玉米开花期相关性状的QTL定位与候选基因分析
2023-08-31刘鹏飞郭向阳王安贵祝云芳陈泽辉
郭 爽, 王 栋, 聂 蕾, 何 玥, 涂 亮, 刘鹏飞,郭向阳, 王安贵, 祝云芳, 吴 迅,3, 陈泽辉
(1.贵州大学农学院, 贵阳 550025; 2.贵州省农业科学院旱粮研究所, 贵阳 550006;3.农业农村部喀斯特山区作物基因资源与种质创新重点实验室, 贵阳 550006)
玉米作为重要的粮食、饲料和工业原料作物,在社会经济发展中发挥着重要作用。开花期相关性状是植物响应环境变化的关键性状,在植物繁衍和适应生态环境等方面都具有非常重要的作用[1],不同玉米品种对光照长度的响应存在差异,表现出在长短日照条件下的开花期差异,如大部分热带玉米种质在热带地区能正常开花,而到了温带地区则出现徒长不能正常开花结实的现象,也是热带玉米引种温带种植迫切需要解决的关键问题。植物花发育的过程一般要历经开花诱导、花原基形成、花器官发育这3个阶段[2]。玉米开花诱导调控网络主要分(光周期途径、自主途径、赤霉素途径和年龄途径)4个途径[1],其作为典型的数量性状,遗传方式复杂且受环境影响严重。为了探究玉米开花期性状的遗传规律,多数研究者围绕开花期相关性状进行QTL定位和基因挖掘,以提高育种选择效率。侯清桂等[3]以豫86×豫M1-7构建的RIL群体为材料,对玉米开花期相关性状进行QTL分析,共鉴定到48个开花期性状相关的QTLs,包括18个抽雄期QTLs、16个吐丝期QTLs和14个散粉期QTLs。这些QTLs在除第4号和第8号染色体外的其他染色体上都有分布,其中单个QTL表型贡献率在1.67%~20.33%之间。王迪等[4]以自交系齐319和掖478分别与黄早四杂交构建的230个和235个F2∶3家系为材料,利用完备区间作图方法,对不同生态环境玉米花期相关性状进行QTL定位,其中在玉米Q/H群体的10个连锁群上共定位到了85个QTLs,有30个QTLs在Y/H群体被检测到,并呈现成簇分布。因此,较多研究者利用经典遗传学方法,通过数量性状基因组(QTL)初步定位、精细定位,最终实现目标基因的图位克隆。众多学者也鉴定出一批与玉米开花相关的重要基因,如ZmCCT[5],Vgt1[6]、ZCN8[7]、Dwarf[8]和ZmCCT9[9]等。高媛等[10]在对Suwan种质代表性自交系T32和QR273的开花相关基因差异表达模式的研究中发现,长日照条件开花促进基因ZmCOL、ZmELF4、ZmGI、ZmHd1、ZmHd6、ZCN8在光钝感系QR273中的相对表达量显著高于光敏感系T32;开花抑制基因ZmCCT9、ZmCCT10的相对表达量则显著低于T32。这些研究为深度解析玉米开花相关性状变异的遗传机制以及育种改良利用提供了依据。综上所述,针对开花期相关性状定位到的QTL在数量、位置、基因效应上因所用材料、标记等不同而存在差异,玉米开花期性状遗传基础和基因调控网络较为复杂。因此,本研究通过选用热带种质选育骨干亲本QR273和T32为亲本构建的150份F2和F2∶3分离家系为材料,基于简化基因组测序(GBS)技术对F2单株进行基因型鉴定,同时对该群体的F2∶3家系进行多个环境开花期相关性状评价,利用完备区间作图法对开花期相关性状进行QTL定位。在此基础上,利用生物信息分析方法筛选玉米开花期相关的候选基因,为基于分子辅助的玉米开花期性状改良提供理论支撑。
1 材料与方法
1.1 试验材料
本研究以热带种质选育玉米骨干亲本QR273和T32为亲本,构建包含150个家系的F2、F2∶3家系为材料。T32是贵州省农业科学院旱粮研究所从Suwan种质中选育而成的骨干自交系,表现为光周期敏感特点,即在长日照地区会出现开花期延迟甚至不能正常开花结实等现象;QR273是以Suwan群体作基础材料,经过5年8代自交选育而成的优良玉米骨干自交系,表现为光周期钝感的特点,即在长、短日照条件下均表现为正常开花结实的特征。
1.2 田间试验及测定方法
2021年10月,将构建的F2分离群体与亲本种植于海南省三亚市,对幼苗5叶期的单株进行取样,并采用改良CTAB法对DNA进行提取,随后DNA质量检测和GBS测序委托北京康普生生物有限公司完成,具体过程参照吴迅等[7]的方法。同时对F2植株进行单株自交后可获得F2∶3家系种子。
2022年4月以及2022年10月,将F2∶3群体以及亲本材料分别种植于贵州省贵阳市、甘肃省张掖市和海南省三亚市,材料的种植采用随机区组设计,2次重复,行长3 m,行距0.65 m。田间管理的方式与当地一致。田间调查玉米抽雄期、散粉期、吐丝期,性状评价参考石云素等[11]的方法。
1.3 数据分析
通过Excel软件对表型数据进行整理,SPSS26.0软件进行统计分析。分别计算3个环境下亲本与不同家系的抽雄期、散粉期、吐丝期的平均值、变异系数、偏度、峰度、方差等,并进行相关性分析。
1.4 QTL定位与候选基因预测
对获得的SNP标记进行筛选,删除等位基因频率MAF<0.05的SNP位点后,共获得68 994个高质量SNP标记可用于后续分析。利用IciMapping4.1软件中的完备区间作图法(ICIM)对3个开花期相关性状进行QTL检测。其中,作图步长为1.00 cM,PIN为0.001,LOD阈值为2.50。根据3种环境中的定位结果和已报道的QTL相关信息,以是否存在重叠区域为标准对“一致性”QTL进行判断,若存在重叠区域,则认为是“一致性”QTL[12]。在此基础上,结合B73参考基因组序列(RefGen_v4),查找在“一致性”QTL区间内控制目标性状的候选基因,并结合基因功能注释和已有研究报道,对关键候选基因进行筛选。
2 结果与分析
2.1 开花期相关性状数据分析
对开花期相关性状的统计分析结果显示,在张掖市和贵阳市,开花期相关性状在基因型间表现出显著差异。亲本T32的抽雄期、散粉期、吐丝期的平均值均高于亲本QR273。且这3个开花期相关性状在不同家系间差异显著。其开花期相关性状的偏度和峰值的绝对值均小于1,符合正态分布,可作QTL定位分析(表1)。

表1 开花期相关性状的描述性统计Table 1 The descriptive statistical of related traits at flowering stage
2.2 开花期性状相关性分析
开花期相关性状间的相关性分析结果列于表2。结果显示,抽雄期与散粉期、吐丝期,散粉期与吐丝期在张掖市、贵阳市、三亚市三地均呈极显著正相关,说明3个开花期性状通过相互协同作用促进玉米生长发育的调控,这与刘颖等[13]研究结果一致。
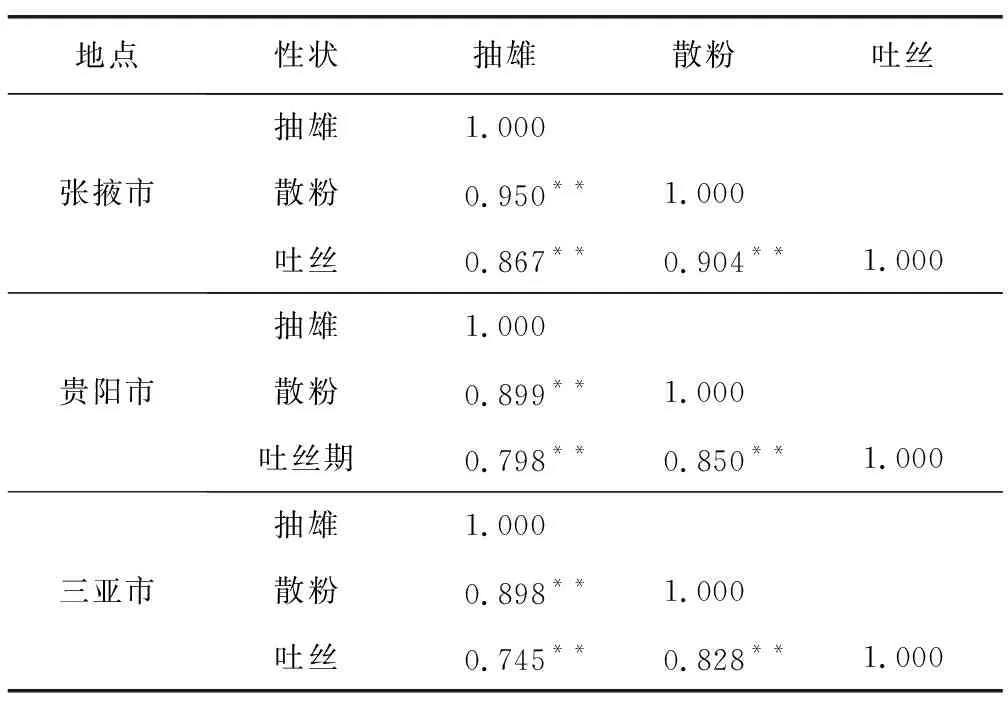
表2 开花期性状的相关性分析Table 2 Correlation analysis between related traits at flowering stage under three environments
2.3 SNP图谱构建
依据获得的高质量SNP标记的物理图谱,68 994个SNPs均匀地分布在染色体上(图1)。各染色体上的标记数量大小依次表现为Ch4、Ch2、Ch1、Ch3、Ch5、Ch8、Ch9、Ch7、Ch6、Ch10,其数量分别为8 901个、8 824个、8 725个、7 623个、7 174个、6 407个、5 782个、5 592个、5 432个、4 534个。

注:每一行代表一个SNP标记,浅色表示标记的密度较低,深色表示标记的密度较高。图1 遗传图谱Fig.1 Genetic map
2.4 控制开花期相关性状的QTL
3个开花期相关性状的QTL定位结果列于表3。结果显示,3个环境下共检测到98个开花期相关QTL,其中37个QTL与抽雄期相关,分别位于1号、2号、5号、6号、7号、9号、10号染色体上,单个QTL的贡献率在0.85%~12.43%之间,且12个QTL的增效基因来源于亲本T32,23个QTL的增效等位基因来源于亲本QR273;与散粉期有关的QTL27个,各个染色体上均有分布。单个QTL的贡献率在3.29%~8.64%之间,且10个QTL的增效基因来源于亲本T32,17个QTL的增效等位基因来源于亲本QR273。

表3 玉米开花期相关性状QTL定位结果Table 3 QTL mapping results of maize related traits at flowering stage
与吐丝期有关的QTL 30个,各个染色体上均有分布。单个QTL的贡献率在1.78%~10.24%之间,其中有6个QTL的增效基因来源于亲本T32,24个QTL的增效等位基因来源于亲本QR273。共检测到8个贡献率大于8%的QTL,其中3个与抽雄期相关,2个与散粉期相关,3个与吐丝期相关,分别位于第4号染色体上的142.2~142.4 Mb之间,第2号染色体上的5.3~5.8 Mb之间、第5号染色体上的115.2~115.4 Mb之间、第9号染色体上的76.4~76.6 Mb之间、第7号染色体上的55.0~55.1 Mb、第9号染色体上的76.4~76.6 Mb之间、第4号染色体上的133.1~133.2 Mb之间、第9号染色体上的88.0~89.1 Mb之间。
2.5 一致性QTL
通过比较不同环境下的QTL定位结果发现,在第1,4,6,9号染色体上分别存在QTL富集区域即同一标记区间影响多个目标性状。如在第1号染色体的68.0~68.3 Mb区间检测到一个控制抽雄期和散粉期的QTL,在第4号染色体的63.2~63.4 Mb区间内检测到一个同时控制抽雄期和散粉期的QTL,在第4号染色体的133.1~134.1 Mb区间检测到一个同时控制散粉期和吐丝期的QTL,在第6号染色体的129.3~129.5 Mb区间内检测到一个同时控制抽雄期和散粉期的QTL。在第9号染色体的76.5~76.6 Mb区间内检测到一个同时控制抽雄期和散粉期的QTL。与已公开QTL数据库的比对发现,本研究有8个检测到的QTL与前人定位结果一致,分别控制抽雄期、散粉期和吐丝期的变异,可作为下一步精细定位的候选靶标。
2.6 候选基因分析
结合Maize GDB数据库的基因功能注释信息以及前人的研究结果,对筛选出的8个一致性QTL进行目标候选基因分析,初步预测出12个候选基因与开花期相关(表4)。侯选基因中,Zm00001d052638、Zm00001d025735、Zm00001d025739、Zm00001d022350、Zm00001d005135参与编码蛋白质丝氨酸/苏氨酸激酶活性。沈辰[14]通过实时定量PCR分析发现,丝氨酸/苏氨酸蛋白磷酸酶基因在山核桃的多个器官中均有表达,其中在花芽中表达量最高,且这些表达存在于山核桃雌花和雄花的整个发育过程。Zm00001d052585编码苯丙素生物合成过程。武绍龙等[15]研究发现,马缨杜鹃花开花至代谢进程中,苯丙素类代谢物表达量逐渐升高,表明苯丙素类代谢物表达可能与马缨杜鹃花发育有关,Zm00001d052637参与对激素的反应。激素是调控植物生长发育重要的物质之一,在植物的开花方面发挥着重要的调控作用。Zm00001d047632参与红光、远红光光传导,曹凯等[16]研究发现,红光与远红光比值低的环境会抑制野生型番茄和phyB1突变体番茄开花。Zm00001d042717与乙烯激活的信号通路相关,张华等[17]研究表明,75 mg/L乙烯利处理对植株花枝数、花朵数、始花期及开花总时长均有明显的促进作用。任小林等[18]研究发现,秋施乙烯利可使牡丹的落叶期提前,萌芽期和开花期延迟,且显著降低了开花率,减小了花径,并且增加了畸形花的百分率。Zm00001d044325、Zm00001d003383均参与生长素激活的信号通路。也有研究表明,生长素对植物的开花具有影响。杨洪全等[19]研究发现,光激活的CRY1和phyB能够通过直接结合和稳定AUX/IAA来抑制生长素信号通路,说明光和生长素可以拮抗调节AUX/IAA的稳定性,从而平衡生长素和光对植物生长发育的影响。生长素能够通过与细胞分裂素、茉莉素以及脱落酸之间的相互作用来共同影响植物的生长发育[20]。Zm00001d003379、Zm00001d042721参与脱落酸激活的信号通路,邓全恩[21]用赤霉素对油茶花进行处理时发现,当赤霉素浓度为400 mg/L时,油茶花始花期最早且盛花期集中,150 mg/L脱落酸处理时,始花期最晚;高英[22]对核桃花芽分化过程中脱落酸进行原位分析发现,由叶芽状态向雌花芽转化时,顶端分生组织中脱落酸的水平明显降低,对生长素和脱落酸的进行平衡分析后发现雌花芽的诱导需要较高水平的脱落酸/生长素。

表4 基因位置及功能注释Table 4 The gene location and function annotation
3 结果与讨论
本研究共定位到8个控制开花期相关性状的一致性QTL,包括郭瑞[23]利用光周期敏感自交系CML288和光周期钝感自交系黄早四为材料构建F2∶3作图群体,在第2号染色体和第9号染色体上分别检测到1个与吐丝期相关的QTL和1个与抽雄期相关的QTL,对应区间为26.1~120.3 Mb和127.0~139.5 Mb(B73RefGen_v4),本研究也在第2号染色体42.0~42.6 Mb和第9号染色体137.4~137.7 Mb分别定位到1个吐丝期QTL和1个散粉期QTL;郭瑞还利用光周期敏感自交系CML288和光周期钝感自交系黄早四为材料构建F2∶3作图群体,在第2号染色体和10号染色体上分别检测到2个与抽雄期相关的QTL,对应区间为26.1~208.9 Mb和115.7~140.3 Mb(B73RefGen_v4);本研究也在第2号染色体160.4~160.5 Mb和第10号染色体127.7~128.2 Mb内分别定位到1个散粉期QTL;魏海忠[24]以玉米自交系80007和80044衍生的包含355个家系的F9重组自交系(RIL)群为材料,采用SSR标记在第3号染色体检测到1个与散粉期相关QTL,对应区间为224.5~225.3 Mb(B73 RefGen_v4);本研究在3号染色体225.2~225.3 Mb定位到1个散粉期QTL;李永明等[25]利用4个玉米自交系组配276/72//A188/交51四交群体,构建了包含213个SSR分子标记的遗传连锁图谱在第3号染色体上和第7号染色体上分别检测到1个与散粉期相关的QTL和1个与吐丝期有关的QTL,对应区间为171.0~194.8 Mb和170.3~177.3 Mb(B73RefGen_v4);本研究在3号染色体178.1~178.4 Mb和第7号染色体175.8~176.1 Mb上也分别定位到1个散粉期QTL和1个吐丝期QTL。李志敏[26]在第4号染色体上检测到1个与抽雄期相关的QTL,对应区间为189.1~216.4 Mb(B73 RefGen_v4);本研究在第4号染色体195.2~195.5 Mb内定位到了1个抽雄期QTL。由此推测这些一致性位点在玉米驯化过程中表达较稳定,可作为进一步精细定位并挖掘关键候选基因的候选靶标。
除此之外,本研究还检测出12个开花期相关的候选基因,主要参与激素合成、运输及信号转导等,所编码蛋白在作物生长发育过程中参与酶类调控和激素调节。与在山核桃、番茄等植物中的研究基本一致。研究结果能够为后续的精细定位与候选基因的图位克隆提供新的靶标。
