江浙地区菱品种遗传多样性的SLAF-seq分析
2023-08-29王凌云叶雪莲权新华王瑞森姚祥坦
袁 晔,刘 睿,王凌云,沈 盟,叶雪莲,权新华,王瑞森,姚祥坦,*
(1.嘉兴市农业科学研究院,浙江 嘉兴 314016; 2.浙江大学 蔬菜研究所,浙江 杭州 310058; 3.金华市农业科学研究院(浙江省农业机械研究院) 浙江省特色水生蔬菜育种与栽培重点实验室,浙江 金华 321000)
菱属(TrapaL.)为菱科(Trapaceae)一年生浮叶水生草本植物,生长于浅水湖泊、河湾、池塘和水田中,在我国分布广泛,尤其是在黑龙江流域和长江中下游地区种类十分丰富[1-2]。菱属植物是重要的水生蔬菜种类,其成熟果实淀粉含量高达80%,被联合国粮农组织(FAO)列为重要的水生粮食作物,在中国和印度多年来被广泛种植[3-4]。栽培菱品种具有产量高、品质优等特点,在我国长江中下游地区尤其是江苏、浙江一带作为特色水生蔬菜广泛栽培[5]。
江浙地区菱的地方传统品种资源丰富[6-7],依果实形态分为五角菱、两角菱和四角菱3种类型,依果实颜色分为青菱和红菱[8],其中无角菱品种少,南湖菱即是其中一个比较特殊的品种[9]。近年来,受种植效益等因素影响,菱的种植面积急速下降,种质资源面临丢失风险。同时,品种间混杂现象越来越严重,例如,在南湖菱群体中,出现了有角的个体,地方品种的特色和优势难以维持,因此亟须开展种质资源的保护,并加以合理利用。种质资源鉴定是保护和利用的前提,但目前对菱的相关研究甚少。丁炳杨等[10]曾根据菱属植物花粉形态,将浙江省内的9个种菱属植物分为野菱类、乌菱类和细果野菱类,并提出野菱类是较乌菱类原始的一个类群,南湖菱在菱属植物中处在较高的进化水平,并认为南湖菱与四角菱亲缘关系更近。胡仁勇等[11]用31个性状将国内菱属37个居群分为3大类,其中9个栽培居群分在一类中,说明栽培菱品种之间的亲缘关系相对比较近;并将9个栽培品种分为3组,分别是四角菱一类,二角的乌菱和二角菱一类,无角的南湖菱单独一类,指出南湖菱与四角菱的关系更近。最近,丁炳扬等[12]把中国菱属分为细果野菱和欧菱2个种,并将欧菱划分为6个变种,将二角菱、乌菱统称为菱变种,将四角菱和南湖菱统称为四角菱变种。但以上分类,均是基于形态学的鉴定和分类结果,分子生物学的相关证据不足。
董晶莱等[13]利用DNA条形码技术对江浙不同产地的5份菱属植物进行了分子鉴定,发现菱栽培种间具有较高的稳定性。保曙琳等[14]利用rDNA ITS片段分析技术对长江中下游地区10个菱属居群进行DNA分子鉴别,表明菱属各居群间rDNA ITS的差异百分率较小,其亲缘关系较近,虽然根据ITS序列的特征可以较好地鉴定野生菱和栽培菱,但在栽培菱品种之间的亲缘关系及遗传距离仍未涉及。由于缺乏充分的基因组相关信息,目前对于菱属植物的遗传多样性尤其是栽培菱品种之间的遗传多样性的认识还是十分有限的,从而也限制了生产商对菱种质资源的开发和利用。利用现代分子生物学相关理论和技术明确栽培菱品种之间的亲缘关系,确定角的个数等形态学性状在亲缘关系远近的决定及菱的分类中的贡献,对于菱种质资源的保护、种质创新利用等均具有十分重要的意义。
单核苷酸多态性(single nucleotide polymorphism,SNP)是指在基因组水平上由单个核苷酸的变异所引起的DNA序列多态性,它是可遗传的变异中最常见的一种,由于其具有密度高、分布广、富有代表性,易实现自动化分析等特点,在遗传学分析中得到广泛应用。SNP分子标记技术,也以其高通量的特征,在分子遗传图谱构建、遗传多样性与种质鉴定、重要性状相关基因的定位和分子标记辅助选择等领域发挥出巨大的优势。其中,简化基因组测序SLAF-seq(specific-locus amplified fragment sequencing)是特异性位点扩增片段测序技术的简称,该技术降低了基因组的复杂度,并且不依赖参考基因组序列,成本低,成为SNP标记开发的首选[15]。通过SLAF-seq,在玉米[16]、沙冬青[17]、葡萄[18]、苹果属植物[19]、山西省地方梨的品种[20],以及高粱[21]等植物中,均已开发获得大量特异性SNP位点,并以SNP进行遗传分析,获得了种质资源遗传多样性的相关信息,明确了不同种质资源之间的亲缘关系。
本研究通过SLAF-seq技术,对江浙地区17份主要的菱栽培品种以及1份野生菱材料进行高通量测序,鉴定SNP,并利用获得的SNP数据进行群体多样性分析,包括进化树分析、群体结构分析及群体特异SNP标记的开发,并与其主要植物学性状分类进行相互验证,以期明确不同类型菱栽培品种之间的亲缘关系,为菱种质资源的鉴定保护和菱新品种的选育提供参考。
1 材料与方法
1.1 试验材料
18份菱材料均为嘉兴市农业科学研究院收集保存(表1)。于2020年上半年采用育苗移栽的方式种植,于3月初每个材料挑选10个健康菱种,进行点播,株距20 cm,每个材料播种一行。将面积为48 m2的水泥池用渔网分隔成4个面积12 m2的小区,于5月初每个材料选2株,分单株各种植2个小区。于8月下旬,菱结果期进行取样。
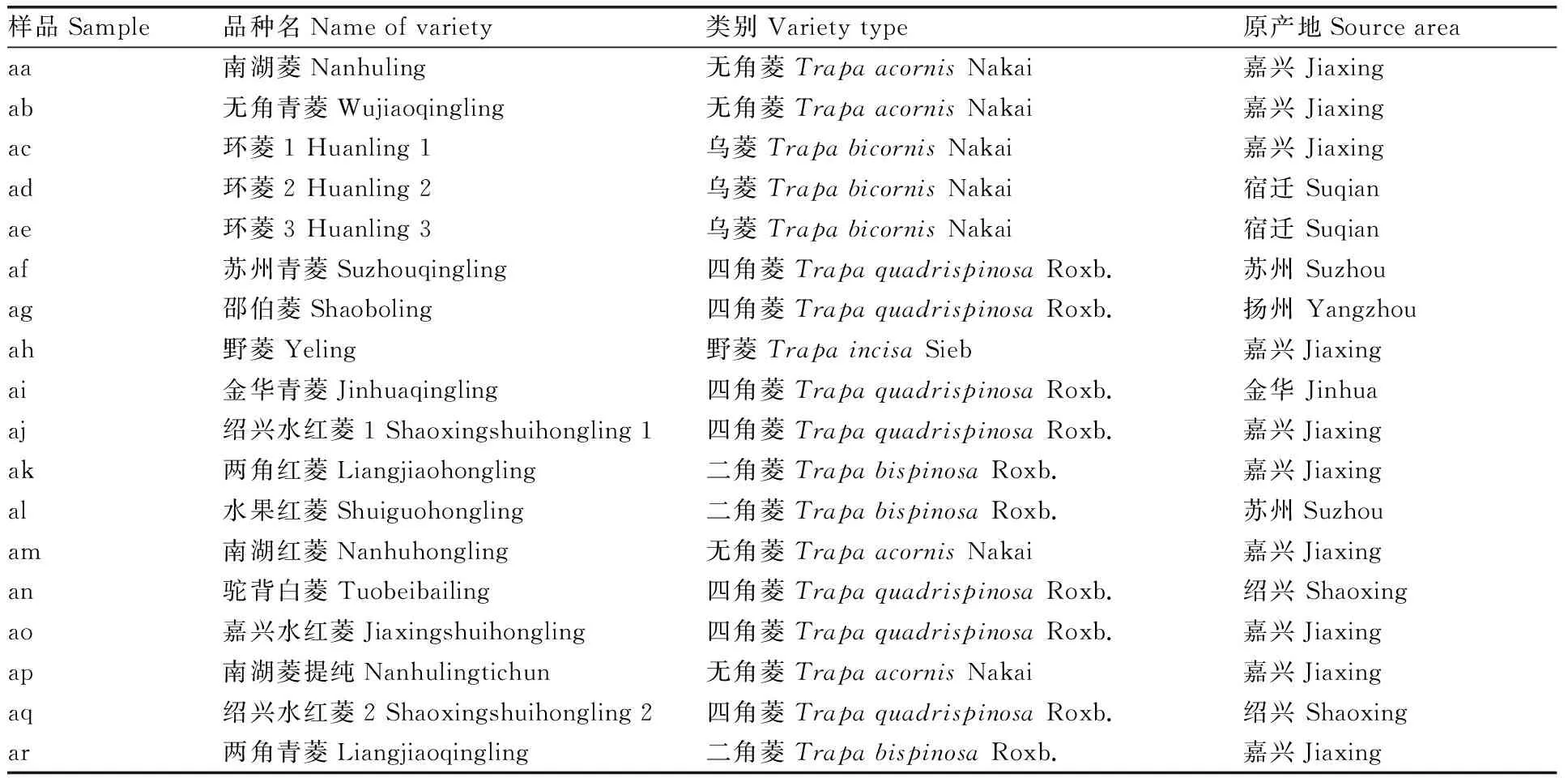
表1 供试菱资源及原产地明细Table 1 Resources and origin of Trapa L. in this study
1.2 主要农艺性状调查
菱生物学性状以菱初花期进行调查,菱果性状于盛产期进行调查。叶片于初花随机取15个菱盘,于每个菱盘中选最大叶进行测量,菱果数据为菱盛产期每种随机取15个菱果进行测定。测量数据采用DPS软件进行分析。
1.3 基因组DNA的制备
采用CTAB法提取菱样品的DNA,1%琼脂糖凝胶电泳检测DNA质量,Nanodrop检测DNA浓度,样品纯度D260/D280分布在1.8~2.0,所提取的样品DNA满足建库要求。
1.4 Illumina HiSeq测序及产出数据的质量分析
1.4.1 设计酶切方案
根据菱基因组大小及 GC 含量等信息,使用Hae Ⅲ+HinC Ⅱ酶切,酶切片段长度在364~464的序列定义为SLAF标签。
1.4.2 酶切片段测序分析
对得到的每一个样本基因组的酶切片段(SLAF标签)进行3签端加A处理、连接Dual-index测序接头[22]、PCR扩增、纯化、混样、切胶选取目的片段,文库质检合格后用Illumina平台IlluminaCasava 1.8进行测序。
1.5 SLAF 标签和SNP标记的开发
利用Dual-index对测序得到的原始数据进行识别,得到各个样品的reads。过滤测序reads的接头后,进行测序质量和数据量的评估。测序产生的reads来源于同一限制性内切酶对不同样品作用产生的长度相同的酶切片段,根据序列相似性将各样品的reads进行聚类,聚类到一起的reads来源于一个SLAF片段(SLAF标签)。同一SLAF标签在不同样品间的序列相似度远高于不同SLAF标签间的相似度;一个SLAF标签在不同样品间序列有差异(即有多态性),即可定义为多态性SLAF标签。
SNP标记的开发是以每个SLAF标签中深度最高的序列类型作为参考序列,利用BWA[23]将测序reads比对到参考基因组上,并使用GATK[24]和Samtools[25]两种方法开发SNP,以两种方法得到的SNP标记交集作为最终可靠的SNP标记数据集。
1.6 群体遗传分析
针对筛选的SNP对测序样本主要进行系统发育树以及群体遗传多样性分析,使用MEGA X[26]和Admixture软件[27]软件,对群体进行系统发育树和遗传结构分析。
2 结果与分析
2.1 测序数据统计与评估
本实验中HaeⅢ+HinC Ⅱ对菱基因组的酶切效率为90.59%。为保证分析质量,采用读长126×2作为后续的数据评估和分析数据。共获得菱reads数54.34 Mb,测序平均Q30为91.68%,平均GC含量为41.32%。所测序列的Q30数据较高,说明碱基出错率很低,测序结果可靠(表2)。
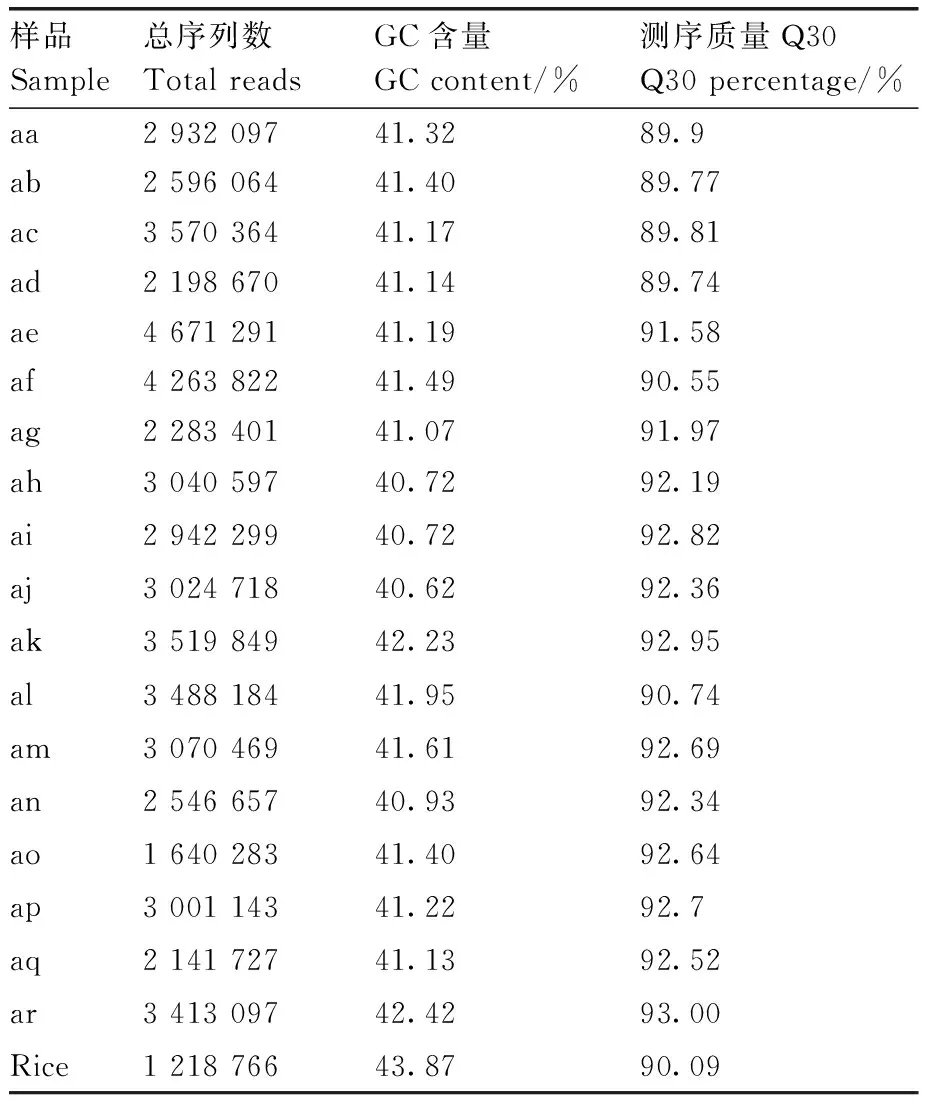
表2 菱各材料测序数据统计表Table 2 Reads statistics of Trapa L.
2.2 SLAF标签与SNP标记的鉴定
本研究在18个菱材料中共开发获得445 594个SLAF标签,标签的平均测序深度为14.10×,鉴定到多态性SLAF标签有95 931个(表3)。以每个SLAF标签中深度最高的序列类型作为参考序列进行SNP标记的开发,得到的SNP标记交集作为最终可靠的SNP标记数据集,共获得269 338个SNP标记(表4)。
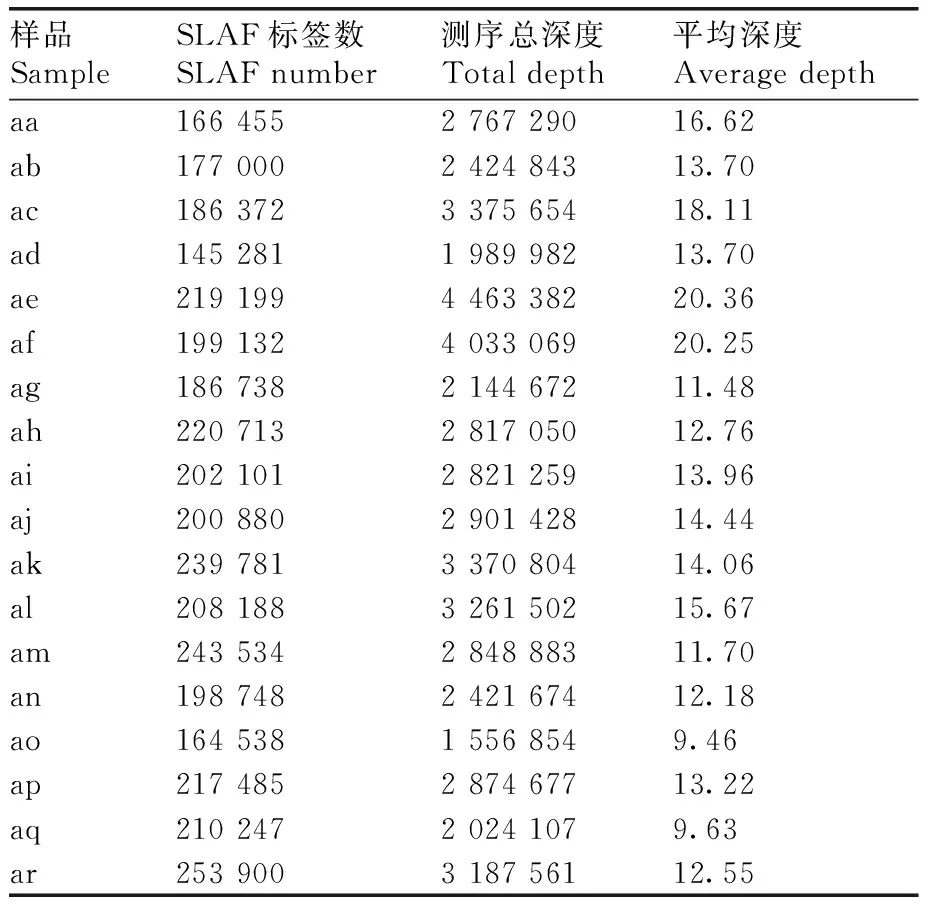
表3 菱各材料的SLAF标签统计Table 3 SLAF label statistics of Trapa L.
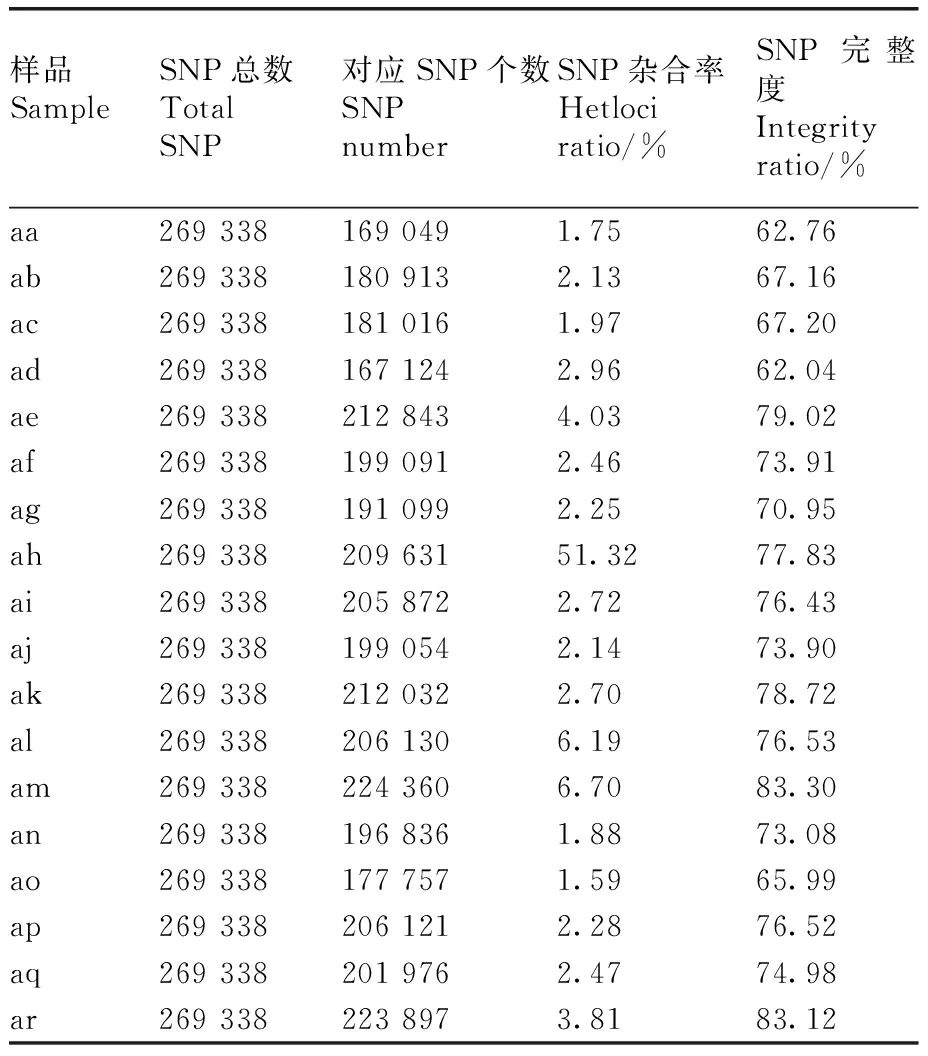
表4 菱各材料的SNP信息统计Table 4 SNP statistics of Trapa L.
2.3 遗传进化分析
基于过滤后得到的269 338个SNP,运用统计学方法,对18份菱完成系统进化树、群体结构,从基因组水平揭示不同个体菱的遗传分化关系。
2.3.1 系统发育分析
从系统发育树中可以看出(图1),18种菱可以划分为3个大类群:其中邵伯菱、南湖红菱、无角青菱、南湖菱和南湖菱提纯在大类群1;环菱1、环菱2、环菱3、野菱、苏州青菱、两角红菱、两角青菱、水果红菱在大类群2;金华青菱、绍兴水红菱2、驼背白菱、嘉兴水红菱、绍兴水红菱1 在大类群3;同一大类群的菱品种之间有更近的亲缘关系。在大类群1中,南湖红菱、无角青菱在遗传发育树上的距离更短,亲缘关系更近;南湖菱和南湖菱提纯亲缘关系更近。在大类群2中,环菱1、环菱3和环菱2的亲缘关系更近,环菱2与环菱3的亲缘关系又近于环菱1;野菱、苏州青菱、两角红菱、两角青菱、水果红菱的亲缘关系更近。在大类群3中,绍兴水红菱2 与驼背白菱的亲缘关系更近,嘉兴水红菱和绍兴水红菱1 的亲缘关系更近。以苏州青菱与野菱的亲缘关系最近,而南湖菱、南湖红菱、无角青菱、水红菱、驼背白菱等与野菱亲缘关系最远。因此,认为苏州青菱在进化上比较原始,而南湖菱等无角菱、水红菱等大果类型的四角菱进化程度更高。
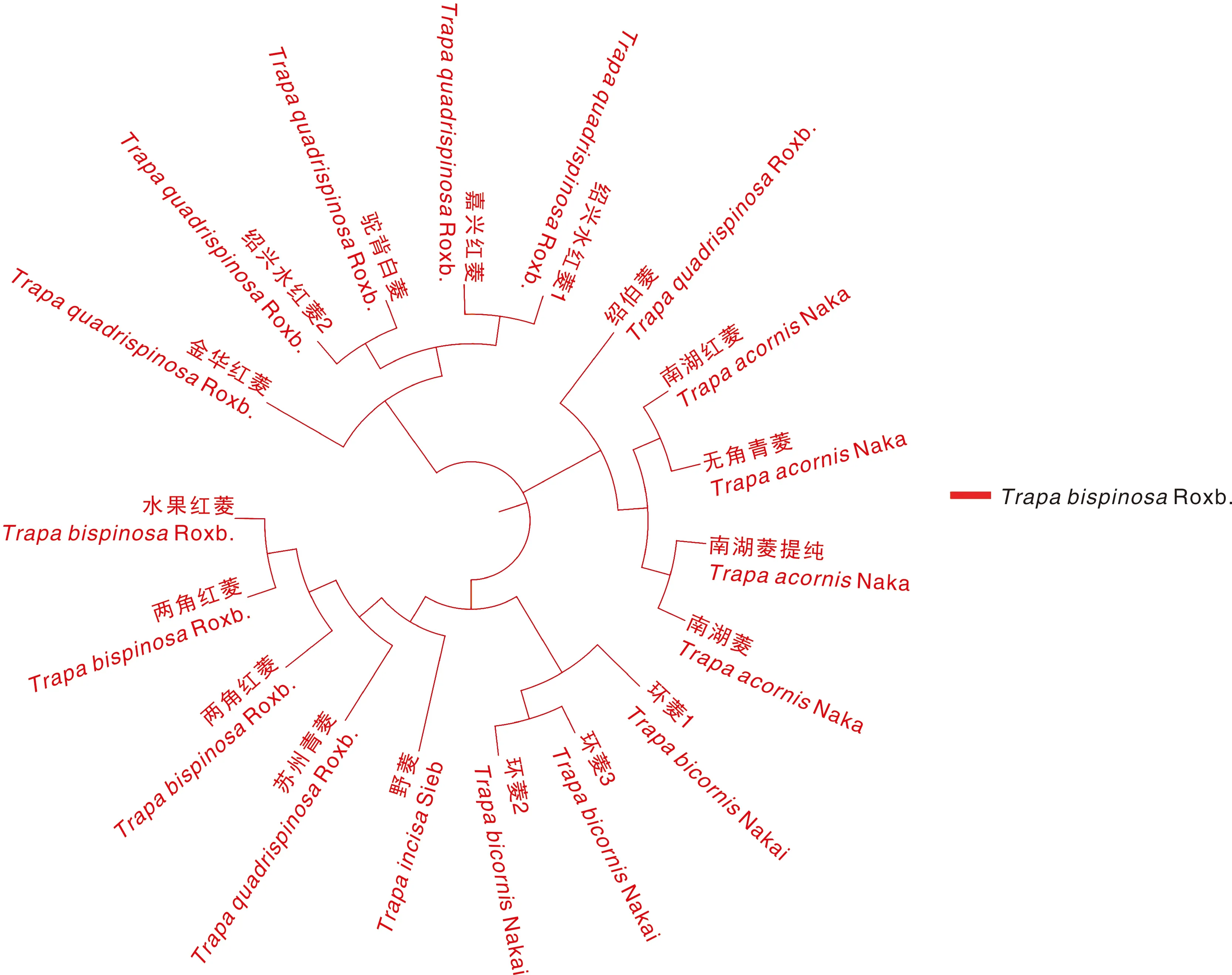
图1 十八个菱材料系统进化树Fig.1 Evolutionary tree of 18 Trapa L. samples
2.3.2 遗传结构分析
基于筛选的有效SNP,通过Admixture软件[27],分析样品的群体结构,分别假设样品的分群数(K值)为1~9,进行聚类。并对聚类结果进行交叉验证,根据交叉验证错误率的谷值(最低值)确定最优分群数。K值为1~9的聚类情况及各个K值对应的交叉验证错误率见图2。由图2-A可知,K=7时,交叉验证错误率最小,因此将18个样品分为7组。根据K=7时的结构图,同一类群用同种颜色表示,同一样本若具有不同颜色,则归到占比颜色最多的一组,详细见图2-B。
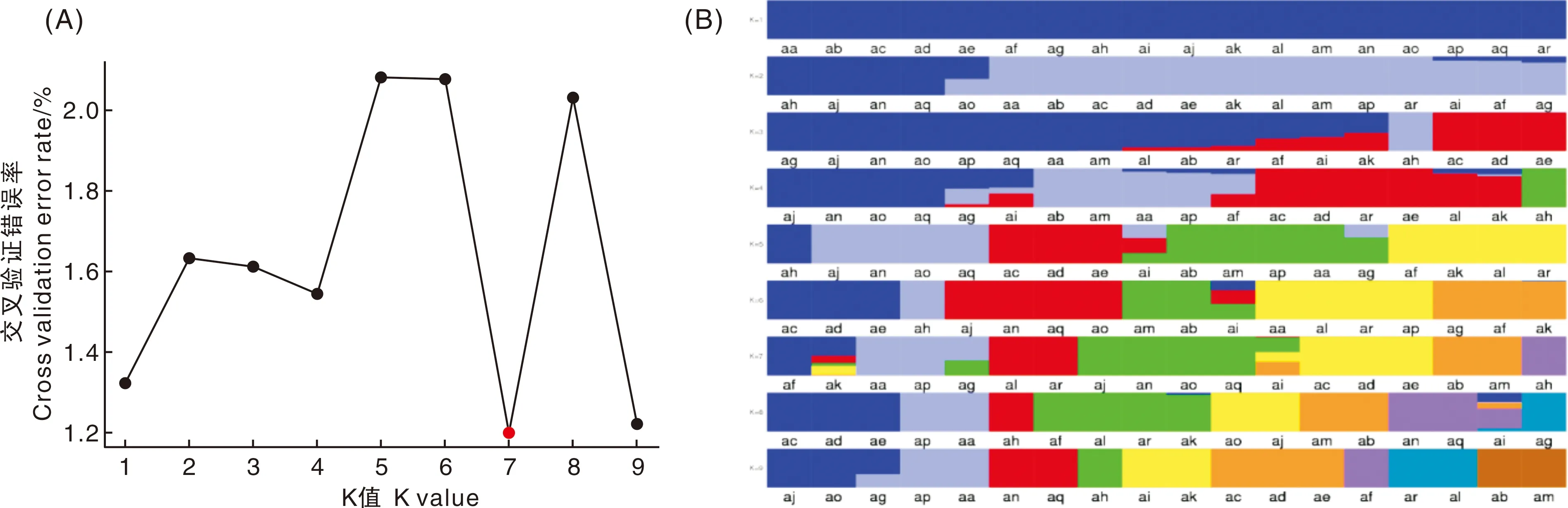
A, Admixture各个K值交叉验证错误率;B, Admixture各个K值对应的样品聚类结果。红框中为K=7的聚类结果图。A, Admixture of each K value cross validation error rate; B, Admixture of each K value corresponding sample clustering results. The red box is the clustering result graph with K=7.图2 十八个菱材料的遗传结构分析结果Fig.2 Genetic structure analysis of Trapa L. samples
交叉遗传错误率最低值出现在K=7时,因此可以将18份材料按照表5分为Q1~Q7一共7个组,理论上同一组的样本有共同祖先。Q1:苏州青菱、两角红菱;Q2:南湖菱、南湖菱提纯、邵伯菱;Q3:两角青菱、水果红菱;Q4:金华青菱、绍兴水红菱1、嘉兴水红菱、驼背白菱、绍兴水红菱2 ;Q5:环菱1、环菱2、环菱3;Q6:无角青菱、南湖红菱;Q7:野菱。

表5 菱材料遗传结构分群表Table 5 The table of the groups of Trapa L. samples classified by genetic structure
2.3.3 遗传多样性分析
遗传多样性分析结果显示,次要等位基因频率为0.354 9,期望等位基因数为1.805 9,与观测等位基因数(2.000 0)较为接近,表明等位基因在群体当中分布均匀;多样性指数为0.455,多态标记数为6 368,观测等位基因数为2;期望杂合度(0.441 5)高于观测杂合度(0.134 5),说明杂合个体较少,群体杂合度较低;多态性信息含量近0.342 7,香农维纳指数小于3,仅有0.632 4,说明本试验中的18个菱材料,其群体遗传变异不丰富。
3 讨论
水生蔬菜是江浙一带重要的特色经济作物,素有“水八仙”之称,而菱是其中最有特色的一个种类,在江浙一带有着广泛的分布和栽培。作为重要的蔬菜作物,菱在各个地方通过长期的演化形成了一些极具地方特色的栽培品种[6-7],比如嘉兴的南湖菱[9]、扬州的邵伯菱、太湖的水红菱、绍兴的驼背白菱,形成了当地著名的特色农产品。但近年来,菱栽培面积特别是传统外河湖荡种植面积大幅度下降,品种混杂退化严重,例如,南湖菱这种无角的、特异性状明显的菱品种,出现了不同程度的长角现象[28],另有一些地方品种如驼背白菱存在着消失的风险,地方特色菱种质资源的保护已刻不容缓。种质资源保护的前提是对种质资源进行鉴定,目前菱不同品种之间亲缘关系远近及遗传多样性仍不明确,并且可能存在品种间的混杂现象。本研究通过SLAF-seq技术对江浙地区主要的菱栽培品种进行简化基因组测序,基于过滤后得到的269 338个 SNP,运用统计学方法,对18份菱完成系统进化树、群体结构分析,从基因组水平首次揭示了不同菱的遗传关系,为了解菱的栽培演化提供了参考,也为菱品种的甄别及种质资源的鉴定保护提供了依据。
基于利用SLAF-seq技术获得的SNP进行遗传进化树及遗传结构分析来看,本试验中的18个菱品种,其群体遗传变异不丰富,说明本试验用样品虽然来源不同,但其之间亲缘关系较近,这与保曙琳等[14]的研究结果一致。本研究可以较好地将大部分无角菱、四角菱、二角菱区分开来,也能将菱栽培种与野菱区分开来,但遗传进化树中将四个无角菱类型品种与四角的邵伯菱分在一起,而将野菱与四角的苏州青菱与几个二角菱品种和乌菱分在一类,这与胡仁勇等[11]研究中9个栽培居群以菱果角的个数分成3大类有明显不同,这说明单纯的以形态学性状为依据进行菱栽培种的分类和亲缘关系认定存在着一定的局限性。同时遗传进化树显示,南湖菱以及以南湖菱为亲本选育的无角类型品种与四角的邵伯菱亲缘关系最近,进一步佐证了丁炳扬等[10,12]、胡仁勇等[11]提出的南湖菱与四角菱关系更加密切的观点。
综上,本论文从基因组的角度揭示了菱主要栽培品种之间的亲缘关系均较近,品种间亲缘关系与菱果的形状、颜色等果实性状无显著相关,南湖菱等无角菱品种与四角菱栽培种之间的亲缘关系更加密切。同时,从基因组信息角度验证了菱栽培种的演化可能是从四角野生菱资源同时向二角类型和大果型四角菱演化的过程,而无角菱栽培种是直接由四角菱演化而来。本实验结果为后续研究菱主要栽培品种的演化过程以及开展地方特色菱种质资源保护利用提供了理论依据。
