少,优良耐盐林木种质资源的缺乏是制约沿海林业可持续发展的关键问题。对现有沿海乡土林木树种资源的挖掘改良是解决此问题的有效途径。
乌桕(Triadica sebifera)是江苏省沿海地区唯一有天然分布的彩叶大乔木,是集观赏、油用和生态应用于一体的多用途树种。乌桕树形高大,秋叶艳丽,观赏价值高,是长江流域重要的园林景观树种[4]。乌桕适应性强,既耐干旱瘠薄,又耐水湿和中度盐碱,可作为优质的沿海先锋造林树种[5]。目前,乌桕相关研究主要包括育苗繁殖技术[6]、表型多样性[7]、生长发育[8]、品种选育[9]、光合特性[10]、叶片呈色生理生化[11]、非生物胁迫响应[12-14]以及分子水平的分子标记开发[15]、基因功能[16]及转录组测序[17]等方面。此外,近期染色体水平的乌桕基因组图谱研究深入揭示了其多倍化、适应性强、种子不饱和脂肪酸及叶片中花青素生物合成机理[18]。伴随植物组学技术的不断发展,从基因组、转录组、代谢组、蛋白质组等的全面研究为深入揭示重要生物学问题提供了新的技术。然而,从蛋白质水平揭示乌桕参与盐胁迫响应的相关研究鲜有报道。
植物细胞内蛋白质组丰度的动态变化能够影响到植物体一系列生命过程。蛋白质组学是把一个物种基因组所表达的全部蛋白或一个体系中所有蛋白进行精准鉴定与定量。同位素标记相对和绝对定量(iTRAQ)是一种在肽段水平进行体外标记技术,结合串联质谱分析可准确定量不同样本中的蛋白质表达情况,具有灵敏度高、重复性好等特性,其在农林领域主要应用在植物生长发育与营养品质[19]、病害及生物防治[20]、抗逆胁迫机制[21-22]等方面。因此,进行乌桕蛋白质组学分析对揭示其逆境条件下蛋白响应盐胁迫研究具有重要意义。
利用组学技术在蛋白质水平探讨乌桕盐胁迫响应的分子机理,有助于利用现代生物技术结合常规育种技术培育出耐盐性优异的乌桕良种。本研究以课题组前期获得耐盐性具有明显差异的2个乌桕无性系[14]为研究材料,利用iTRAQ 技术鉴定出盐胁迫不同时期乌桕叶片的盐胁迫响应蛋白,通过对比分析,从蛋白质水平上初步揭示乌桕耐盐分子机理,为培育耐盐乌桕良种及林木耐盐机制研究提供科学依据。
1 材料与方法
1.1 研究材料
试验所用的乌桕盐敏感型无性系(salt-sensitive P18,SS18)和盐耐受型无 性系(salt-tolerant P21,ST21)为江苏省林业科学研究院通过嫁接繁殖的无性系苗。选取苗木健壮及生长状况较为一致的2年生株系,分为2组,每组选取6株作为生物学重复。在7 月份,按照每盆1 000 mL·d-1的0.4%NaCl 溶液进行浇灌处理,设置不同处理时间(0、24、72 h)。采集不同处理时间下的乌桕叶片(每组叶片样本重复3 次)迅速放入液氮中保存,用于后续蛋白质提取和生物学分析。
1.2 蛋白组学分析
样品送至南京捷倍思生物基因技术有限公司进行蛋白提取、含量测定、蛋白酶解与标记、肽段反向色谱分离和反向色谱-Triple TOF 检测。实验主要方法和步骤介绍如下。
1.2.1 样品蛋白质提取
采用三氯乙酸/丙酮沉淀法[23-24]进行乌桕叶片蛋白质提取。
1.2.2 蛋白质酶解
参 照Wu 等[25]使 用 的 蛋 白 滤 膜 辅 助 酶 解(FASP)法并进行部分调整,制取肽段。
1.2.3 iTRAQ 标记
使 用iTRAQ 试 剂 盒(iTRAQ Reagent-8 plex Multiplex Kit,AB Sciex,USA)对各样品进行iTRAQ标记,具体步骤参照试剂盒说明书。
1.2.4 肽段HPLC分离
冷冻干燥后得到的样品用100 μL Buffer A 溶液充分溶解。在Agilent 1200 HPLC 上进行SCX 分离,C18 色 谱 柱 购 至Michrom 公 司(California,USA),其主要参数为Poly-SEA 5 μm 300Å 2 mm×150 mm,检 测 波 长 为 紫 外215、280 nm。流 速0.3 mL·min-1。按照试剂盒说明书中的强阳离子交换色谱溶液时间表的间隔收集样品溶液,共计12管样品溶液,然后将所有溶液彻底冷冻干燥。
1.2.5 反向色谱-Triple TOF检测
用Nano-RPLC Buffer A 重新溶解冻干多肽样品,利用Eksigent nanoLC-Ultra™ 2D 系统(AB SCIEX,USA)的C18 预柱(100 μm×3 cm,3 μm,150Å)进行冲洗脱盐10 min,分析柱为C18 反相色谱柱(75 μm×15 cm,3 μm,120 Å),试验用梯度为70 min内流动相B 从5%升高到35%。质谱分析采用Triple TOF 5600系统(AB SCIEX,USA)结合纳升喷雾Ⅲ离子源(AB SCIEX,USA)收集数据。
1.2.6 数据分析
获得的实验数据采用 Protein Pilot Software v.5.0(AB SCIEX,USA)软件进行分析,其数据库来源于Uniport蛋白库。
1.2.7 差异蛋白筛选
差异蛋白筛选标准参考姜志磊等[24]方法,首先去除反库数据和比值为空白的无效数据,筛选出错误发现率(FDR)<1%且肽段数≥1条的可信总蛋白数量;再以可信蛋白为基础,差异1.5 倍以上(Fold change>1.5 或<0.67)且统计学ANOVA 检验P<0.05视为显著差异表达上调或下调的蛋白。
1.2.8 差异蛋白生物信息分析
实验结果筛选出差异蛋白之后,根据序列相似性将差异蛋白BLAST 到模式植物拟南芥(Arabidopsis thaliana)中,再进行GO 富集分析、京都基因与基因组百科全书(KEGG)通路分析及蛋白质-蛋白质相互作用网络构建分析(PPI)。
1.3 统计分析
基于蛋白表达丰度,进行Spearman 相关性分析,基于显著相关的蛋白,利用Cytoscape 软件(https://cytoscape.org/)构建蛋白-蛋白互作网络图。采用SPSS 21.0 软件进行统计学分析,用平均值±标准差(± SD)表示,统计结果为双尾检验,P<0.05表示显著统计学意义。
2 结果与分析
2.1 乌桕叶片响应盐胁迫的差异蛋白
通过iTRAQ 分析,在盐敏感型无性系SS18 和盐耐受型无性系ST21中分别获得3 374、2 725种蛋白,筛选出可信蛋白分别为3 187、1 831 种。与对照组(0 h)相比,0.4%NaCl 盐胁迫处理24、72 h 的SS18 叶片分别鉴定出53、253 种差异蛋白,其中有53 种共同差异蛋白;0.4% NaCl 盐胁迫处理24、72 h 的ST21 叶片分别鉴定出54、68 种差异蛋白,其中有53种共同差异蛋白。
对于SS18叶片响应盐胁迫的已知差异蛋白有31 种,其中包括21 种上调蛋白和10 种下调蛋白。上调蛋白主要涉及能量代谢(丙酮酸脱氢酶E1α亚基、紫外线受体蛋白、β-己糖胺酶、丝氨酸/苏氨酸蛋白磷酸酶)、蛋白翻译(延伸因子-1α、核糖体L9、40S 核糖体蛋白S12、核糖体S10、酰基转运蛋白、真核翻译起始因子5A)和氧化应激(过氧化氢酶、过氧化物酶4、硫氧还蛋白)等代谢过程。下调蛋白主要涉及叶绿体光合作用(叶绿体铁氧还蛋白-NADP 还原酶、叶绿素a-b 结合蛋白、ATP 合成酶β亚基)、能量代谢(丙二酸辅酶A:ACP 转酰基酶、丙酮酸激酶)和蛋白降解(氨基酸氧化酶、泛素羧基末端水解酶)等代谢过程。可以看出SS18 在盐胁迫条件下,其光合作用、能量代谢及氧化应激过程的相关蛋白变化较为显著。
对于ST21 叶片响应盐胁迫的已知差异表达蛋白有33种,其中包括16种上调蛋白和17种下调蛋白。上调蛋白主要涉及光合作用(光系统ⅠP700 叶绿素a 载脂蛋白A2、光系统Ⅰ P700叶绿素a载脂蛋白A1、光系统Ⅱ CP43 反应中心蛋白)、氧化应激(过氧化氢酶、硫氧还蛋白)、翻译调控(延伸因子-1α)等代谢过程。下调蛋白主要涉及三羧酸循环(果糖二磷酸醛缩酶、磷酸甘油酸激酶、谷氨酰胺合成酶、核酮糖二磷酸羧化酶)等代谢过程。可见ST21 光合作用及氧化应激等相关蛋白上调表达,而糖代谢相关蛋白下调表达。
盐胁迫条件下,在SS18 和ST21 叶片中均上调表达的蛋白有4种,分别为过氧化氢酶、延伸因子-1α、含H+-ATPase_c 结构域的蛋白和硫氧还蛋白,这些蛋白在耐盐胁迫响应过程中可能作为乌桕耐盐的重要潜在靶标蛋白。
2.2 GO功能注释分析
按照生物过程、细胞组分和分子功能3方面进行差异蛋白的GO 功能注释分析。如图1A 所示,SS18在盐胁迫24 h时的差异蛋白主要涉及小分子代谢过程、氧化还原辅酶代谢过程、有机酸代谢过程、辅酶代谢过程和辅因子代谢过程、烟酰胺核苷酸代谢过程等;而SS18 在盐胁迫72 h 时的差异蛋白主要涉及光合作用、前体代谢物和能量的产生、光合作用与光反应、对非生物刺激的响应、对温度刺激的响应等(图1B)。
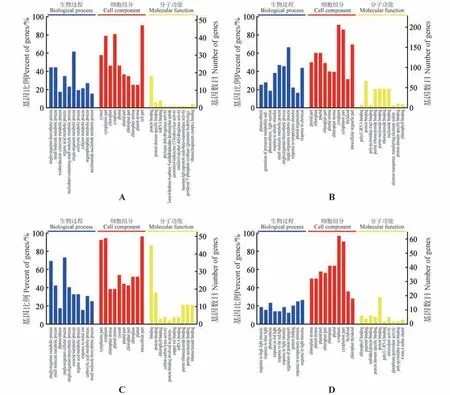
图1 乌桕叶片在盐胁迫下差异蛋白的GO富集分析A.与SS18未处理(0 h)相比,盐胁迫处理24 h后SS18叶片差异蛋白的富集;B.与SS18未处理(0 h)相比,盐胁迫处理72 h后SS18叶片差异蛋白的富集;C.与ST21 未处理(0 h)相比,盐胁迫24 h 后ST21 叶片差异蛋白的富集;D.与ST21 未处理(0 h)相比,盐胁迫处理72 h 后ST21叶片差异蛋白的富集Fig.1 GO enrichment analysis of DEPs in T. sebifera leaves under salt stress A.GO analysis of DEPs in SS18 leaves after 24 h salt stress,compared with 0 h no salt stress;B.GO analysis of DEPs in SS18 leaves after 72 h salt stress,compared with 0 h no salt stress;C.GO analysis of DEPs in ST21 leaves after 24 h salt stress,compared with 0 h no salt stress;D.GO analysis of DEPs in ST21 leaves after 72 h salt stress,compared with 0 h no salt stress
对于盐耐受型无性系ST21,在盐胁迫24 h 的叶片差异蛋白主要富集在光合作用、氧酸代谢过程、有机酸代谢过程和羧酸代谢过程等生物代谢过程(图1C);ST21 在盐胁迫72 h 的叶片差异蛋白主要富集在对高光强度的响应、对蓝光的响应、光合作用、对红光的响应、对远红光的响应、对光强度的响应、对温度刺激的响应、对光刺激的响应、质子转运的调节和蛋白质折叠等生物代谢过程(图1D)。
可以看出,在盐胁迫刺激下,乌桕叶片中的差异蛋白多涉及光合作用过程,尤其在胁迫处理72 h时,表现更为明显。
2.3 KEGG代谢通路分析
为了进一步了解乌桕叶片对盐胁迫响应的差异蛋白在代谢途径中的变化,将筛选出的差异蛋白映射到KEGG 代谢通路上,进行通路注释和富集分类分析,总计富集到36 个代谢通路(图2)。盐胁迫24 h,SS18叶片表达差异蛋白主要富集到6个代谢通路(图2A),分别是丙酮酸代谢、碳代谢、糖酵解/糖异生、光合作用、鞘糖脂生物合成—神经节苷脂系列和糖胺聚糖降解;盐胁迫72 h,SS18叶片表达差异蛋白富集到14 个代谢通路(图2B),分别为光合作用、光合有机体中的碳固定、碳代谢、代谢途径、乙醛酸和二元酸代谢、糖酵解/糖异生、氨基酸生物合成、磷酸戊糖途径、α-亚麻酸代谢、半乳糖代谢、醛酸代谢与抗坏血酸、缬氨酸、亮氨酸和异亮氨酸的生物合成、内质网的蛋白质加工和脂肪酸降解。上述结果可见盐敏感型无性系SS18 在盐胁迫初期 (24 h),糖类代谢和光合作用均有所增强,随着盐胁迫时间的延长(72 h),氨基酸代谢、脂肪酸代谢及有机酸代谢变化较大,这些可能在其参与盐胁迫响应过程中发挥作用。

图2 乌桕叶片盐胁迫响应蛋白KEGG代谢通路富集分析A.与SS18 未处理(0 h)相比,盐胁迫处理24 h 后SS18 叶片差异蛋白的KEGG 通路富集;B.与SS18 未处理(0 h)相比,盐胁迫处理72 h 后SS18 叶片差异蛋白的KEGG 通路富集;C.与ST21 未处理(0 h)相比,盐胁迫24 h 后ST21 叶片差异蛋白的KEGG 通路富集;D.与ST21 未处理(0 h)相比,盐胁迫处理72 h后ST21叶片差异蛋白的KEGG通路富集Fig.2 KEGG analysis of DEPs in T. sebifera leaves under salt stress A.KEGG analysis of DEPs in SS18 leaves after 24 h salt stress,compared with 0 h no salt stress;B.KEGG analysis of DEPs in SS18 leaves after 72 h salt stress,compared with 0 h no salt stress;C.KEGG analysis of DEPs in ST21 leaves after 24 h salt stress,compared with 0 h no salt stress;D.KEGG analysis of DEPs in ST21 leaves after 72 h salt stress,compared with 0 h no salt stress
对于盐耐受型乌桕无性系ST21,盐胁迫24 h时,叶片差异表达蛋白主要富集到12 个代谢通路(图2C),分别为光合有机体中的碳固定,碳代谢,乙醛酸和二元酸代谢,丙氨酸,天冬氨酸和谷氨酸代谢,光合作用,精氨酸生物合成,氨基酸生物合成,代谢途径,α-亚麻酸代谢,氮代谢,糖酵解/糖异生,缬氨酸、亮氨酸和异亮氨酸降解;当盐胁迫72 h时,叶片差异表达蛋白主要富集到4个代谢通路(图2D),分别为光合作用、内质网的蛋白质加工、光合作用—天线蛋白和碳代谢。上述结果可见ST21在盐胁迫初期(24 h),其光合作用、糖类代谢和氨基酸代谢变化较大,随着盐胁迫时间的延长(72 h),乌桕叶片的蛋白质合成能力和光合作用增加,推测其可能在盐胁迫响应过程中发挥重要作用。
2.4 代谢通路与差异蛋白网络互作分析
为进一步探索乌桕叶片对盐胁迫的响应情况,基于KEGG 代谢通路的强度和差异表达蛋白的丰度,采用Spearman 相关性分析,构建蛋白-蛋白互作网络图。与0 h相比,盐胁迫24 h下SS18叶片差异表达蛋白主要富集在3个代谢通路:丙酮酸代谢、碳代谢和糖酵解/糖异生,根据差异蛋白节点的显著相关节点数,A0A2C9W2F6、A0A2C9 WNR2、A0A2C9VR40、A0A067KBY1 和A0A2C9V8 B3 等5 个差异蛋白连接的节点数较多(图3A),基于对照组的变化倍数分别为0.33、3.11、0.32、3.76和2.58,可能是乌桕叶片响应盐胁迫的糖分解代谢核心蛋白。其中,A0A2C9WNR2和A0A2C9VR4 0蛋白分别为丙酮酸脱氢酶E1α和丙酮酸激酶,主要参与糖的有氧化、三羧酸循环和氧化磷酸化等代谢过程,在细胞线粒体呼吸链能量代谢中发挥重要作用。丙酮酸激酶是糖酵解过程中的主要限速酶之一,其表达下调提示糖分解代谢受到抑制,这可能是SS18 响应盐胁迫的重要特征之一。与0 h相比,盐胁迫72 h下SS18叶片差异表达蛋白主要富集在光合作用、光合固碳作用、碳代谢和氨基酸生物合成等代谢途径中,A0A067KCM0、A0A2C9UHW9、A0A2C9V8B3 等3 个差异蛋白的连接点较多(图3B),可能是能量代谢的关键分子,其中,A0A2C9UHW9 蛋白为磷酸甘油酸激酶,它是能量代谢调控关键酶,在植物体呼吸作用中发挥重要的调控作用。磷酸甘油酸激酶与其他相关差异蛋白均负相关,盐胁迫72 h时磷酸甘油酸激酶下调表达。

图3 乌桕叶片对盐耐受响应的差异蛋白互作网络圆形结点表示差异蛋白,结点大小表示蛋白表达的相对丰度,绿色越深表示差异蛋白变化倍数越低,红色越深表示差异蛋白变化倍数越高;方形结点表示代谢通路,蓝色越深表示-log(P)越大,黄色越深表示-log(P)越小。圆形结点之间的连线表示显著相关性,红线表示显著正相关,即激活作用;绿线表示显著负相关,即抑制作用;黑线表示显著相关;A.与SS18未处理(0 h)相比,盐胁迫处理24 h后SS18 叶片差异蛋白的互作分析;B.与SS18 未处理(0 h)相比,盐胁迫处理72 h 后SS18 叶片差异蛋白的互作分析;C.与ST21 未处理(0 h)相比,盐胁迫处理24 h后ST21叶片差异蛋白的互作分析;D.与ST21未处理(0 h)相比,盐胁迫处理72 h后ST21叶片差异蛋白的互作分析Fig.3 PPI analysis of DEPs in T. sebifera leaves under salt stress The nodes represented the different proteins,the size of node represented the relative abundance,the green nodes meant the low change fold,and the red nodes meant the high change fold,the square nodes represented the KEGG pathways,the blue nodes meant greater value of -log(P),and the yellow nodes meant smaller value of -log(P),the red lines meant the significantly positive correlations,the green lines meant the significantly negative correlations,and the black lines meant the significant correlations;A.PPI analysis of DEPs in SS18 leaves after 24 h salt stress,compared with 0 h no salt stress;B.PPI analysis of DEPs in SS18 leaves after 72 h salt stress,compared with 0 h no salt stress;C.PPI analysis of DEPs in ST21 leaves after 24 h salt stress,compared with 0 h no salt stress;D.PPI analysis of DEPs in ST21 leaves after 72 h salt stress,compared with 0 h no salt stress
与0 h 相比,盐胁迫24 h 时ST21 叶片差异表达蛋白主要富集在碳代谢、光合固碳作用、代谢途径和光合作用代谢途径中,其中鉴定出5个显著正相关核心差异蛋白(图3C),A0A067JME5、C0LE70、A0A2C9UVU5、B1NWE9 和C0LE74 分别为磷酸甘油酸激酶、光系统Ⅱ CP43 反应中心蛋白、PsaL 蛋白、光系统I P700 叶绿素a 载脂蛋白A2和光系统I P700 叶绿素a 载脂蛋白A1。乌桕ST21在胁迫前期(24 h)可能通过增强光系统I 和光系统Ⅱ相关蛋白质的合成抵御对盐胁迫的抗性。与0 h 相比,盐 胁 迫72 h 下ST21 叶 片 中 共 鉴 定 出12 个显著正相关核心差异表达蛋白(图3D),其中8 个与内质网的蛋白质加工功能相关,分别为A0A2C9VQM0(未知蛋白)、A0A2C9VY21(H+-ATP酶)、A0A2C9WD89(小分子热休克蛋白)、A0A067K5L6(H+-ATP 酶)、A0A067KV00(含TOG结构域的蛋白质)、A0A067JSA0(小分子热休克蛋白)、A0A067LGB8(小分子热休克蛋白)、A0A067K3C8(小分子热休克蛋白);4 个与叶绿素a-b 结合蛋白相关,分别为A0A2C9VK48、A0A067K1A4、A0A067JKE4、A0A2C9UIB0。乌桕ST21 可能通过促进叶绿素a-b 结合蛋白参与对盐胁迫的抗性。
3 讨论
植物耐盐性是在长期进化过程中形成的一套复杂生理生化响应逆境胁迫机制。当植物体遭到盐胁迫时,细胞膜结构和功能易遭到破坏,细胞膜透性增加,细胞内水势较高,导致植物细胞从外界吸水困难,严重时会导致体内水分大量流失,从而造成植物体生长发育严重受阻[26]。为了缓解盐胁迫造成的伤害,植物体会主动生成并积累一些可溶性有机或无机物质,如可溶性糖、脯氨酸、氨基酸和生物碱等,提高细胞内浓度,降低渗透势,增强细胞吸水能力,从而维持其正常的生长发育[27]。
本研究通过iTRAQ 蛋白定量技术分析不同盐耐受乌桕的耐盐情况,与对照组(0 h)相比,盐胁迫处理24 h 下,两种不同盐敏感度的无性系叶片中差异蛋白数目十分接近(SS18 中为53 种,ST21 中为54种),而72 h处理下的差异蛋白数目表现出较大差异(SS18 中为253 种,ST21 中为68 种)。伴随处理时间的延长,SS18 中表现出更多差异蛋白参与到盐胁迫响应过程中。盐胁迫条件下2 种无性系叶片中同时存在4 种均显著上调表达的差异蛋白,包括过氧化氢酶(A0A2C9WCZ7 和A0A2C9VX 86)、延伸因子-1α(A0A067JYP3 和A0A2C9W7J 6)、含H+-ATPase_c 结构域的蛋白(A0A2C9VY21)和硫氧还蛋白(A0A2C9UAC6 和A0A067KJ27),已有研究发现这些蛋白在植物耐盐应激响应过程中发挥重要作用,推测其可能是乌桕耐盐响应的重要潜在靶标蛋白。尚晓蕊等[28]研究发现,盐胁迫下拟南芥角果中也存在3 种(Q42578、Q9LHB9、F4IQ05)过氧化物酶蛋白发生差异表达,其在拟南芥耐盐胁迫发挥重要作用。王强等[29]研究番茄(Lycopersicon esculentum)幼苗盐胁迫下响应差异表达蛋白,筛选出11种重要的潜在靶标蛋白质,其中包括过氧化物酶和延长因子1α等重要蛋白。宁夏枸杞(Lycium barbarum)液泡膜H+-ATPase 基因LbHA1表达量与植株体内Na+含量存在一定的相关性,在胁迫初期,液泡膜H+-ATPase 参与了根细胞中Na+及时排出胞外和区隔于液泡,从而维持根细胞内Na+的稳定性[30]。此外,花生(Arachis hypogaea)在受到盐胁迫后,为Na+反向转运和区域化的Na+/H+逆转运蛋白提供跨膜驱动力的质膜H+-ATPase 基因AhHA1的表达水平发生显著变化,研究证实花生AhHA1基因能够促进转基因拟南芥植株根的生长发育和提高耐盐能力[31]。比拉底白刺(Nitraria billardieri)幼苗叶片应答盐胁迫研究中发现硫氧还蛋白F1 在蛋白互作网络中发生相互作用进而参与盐胁迫应答响应过程[32]。本研究获得的4 种潜在靶标蛋白与上述研究获得的耐盐响应过程中的重要差异蛋白一致。在对上述4 种显著上调表达的重要潜在靶标蛋白进行KEGG 富集分析发现,过氧化氢酶(A0A2C9WCZ7 和A0A2C9VX86)富集到过氧酶体通路;延伸因子-1α(A0A067JYP3)富 集 到RNA 转 运 通 路;含H+-ATPase_c 结构域的蛋白(A0A2C9VY21)富集到植物—病原菌相互作用通路;硫氧还蛋白(A0A2C9 UAC6)富集到核糖体通路。
碱地肤(Kochia sieversiana)受到盐碱胁迫时,植物体会通过积累大量的有机酸(如草酸、苹果酸等),维持细胞内离子平衡及调节pH,有效缓解盐碱胁迫造成的伤害[33]。乌桕叶片可能通过提升光合作用,加强糖类代谢和氨基酸代谢,增加细胞内小分子有机物(可溶性糖、氨基酸等)含量,减轻盐胁迫造成的伤害。通过对比2 种盐耐受型乌桕在不同盐胁迫时间的差异蛋白代谢通路,发现乌桕ST21 的叶片在盐胁迫初期(24 h),会增强多个糖类代谢和氨基酸代谢途径参与到盐胁迫响应过程中。本研究中差异蛋白KEGG 富集分析显示,SS18 在盐胁迫24 h 时和ST21 在盐胁迫72 h 时,α-亚麻酸代谢、乙醛酸和二元酸代谢通路中的差异蛋白均显著富集,可见有机酸的变化对乌桕盐胁迫响应过程可能具有重要意义。
大量的研究表明,植物响应盐胁迫与能量代谢密切相关,参与能量代谢蛋白主要涉及糖酵解、三羧酸循环、磷酸戊糖途径和ATP 合成等相关调控酶[34-36]。番茄幼苗受到盐碱胁迫后,糖酵解途径关键酶甘油醛-3-磷酸脱氢酶表达量增加,表明植物通过增强呼吸代谢,释放大量的能量,来维持自身生长发育及抵御外界盐胁迫[37]。张恒等[38]研究发现三倍体小黑杨(Populus simonii×P.nigra)杂种无性系受到盐胁迫后,苹果酸脱氢酶呈下调表达,影响TCA 循环的正常运转,进而影响植株的生长。Kav 等[39]研究豌豆(Pisum sativum)苗受到盐胁迫处理时,发现根中三羧酸循环途径能量代谢关键酶,如α-酮戊二酸脱羧酶、丙酮酸脱氢酶等,表达均显著上调,表明充足的能源对植物能够适应盐环境发挥重要的作用。本研究中,当SS18 受到盐胁迫24 h 后,糖酵解和丙酮酸代谢途径增强,其关键调控酶(丙酮酸激酶和丙酮酸)表达上调;盐胁迫72 h 后,糖酵解关键调控酶磷酸甘油激酶表达下调,这些变化可能在其抵御外界盐胁迫中发挥一定作用。ST21 在盐胁迫72 h 后,磷酸甘油激酶表达上调,推测其可能与ST21 植株遭受长时间盐胁迫下的抵御盐胁迫能力有关。
植物遭受盐胁迫不仅仅表现在能量代谢受阻,还会严重影响其光合作用。相关的研究表明盐胁迫对植物光合作用的抑制是多方面的,不仅包括CO2的吸收、运输和固定,还包括对光能的吸收、转运和电子传递等的影响[34]。赵海燕等[40]研究表明,在盐胁迫条件下,银杏(Ginkgo biloba)幼树叶片内叶绿素合成受阻。张若溪等[41]研究发现,栾树(Koelreuteria paniculata)受较高浓度混合盐胁迫时,非气孔限制和光化学活性失活影响其光合作用,是导致植物生长受阻的主要原因。张潭等[42]发现高浓度的盐碱胁迫破坏枸杞幼苗的叶片光合结构,降低光环境适应能力和光合作用效率,是影响枸杞幼苗正常生长的关键机制。赵娟娟等[43]研究发现NaCl 胁迫损伤酸枣(Ziziphus acidojujuba)幼苗PSⅡ反应中心,导致酸枣幼苗光合作用下降。李俊良[44]利用iTRAQ 蛋白定量技术,在甜菜(Beta vulgaris)叶片中鉴定出70 个差异表达蛋白,证明甜菜通过上调光系统Ⅱ(PSⅡ)的PsbQ 蛋白、质体蓝素和硫氧还原蛋白保障光合作用,是其对盐胁迫应答的主要分子机制之一。本研究结果显示,在受到盐胁迫时,乌桕叶片差异表达蛋白在光合作用和光合固碳作用代谢通路中显著富集,表明光合作用是受到盐胁迫影响的重要生物过程之一;通过构建PPI 网络图分析,SS18 植株在遭受盐胁迫后,光合作用代谢发生变化。而ST21植株在盐胁迫24 h后,光系统Ⅱ CP43反应中心蛋白、PsaL 蛋白,光系统I P700 叶绿素a 载脂蛋白A2 和光系统Ⅰ P700 叶绿素a 载脂蛋白A1 等光合作用相关蛋白表达显著上调;在盐胁迫72 h 时,4个与叶绿素a-b结合蛋白表达显著上调。可见盐耐受型乌桕可能通过增强光合系统(光系统Ⅰ和光系统Ⅱ)进而提高自身耐盐能力。
综上所述,乌桕叶片响应盐胁迫涉及多条代谢通路,并且参与各个代谢过程的相关蛋白并不是呈现单一的变化趋势,说明其响应盐胁迫机制十分复杂。与SS18相比,ST21在遭受盐胁迫时,会启动多个糖类代谢、氨基酸代谢和脂肪酸代谢途径,在细胞内生成并积累大量可溶性糖、氨基酸和有机酸等小分子可溶性物质,这些变化可能是其抵御外界盐胁迫对植物体造成伤害的主要方面。本研究初步揭示了乌桕响应盐胁迫的蛋白变化,为进行耐盐乌桕品种选育及研究提供理论依据。
