猪TRIM32分子的克隆、表达及其功能分析
2023-09-08张彦兵解艺璇魏建超李宗杰邵东华李蓓蓓马志永邱亚峰
李 慧,张彦兵,解艺璇,陆 艳,相 笑,刘 珂,魏建超,李宗杰,邵东华,李蓓蓓,马志永,邱亚峰
(中国农业科学院上海兽医研究所,上海 200241)
TRIM32(The tripartite motif 32)作为TRIM家族中的成员之一,与TRIM家族其他成员的结构相似,其N端具有典型的RBBC结构,即一个RING结构域、两个B-BOX结构域、一个Cioned-coin卷曲螺旋结构域,与此相连的为1个间隔片段和6个NHL重复序列[1-2]。作为一种E3泛素连接酶,TRIM分子可催化泛素(Ub)分子从E2传递到靶蛋白,从而使靶蛋白发生泛素化修饰。在此过程中,其N端的RING结构域是必须的。已有研究表明,TRIM32依赖于RING结构域能够靶向多种蛋白并通过蛋白酶体途径使之泛素化降解[3-6]。基于这种广泛的底物特异性,TRIM32参与多种生命活动包括发育和分化[4,7-10]、Micro RNA的调节[4,11]、肿瘤发生过程[12-13]。
此外,现有的结果显示TRIM32在先天性抗病毒过程中也发挥着重要的作用:1)TRIM32调控IFN-β的产生发挥先天性抗病毒的作用,具体的机制为TRIM32通过介导STING的泛素化,从而显著增强STING介导的IFN-β产生[6];2)此外,TRIM32在甲型流感病毒(Influenza A virus,IAV)感染过程中,能够直接增加PB1的泛素化,通过降解PB1抑制IAV的感染[14]。
我们感兴趣于猪TRIM32分子在先天性抗病毒中的作用,然而目前,对于猪源TRIM32分子的研究还未见报道。本研究从猪肺脏中克隆了猪TRIM32基因,利用过表达和RNA干扰的方法探讨了TRIM32分子在PRRSV感染中的作用。这些研究结果为揭示猪TRIM32在先天性抗病毒中的分子机制奠定基础。
1 材料与方法
1.1 主要材料 293T细胞、Marc145细胞、HP-PRRSV毒株SY0608及真核表达载体p3xFlag-CMV-14都由本实验室保存。LATaq®高保真酶,HindⅢ/BamHⅠ限制性内切酶,PrimeScriptTMRT Master Mix(Perfect Real Time)、TB GreenTMPremix ExTaqTMⅡ购自TaKaRa 生物技术有限公司;DMEM、opti-MEM、胎牛血清、胰酶、PBS、Lipofectamine®RNAiMAX Reagent、Lipofectamine 3000、蛋白Marker均购自Thermo公司。
1.2 基因的扩增 根据GenBank上预测的猪TRIM32基因的序列设计引物,利用猪肺组织提取总RNA并反转录成cDNA为模板,利用LATaq高保真酶进行PCR扩增,引物序列见表1。PCR反应程序为:95℃预变性5 min;95℃变性30 s,68℃复性30 s,72℃延伸2 min;共35个循环;72℃再延伸7 min;4℃保存。将胶回收目的片段连接至pMD-18T载体,并送公司进行测序分析,选用测序正确的菌液提取质粒,命名为pUC-porTRIM32。

表1 PCR引物序列Table 1 PCR primers sequence
1.3 基因的真核载体构建 根据真核表达载p3xFlag-CMV-14以及猪TRIM32中限制性内切酶位点的特征,设计引物并在引物两端添加合适的保护性碱基和限制性内切酶序列(表2),以pUC-porTRIM32质粒作为模板,进行PCR扩增和琼脂糖凝胶分析。将PCR胶回收产物HindⅢ和BamHⅠ双酶切。胶回收酶切产物后,利用T4 DNA 连接酶将酶切回收的片段与同样酶切的载体进行连接,16℃过夜后,将连接产物转化到DH5α大肠杆菌中;将转化的大肠杆菌涂布含有Amp的LB琼脂平板,37℃过夜培养后,分别挑取单菌落于含有Amp的LB中,在恒温培养箱里摇菌过夜后,通过菌液PCR以及质粒酶切的方法,进行阳性质粒的鉴定。最后,对鉴定阳性的克隆,进行测序分析。
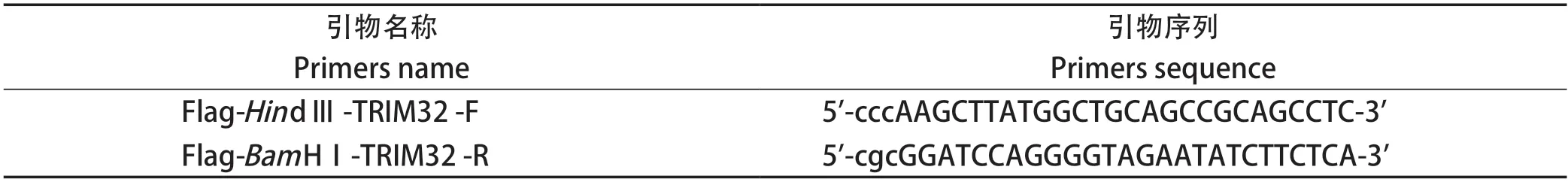
表2 PCR引物序列Table 2 PCR primers sequence
1.4 瞬时转染 将状态良好的293T细胞或Marc145细胞铺于细胞板中,待细胞长至80%密度时,利用Lipofectamine®3000试剂,将重组质粒pFlagporTRIM32或p3xFlag-CMV-14空载体转染到细胞中,具体步骤依据Lipofectamine 3000说明书进行。
1.5 RNA干扰与荧光定量PCR分析 根据猴TRIM32基因的序列,由吉玛公司设计并合成干扰RNA,利用Lipofectamine®RNAiMAX将特异的siRNA和NC转染Marc145细胞,具体根据脂质体的说明书进行。于转染后48 h后,感染HP-PRRSV(0.5 MOI),约18 h后收取细胞上清液用于TCID50分析;同时,提取细胞总RNA用于荧光定量RT-PCR分析[15]。定量PCR的引物详见表3。

表3 荧光定量PCR引物Table 3 Primers for real-time PCR
2 结果
2.1 猪TRIM32基因的克隆及真核表达载体的构建 为了克隆猪TRIM32基因,利用RT-PCR从猪肺组织中扩增获得约2000 bp的基因片段(图1A);将PCR胶回收产物克隆至pMD18-T载体中,对阳性的克隆进行测序并对获得的序列进行分析,结果显示扩增获得的片段为1956 bp,与GenBank预测的序列相同。随后,通过酶切、连接将目的片段克隆入p3xFlag-CMV-14的HindⅢ和BamHⅠ位点,阳性质粒利用HindⅢ和BamHⅠ双酶切,可以切出两个片段,分别为6300 bp和2000 bp(图1B),阳性质粒命名为pFlag-porTRIM32。
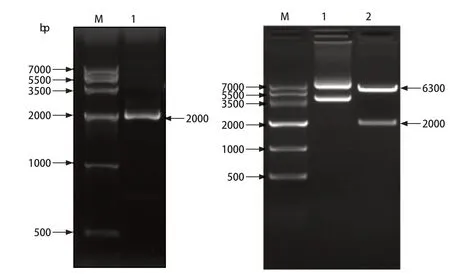
图1 猪TRIM32的基因克隆及真核表达载体的构建Fig.1 The segmental amplification of porcine TRIM32 gene and construction of eukaryotic expression vector of porcine TRIM32
2.2 重组质粒pFlag-porTRIM32的表达分析 为了验证重组质粒pFlag-porTRIM32的表达,通过脂质体将重组质粒和空载体瞬时转染入293T细胞。于转染后24 h,收获细胞并制备蛋白样品。利用Western blot对重组质粒的表达进行分析,结果显示,与转染空载体的样品相比,在转染重组质粒的细胞中,抗Flag的抗体检测到约72 kDa的特异条带(图2)。
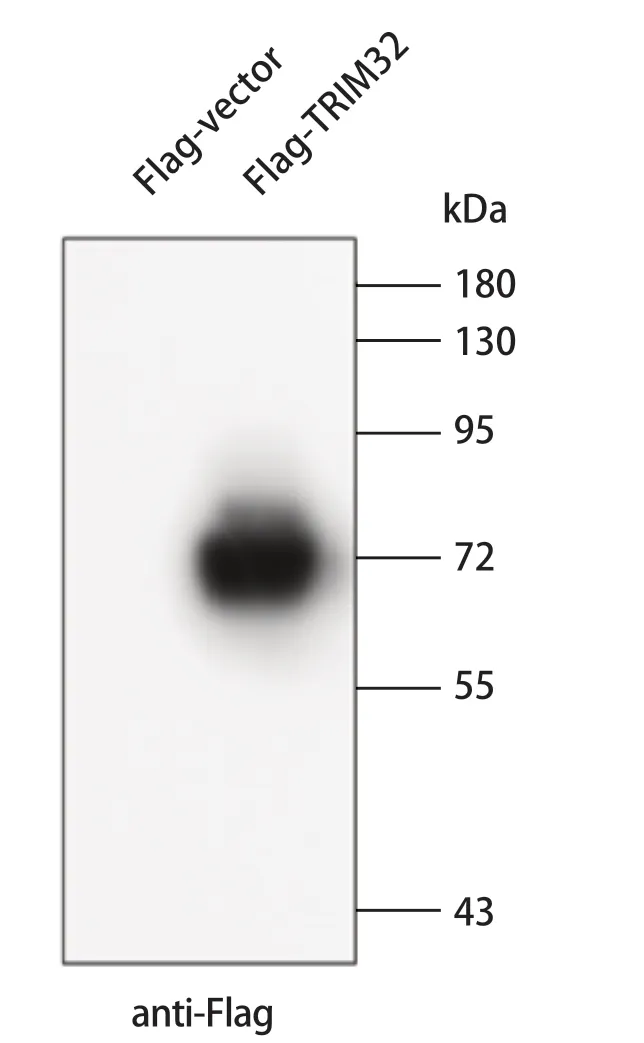
图2 Western blot鉴定重组质粒pFlag-porTRIM32的表达Fig.2 Analysis of expression of recombinant plasmid pFlag-porTRIM32 by Western blot
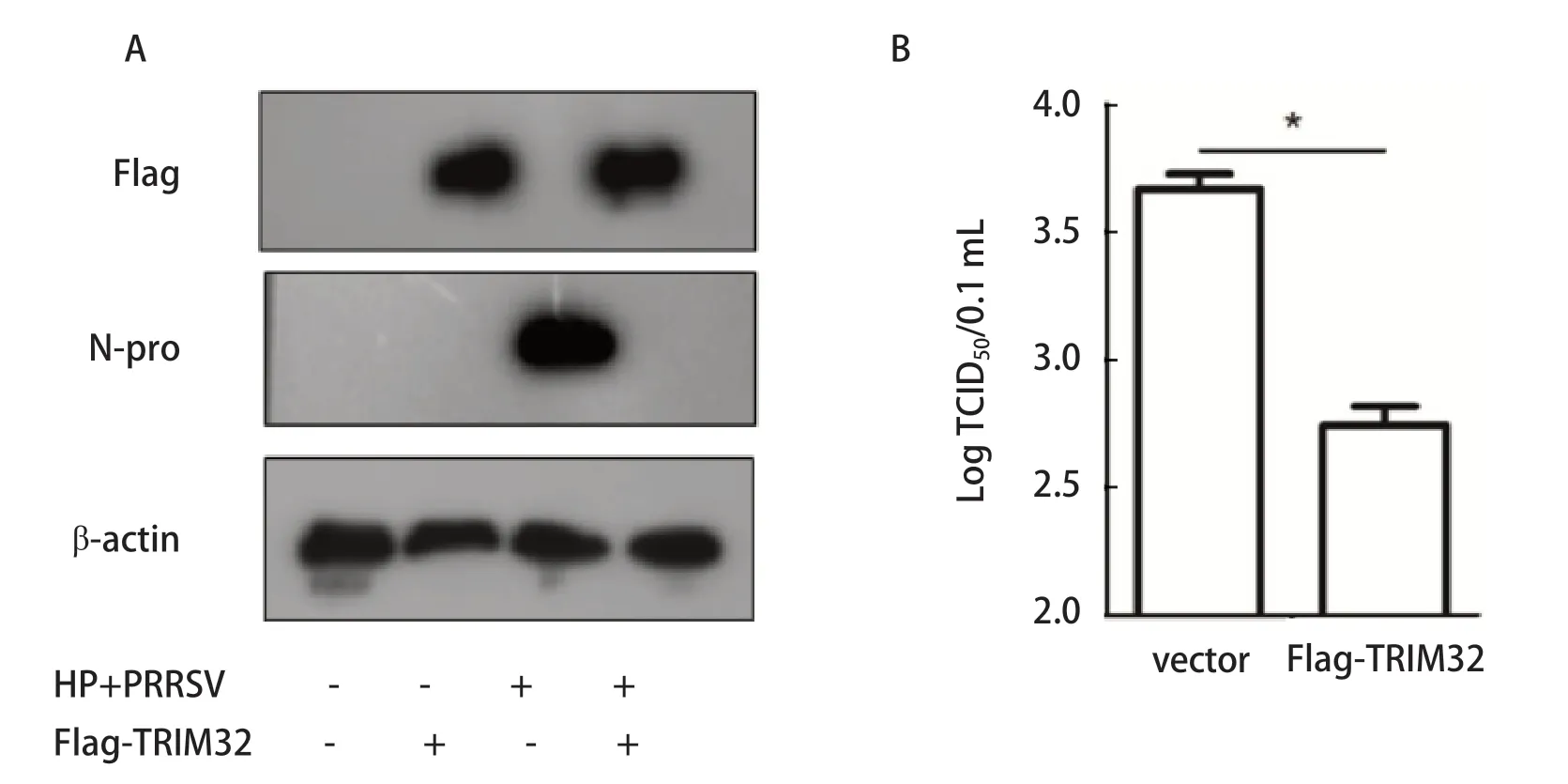
图3 过表达猪TRIM32对HP-PRRSV感染的影响分析Fig.3 Effect of overexpression of porcine Flag-TRIM32 on HP-PRRSV infected cells
2.3 过表达猪TRIM32分子对HP-PRRSV感染的影响 实验室前期数据显示PRRSV感染抑制TRIM32的表达(mRNA水平,结果未显示),然而,TRIM32在PRRSV感染中的作用不清楚。鉴于TRIM32介导先天性抗病毒的作用,本研究利用PRRSV为模型,探究猪TRIM32在HP-PRRSV感染中的作用。我们利用过表达猪TRIM32分子验证其在PRRSV感染中的作用。Western blot的结果显示,Flag-porTRIM32在Marc145细胞成功过表达,而且与Flag-vector 组相比,过表Flag-porTRIM32组,病毒结构蛋白N蛋白的表达量明显受到抑制(图4A);进一步,利用TCID50对上清液中的病毒量进行了分析,结果同Western blot相符,与vector组相比,过表达猪Flag-TRIM32组能有效抑制HP-PRRSV的感染(图4B)。
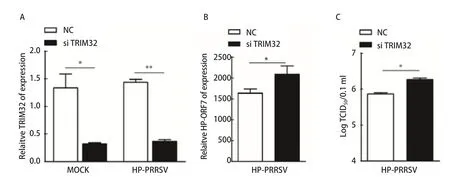
图4 沉默TRIM32对HP-PRRSV感染的影响分析Fig.4 Effect of silencing of TRIM32 on HP-PRRSV infected cells
2.4 沉默TRIM32的表达对HP-PRRSV感染的影响 为了进一步探究TRIM32在HP-PRRSV感染中的作用,我们利用Lipofectamine®RNAiMAX将猴TRIM32特异的siRNA转染到Marc145细胞中,通过qPCR的检测结果显示:RNA干扰能够有效限制TRIM32的表达(图4A);同时,相对于NC组,RNA干扰组的PRRSVORF7基因表达有明显上升趋势(图4B)。进一步,通过对细胞上清液进行TCID50分析,结果显示沉默内源TRIM32的表达显著促进病毒的感染(图4C),这一结果与qPCR的结果一致。
3 讨论
鉴于猪TRIM32的分子还没有克隆,本研究首先克隆了猪TRIM32分子,并将其克隆入真核表达载体中进行表达分析。结果显示猪TRIM32的分子量约为72 kDa,与预期的大小一致。尽管可以利用商业化的抗体(针对人TRIM32分子的抗体)进行猪TRIM32分子的表达鉴定,但目前还没有筛选到合适的抗体应用猪TRIM32的表达检测。下一步我们将根据克隆的基因序列,进行针对猪TRIM32分子特异抗体的制备。
尽管我们在前期的研究中揭示了PRRSV感染影响TRIM32在猪肺泡巨噬细胞上的表达(结果未显示),然而,对TRIM32在PRRSV感染中的作用不清楚。我们利用过表达猪TRIM32分子揭示了TRIM32具有抑制PRRSV感染的作用。进一步利用RNA干扰方法,沉默Marc145细胞中内源性的TRIM32,可以显著地增加PRRSV的感染这一研究结果揭示了TRIM32具有抗PRRSV感染的作用。已有的结果显示TRIM32分子对多种病毒的感染具有抑制作用,包括水泡性口炎病毒(VSV)、新城疫病毒(NDV)、流感病毒(AIV)等。其中,TRIM32介导的抗病毒的机制包括:1)TRIM32通过STING的泛素化调控Ⅰ型干扰素的产生,继而发挥抗病毒的作用;2)TRIM32直接靶向于病毒蛋白,介导病毒蛋白的降解。目前这个阶段,猪TRIM32是如何发挥抗体PRRSV感染的作用机制,还不清楚。我们接下来的工作,将聚集于TRIM32的这些作用进行相关机制的解析。
总之,本研究成功克隆了猪TRIM32基因,并鉴定了猪TRIM32的表达,揭示了TRIM32具有抗PRRSV感染的作用。下一步,我们将聚焦于猪肺泡巨噬细胞(PAM)上TRIM32抗PRRSV感染的机制的研究。本研究结果不仅为揭示猪TRIM32抗PRRSV感染的机制奠定基础,而且为进一步理解猪TRIM32的生物学功能奠定基础。
