城市公园木本植物叶功能性状及其对固碳能力的驱动
2023-08-27张昊张兴高飞曲彦婷陈西林李雨桐李弘毅
张昊 张兴 高飞 曲彦婷 陈西林 李雨桐 李弘毅

摘要:了解城市木本植物叶水平固碳能力差异,从叶功能性状探讨对固碳能力的驱动路径,明确植物在长期生长过程中通过叶片性状来保持碳收获最大化的权衡策略。以城市公园20种常见木本植物为研究对象,测定其叶片碳储量(Cmass)、单位叶面积固碳量(WCO2)以及叶功能性状指标,以方差分析、相关性分析、路径分析和多元线性回归分析探究叶性状对固碳能力的影响。通过方差分析发现,木本植物的叶水平固碳能力具有较大差异,WCO2在4.70~26.06 g/(m2·d) 之间,平均为11.623 g/(m2·d),具有更明显的种间差异;Cmass在352.410~565.290 mg/g之间,平均为458.319 mg/g,具有更明显的种内差异。通过相关性分析发现,叶功能性状间存在显著的相关性,其中叶干质量(LDW)、叶干物质含量(LDMC)、叶厚度(LT)、比叶质量(LMA)、单位质量叶氮含量(Narea)、光合氮利用率(PNUE)、单位质量最大净光合速率(Amass)、气孔导度(Gs)等性状与WCO2和Cmass均极显著相关。通过路径分析和回归分析发现,单位面积最大净光合速率(Aarea)对WCO2具有直接驱动作用,可解释WCO255.4%的变化;碳氮比(C/N)、Amass、Nmass和PNUE等对Cmass具有直接驱动作用,可解释Cmass30.3%的变化。城市公园常见木本植物的叶功能性状可作为联系植物光合固碳与碳固存能力的载体,对植物固碳能力具有良好的指示作用。
关键词:木本植物;叶功能性状;固碳能力;驱动路径;城市公园
中图分类号:S684.01文献标志码:A
文章编号:1002-1302(2023)14-0173-07
人口高度密集、环境气候变化剧烈的城市,是二氧化碳(CO2)等温室气体的主要排放来源。公园绿地作为城市内具备直接碳汇价值的用地类型之一,能够通过植物群落进行自然固碳。其中,木本植物在碳循环中扮演了重要角色,其固碳能力的评价引起了人们的广泛关注[1-2]。叶片是所有植物器官中特征变化最显著的,也是植物进行光合作用固定CO2的主要场所,叶水平固碳能力是研究植株固碳能力的基础。目前,大部分研究是从环境土壤角度与植物生产力关系等方面展开的,对叶水平功能性状与初级生产力的关系等研究相对较少,且结论各不相同[3]。植物功能性状在反映植物获取、利用和保存资源等方面的能力已经得到公认,近年来已成为生态学和全球变化研究的热点[4]。植物叶功能性状对生态功能的变化有显著指示作用,可以将环境、植物个体和生态系统结构、过程与功能紧密地联系起来[5-6]。其中,比叶面积(SLA)、比叶质量(LMA)、叶干物质含量(LDMC)、叶片碳氮比(C/N)等是被广泛认为显著影响植物固碳能力的叶功能性状[7]。同时,光合氮利用效率(PUNE)、单位质量叶氮含量(Nmass)和单位面积叶氮含量(Narea)均对净光合速率(Pn)具有显著影响,共同体现植物的光合固碳能力[8]。因此,为了充分揭示影响叶功能性状驱动植物固碳能力的真实机理,不能仅将植物固碳能力的差异归因于单个或单类叶性状的差异,而应关注叶性状之间共同形成的耦合影响[9]。本研究选取江苏省苏州市城市公园20种木本植物,采用路径分析和线性回归模型,探讨能够影响及指征固碳能力的关键叶功能性状和驱动路径,综合考虑植物光合固碳能力与碳固存能力,以期为城市公园绿地木本植物质量的提升策略提供科学依据。
1 材料与方法
1.1 研究区域
江苏省苏州市位于30°47′~32°2′N、119°55′~121°20′E,地处温带,四季分明,屬北亚热带季风气候,年均降水量为1 100 mm,年均气温15.7 ℃,1月平均气温2.5 ℃,7月平均气温28 ℃。本试验选择苏州市建成时间较久且养护较好的18个城市公园作为样地,各类样地条件接近。
1.2 研究对象
在对苏州市城市公园深入调查的基础上,选择18个城市公园共有的20种常见木本植物作为研究对象(表1),共选取样本树种1 086株,每个树种不少于54株,含17科20属,包括常绿乔木、常绿灌木、落叶乔木和落叶灌木共4种生活型,所选木本植物均为生长旺盛且树龄相近的成年植株个体。
1.3 试验方法与数据分析
试验于2021年9—11月进行,样本采集时间在多数植物叶片物质积累最丰富的夏末秋初季节,各待测植株选取向阳面、光照度一致的当年生、完全伸展、无病虫害的成熟新鲜叶片,每个树种采集叶片数量大于200张。叶鲜质量的称质量方式为:去除叶片长柄,吸干叶片水分后将其放在电子天平(精确到0.000 1 g)上称量。将叶片置于80 ℃烘箱中杀青 3 h,用电子天平称量叶片干质量,按公式LDMC(g/g)=LDW(g)/LFW(g)计算叶干物质含量,其中:LDW为叶片干质量;LFW为叶片鲜质量。用叶面积扫描仪(MICROTEK ScanMaker i800plus)测定待测叶片的面积LA(cm2)。按公式LMA(g/cm2)=LDW(g)/LA(cm2)计算比叶质量,按公式SLA(cm2/g)=LA(cm2)/LDW(g)计算比叶面积。
在无风晴朗之日09:00—11:00采集叶片,使用光合仪(PPsystem,LiCor6400XT,USA)测定净光合速率、蒸腾速率(Tr)和气孔导度(Gs),光源选择LED红蓝光源,光照度设定为1 500 μmol/(m2·s),CO2浓度设定为400 μmol/L,叶室温度设定为30 ℃,测得单位面积最大净光合速率Aarea[μmol/(m2·s)],计算单位质量最大净光合速率Amass[μmol/(g·s)],计算公式为Amass=Aarea×SLA÷10 000。随后在相同环境条件下,使用便携式叶绿素仪(TYS-B)测定植物叶片SPAD值,并使用元素分析仪(EURO EA3000)测定植物叶片中的碳氮含量,以此来计算叶片单位质量氮含量Nmass和单位叶面积氮含量Narea(g/m2),计算公式为Narea=Nmsaa/SLA×10。叶片的光合氮利用效率[μmol/(g·s)]计算公式为PNUE=Amass/Narea。
目前广泛应用于确定植物固碳能力的研究方法主要有2种,一是同化法(或称过程法),即测定植物叶片净光合速率,推算出单位叶面积固碳量(WCO2),侧重强调植物通过光合作用在一段时间内产生有机物的量来评价植物的光合固碳能力;二是生物量法(或称收获法),在植物生物量测定的基础上直接测定植物器官的碳含量即单位质量叶碳含量(Cmass),侧重强调植物在一定时刻储存在器官中的有机物的量来评价植物的碳固存能力[10]。本研究综合考虑2种固碳能力评价方法。
植物叶碳含量根据植物的净光合速率计算,采用同化量法,测定当天各树种的净同化量的计算公式为
式中:P表示同化总量,mmol/(m2·s);Pt表示初测点的瞬时净光合速率;Pi表示下一测点的瞬时净光合速率,μmol/(m2·s);ti表示初测点的瞬时时间,h;ti+1表示下一测点的时间,h;j表示测试次数;3 600 表示3 600 s/h;1 000表示“1 mmol为 1 000 μmol”。
用测定日的同化总量换算为测定日固定量为
WCO2=P×(1-0.2)×44/1 000。
式中:44表示CO2的摩尔质量;WCO2表示单位面积叶片固定CO2的质量,g/(m2·d)。
1.4 分析方法
本研究对所采集的样本进行试验测定,试验指标包括16个叶功能性状与2个固碳能力指标,所得数据采用SPSS 22软件进行分析。采用方差分析探究20种木本植物功能性状间的差异以及光合固碳能力、碳固存能力差异,采用相关性分析研究木本植物叶功能性状与固碳能力的相关关系。已有研究结果表明,当研究变量间具有大量复杂的因果关系时,路径分析可以解决多元回归方程中无法关注自变量间相关关系的问题,对于验证多因素耦合影响过程具有巨大优势[11]。因此,本研究采用路径分析和线性回归模型共同探讨叶功能性状对固碳能力的驱动路径。
2 结果与分析
2.1 城市公园树种固碳能力差异性与叶性状差异性
2.1.1 固碳能力差异性分析
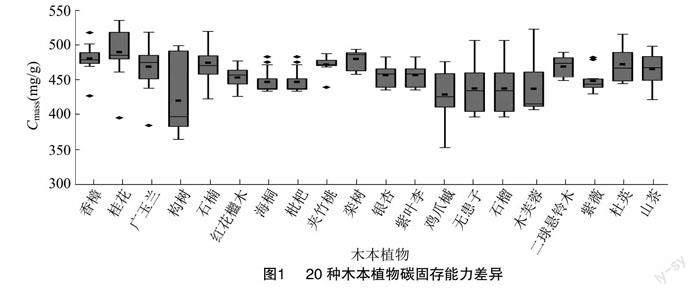
不同树种由于自身生理特性不同,其固碳释氧能力存在一定的差异性。20种木本植物叶水平固碳能力差异明显,且碳固存能力具有更明显的种内差异(图1),光合固碳能力具有更明显的种间差异(图2)。木本植物单位叶面积固定量在4.70~26.06 g/(m2·d)之间,平均为11.623 g/(m2·d);单位质量叶碳含量在352.410~565.290 mg/g之间,平均为458.319 mg/g。
2.1.2 叶功能性状差异性分析
在20种木本植物及其16个叶经济谱性状的数据中,各性状间差异明显(表2)。在统计分析的16个性状中总体变异系数范围较大,为20.209%~123.690%,其中变异系数最大值出现在Tr,之后是LA,分别为123.690%、118.999%,说明两者携带的物种信息较多;变异系数较小值是SPAD与LDMC,分别为20.209%、24.837%,说明二者是资源轴上较稳定的变量。
2.2 叶功能性状间及其与固碳能力的相关性分析
2.2.1 叶功能性状间的相关性分析
本研究木本植物的叶功能性状之间普遍存在极显著关联,其中大部分与全球尺度一致[12](图3)。具有极显著正相关关系(P<0.01)的有:C/N和PNUE;LA分别和LFW、LDW、SLA、PNUE、Amass、Aarea、Gs、Tr;LFW分别和LDW、Aarea、Gs、Tr、SPAD;LDW分别和GS、SPAD;LDMC和Narea;LT和LMA、Narea、Aarea、SPAD;SLA和Nmass、PNUE、Amass;LMA分別和Narea、Aarea、SPAD;Nmass分别和Narea、Amass;Narea和Aarea、SPAD;PNUE和Amass、Aarea、Gs、Tr;Amass和Aarea、Gs、Tr;Gs和Tr、SPAD。
具有极显著负相关关系(P<0.01)的有:C/N分别和SLA、Narea、Nmass、Amass;LA和LT、LMA、Narea;LDMC分别和LT、SLA、PNUE、Amass、Aarea;LT分别和SLA、PUNE、Amass、Tr;SLA和LMA、Amass、SPAD;LMA和PNUE、Amass、Tr;Nmass和PNUE;Narea和PNUE、Amass、Gs、Tr;PNUE和SPAD;Amass和SPAD。
2.2.2 叶功能性状与固碳能力的相关性分析
对20种木本植物的叶性状和叶片固碳能力进行相关性分析,其中LA、LFW、LDMC、LT、Tr、PNUE、Amass、Aarea、Gs和SPAD等叶功能性状均与WCO2之间产生极显著相关性(P<0.01),LDW、LMA、Narea与WCO2之间产生显著相关性(P<0.05),SLA、Nmass和Tr等3个性状与WCO2间不具有明显相关性;C/N、LDMC、LT、SLA、LMA、Amass、Gs和SPAD等叶功能性状与Cmass产生极显著相关性(P<0.01),LDW、Nmass、Narea、PNUE与Cmass产生显著相关性(P<0.05),LA、LFW、Narea、PUNE、Aarea、Tr等5个性状与Cmass间不具有明显相关性。结果表明,WCO2和Cmass在叶性状间既存在共同相关关系,也存在较大差异(表3)。
2.3 叶功能性状对固碳能力的影响性分析
2.3.1 路径分析
本研究叶功能性状之间互相关联显著,且对于固碳能力具有明显的影响关系,可作为连系WCO2和Cmass的桥梁,因此选择路径分析以明确叶功能性状的影响关系并说明变量间的直接效应和间接效应(图4)。结果表明,WCO2和Cmass并不存在直接连系,但二者均受到相关性状的直接驱动。其中,Aarea对WCO2产生直接正向驱动,而LA、LFW、LDW、LDMC、LT、LMA、Tr、Narea、PNUE、Aarea、Gs和SPAD等相关性状并不会对WCO2产生直接驱动;Nmass、C/N和Amass对Cmass产生直接正向驱动,PUNE对Cmass产生直接负向驱动,而LDMC、LT、SLA、LMA和Gs等相关性状并不会对Cmass产生直接驱动。
2.3.2 多元线性回归分析
上述结果表明部分叶功能性状可直接驱动固碳能力,为进一步明确叶功能性状在路径中的影响程度,将叶功能性状分别对Cmass和WCO2进行方程拟合(图5)。
将路径分析中极显著的叶功能性状作为自变量,将Cmass作为因变量进行线性回归分析,得到模型公式为Cmass=243.274+3.646×Nmass+166.292×Amass+6.426×C/N-4.856×PUNE,模型R2值为0.303,(F=38.624,P=0.000**<0.05);将路径分析中极显著的叶功能性状作为自变量,将WCO2作为因变量进行线性回归分析,得到模型公式为WCO2=3.966+0.475×Aarea,模型R2值为0.554,(F=445.588,P=0.000**<0.05)。
3 讨论与结论
3.1 固碳能力与叶功能性状特征分析
本研究中Tr和LA的种间变异较大,SPAD和LDMC的种间变异较小,这与前人的研究结果基本一致,说明蒸腾作用在木本植物种间具有较大的差异和良好的指示作用[13-14]。进而可知,木本植物通过增加LA来弥补由于SLA减小而导致的碳摄取效率损失的生理现象在种间较显著,且值得进一步探讨。而LDMC种间差异较小,作为反映植物对养分元素保有能力的重要指標,表明城市公园木本植物处于适宜的气候和良好的水土环境[15]。
本研究选取的20种园林木本植物中WCO2和Cmass均存在不同程度的差异。光合固碳能力强调资源获取,具有更明显的种间差异;碳固存能力侧重资源利用储存,具有更小的种间差异和更大的种内差异。这进一步说明光合固碳能力在指征植物自身物种特性时可能具备更大的优势,而碳固存能力作为跨种间的研究指标可能会更加稳定。
3.2 叶功能性状与固碳能力的相关关系
本研究中的SLA与LDMC呈极显著负相关(P<0.01)、LMA与LDMC呈极显著正相关(P<0.01),这与全球叶性状之间的相关关系基本相同。叶片生理性状可以反映植物的生长代谢情况,本研究中Tr和Gs呈极显著正相关(P<0.01),与LMA呈极显著正相关(P<0.01),这与多数研究一致,展现了树种可以通过较强的光合能力节约叶形态投入的适应策略。
本研究进一步印证了张林等多位学者的研究结果,即植物叶性状与固碳能力之间具有相关性,部分性状可在一定程度上表示植物的碳同化能力,如SLA大、Nmass高的植物其叶片净光合速率大,能有效进行光合作用并快速储存周边环境中的养分[16-18]。
3.3 叶功能性状对固碳能力的驱动路径
在路径分析中发现,SPAD、C/N、Aarea、Nmass、PUNE、Amass和SLA等涉及到叶片生理、化学和结构性状的7个叶功能性状与WCO2和Cmass具有显著影响性。Aarea和Amass在WCO2和Cmass之间显示出明显分异,即WCO2与叶功能性状中的面积归一化联系更紧密,Cmass与叶功能性状中的质量归一化更密切,印证了2种假说在联系植物固碳能力时可能独立存在[19]。其中,WCO2仅受Aarea直接驱动,表明叶功能性状在植物光合固碳过程中变化复杂。本研究中PUNE和SLA并未对WCO2产生直接驱动作用,说明二者虽可以较好反映植物的光合固碳能力,但也可能更易受到如Nmass、SPAD和Aarea等性状的间接驱动,而PUNE对Cmass形成显著的负向路径,说明在评价植物碳固存能力时植物本身的PUNE不可忽视。此外,在7个叶功能性状中SLA作为较易测得的叶性状指标,对Cmass和Aarea具备明显的驱动效应,表明其具备一定的直接指示价值。
为进一步明确植物在长期生长过程中通过叶片性状来保持碳收获最大化的权衡策略,本研究通过回归分析对比得出WCO2和Cmass均可以和叶功能性状形成良好的线性关系,其中C/N、Amass、Nmass、PUNE共同阐释了树种的碳同化差异并直接反映其最大化构建途径:即较高C/N、Amass、Nmass和较低PUNE的植物叶碳固存能力更强;较高Aarea的植物叶光合固碳能力更强,而SPAD和Nmass的直接影响并不显著,这可能与SPAD和Nmass较小的种间差异有关。
3.4 结论
WCO2和Cmass与叶功能性状直接相关,其中Narea没有和WCO2形成明显驱动关系,这可能是由于本研究所选木本植物涉及科属较多,其叶片氮分配及CO2扩散阻力的差异较大[20]。SPAD均与WCO2及Cmass产生极显著相关关系,但均未与WCO2和Cmass形成明显驱动路径,也可能是由于本研究中部分树种冠高较高,为避免上层叶片光损伤,其通过降低光吸收并增强氮元素及叶绿素利用能力的碳权衡现象较显著[21]。因此,本研究所提出的叶功能性状对固碳能力的驱动路径是否会在植物生活型种类、绿地类型之间产生结果差异还需进一步研究。总而言之,SPAD、C/N、Aarea、Nmass、PNUE、Amass和SLA等叶性状之间明确的权衡关系共同表明,植物固碳能力不仅取决于净光合速率或叶片质量,而是由多种叶功能性状共同权衡决定的,但这种驱动关系在非城市公园等人工扰动较低的环境及环境胁迫中,是否存在或发生改变尚不明确[22-24]。因此,为全面了解植物功能性状对固碳能力的驱动路径,在后续的研究中可选择实验室环境加以验证,并针对不同生活型植物及不同生长环境、整株水平(包括根、茎、枝、叶柄和叶)进行更多的系统研究。
参考文献:
[1]李宸宇,朱建华,张 峰,等. 基于NbS的北京市乔木林固碳能力分析[J]. 北京林业大学学报,2021,43(6):13-22.
[2]郭 晖. 河南省郑州市公园绿地常见植物固碳释氧能力分析[J]. 江苏农业科学,2017,45(15):103-106.
[3]曹小玉,赵文菲,李际平,等. 中亚热带几种典型森林土壤养分含量分析及综合评价[J]. 生态学报,2022,42(9):3525-3535.
[4]陈莹婷,许振柱. 植物叶经济谱的研究进展[J]. 植物生态学报,2014,38(10):1135-1153.
[5]张德顺,刘 鸣. 基于“植物功能性状-生态系统服务”评价框架的园林树种选择方法——以上海为例[J]. 中国园林,2020,36(2):106-111.
[6]Wright I J,Reich P B,Westoby M,et al. The worldwide leaf economics spectrum[J]. Nature,2004,428(6985):821-827.
[7]Ishida A,Nakano T,Yazaki K,et al. Coordination between leaf and stem traits related to leaf carbon gain and hydraulics across 32 drought-tolerant angiosperms[J]. Oecologia,2008,156(1):193-202.
[8]陈高路,陈 林,庞丹波,等. 贺兰山10种典型植物固碳释氧能力研究[J]. 水土保持学报,2021,35(3):206-213,220.
[9]张 磊,吕光辉,蒋腊梅,等. 干旱区荒漠植物初级生产力及碳储量与功能性状之间的关系[J]. 新疆大学学报(自然科学版),2020,37(1):1-12.
[10]张 颖.建筑垃圾堆山公园植物群落固碳效益量化与优化研究——以天津南翠屏公园为例[D]. 天津:天津大学,2019.
[11]Yu M F,Pan Y T,Yang L L,et al. Response of sediment microbial communities to the rural wastewater in the pond-ditch circulation system[J]. Frontiers in Environmental Science,2021,9:717458.
[12]Osnas J L D,Lichstein J W,Reich P B,et al. Global leaf trait relationships:mass,area,and the leaf economics spectrum[J]. Science,2013,340(6133):741-744.
[13]張姗姗,张 兴,曲彦婷,等. 留园植物叶性状及其叶经济谱研究[J]. 北方园艺,2022(14):57-65.
[14]Valencia E,Gross N,Quero J L,et al. Cascading effects from plants to soil microorganisms explain how plant species richness and simulated climate change affect soil multifunctionality[J]. Global Change Biology,2018,24(12):5642-5654.
[15]靳 莎,闫淑君,黄柳菁,等. 植物叶功能性状间的权衡研究进展[J]. 四川林业科技,2019,40(5):96-103.
[16]张 林,罗天祥.植物叶寿命及其相关叶性状的生态学研究进展[J]. 植物生态学报,2004,28(6):844-852.
[17]宋 贺,于鸿莹,陈莹婷,等. 北京植物园不同功能型植物叶经济谱[J]. 应用生态学报,2016,27(6):1861-1869.
[18]庞志强,卢炜丽,姜丽莎,等. 滇中喀斯特41种不同生长型植物叶性状研究[J]. 广西植物,2019,39(8):1126-1138.
[19]何芸雨,郭水良,王 喆.植物功能性状权衡关系的研究进展[J]. 植物生态学报,2019,43(12):1021-1035.
[20]黄 凯. 物种和群落水平上外来入侵植物与共生本地植物叶功能性状和养分循环的差异及环境的影响[D]. 沈阳:沈阳农业大学,2021.
[21]郭美玲,姚步青,石国玺,等. 高寒草甸植物叶片碳含量及其可塑性与系统发育的关系[J]. 生态学杂志,2018,37(6):1841-1848.
[22]王園博,赵锐锋,张丽华,等. 黑河中游湿地不同植物群落土壤有机碳分布及影响因素[J]. 草业科学,2020,37(5):833-844.
[23]陈 珊,张 兴,曲彦婷,等. 石湖园林植物LES性状对水分环境响应的研究[J]. 东北农业大学学报,2021,52(6):34-44,77.
[24]Gago J,de Menezes D D,Figueroa C M,et al. Relationships of leaf net photosynthesis,stomatal conductance,and mesophyll conductance to primary metabolism:a multispecies meta-analysis approach[J]. Plant Physiology,2016,171(1):265-279.
收稿日期:2022-11-02
基金项目:黑龙江省科学院双提雁阵项目(编号:STYZ2022ZR01);江苏省研究生科研创新计划(编号:KYCX21_3054);宿迁交通科研基金(编号:2022Y11);风景园林学“十四五”江苏省重点学科、江苏省研究生工作站共同资助项目(编号:082240006/002/003)。
作者简介:张 昊(1997—),女,河北邢台人,硕士研究生,主要从事地域生态环境与景观规划研究。E-mail:29569116@qq.com。
通信作者:张 兴,教授,研究员,主要从事地域生态环境与景观规划研究。E-mail:2605@usts.edu.cn。
