多样化种植对土壤团聚体组成及其有机碳和全氮含量的影响
2023-08-15孙涛冯晓敏高新昊邓艾兴郑成岩宋振伟张卫建
孙涛,冯晓敏,高新昊,邓艾兴,郑成岩,宋振伟,张卫建
多样化种植对土壤团聚体组成及其有机碳和全氮含量的影响
孙涛1,3,冯晓敏2,3,高新昊1,邓艾兴3,郑成岩3,宋振伟3,张卫建3
1养分资源高效利用全国重点实验室/农业农村部废弃物基质化利用重点实验室/山东省农业科学院农业资源与环境研究所,济南 250100;2山西农业大学高粱研究所,山西晋中 030600;3中国农业科学院作物科学研究所/农业农村部作物生理生态重点实验室,北京 100081
【目的】阐明东北黑土区禾本科与豆科作物多样化种植模式下土壤团聚体组成及其碳氮分布特征,为促进黑土地用养结合型种植制度优化提供理论依据与技术指导。【方法】田间试验于2016—2020年在中国农业科学院作物科学研究所公主岭试验站进行,设置玉米大豆间作(M/S)、玉米大豆轮作(M-S)、玉米花生间作(M/P)和玉米花生轮作(M-P)等多样化种植模式,以玉米连作(CM)常规种植模式为对照。2020年作物收获后采集0—20和20—40 cm土层土壤样品,分析多样化种植模式对土壤团聚体及其有机碳、全氮含量的影响。【结果】多样化种植有利于增加0—20和20—40 cm土层水稳性大团聚体(>0.25 mm)含量,降低黏粉粒(<0.053 mm)含量,4种多样化种植模式的土壤团聚体平均重量直径和平均几何直径均显著高于CM处理(<0.05)。M/S和M-S处理0—20 cm土层>0.25 mm土壤团聚体比例分别比CM处理高17.6%和13.4%,M/S、M-S和M-P处理20—40 cm土层 >0.25 mm土壤团聚体比例则分别比CM处理高10.4%、8.3%和10.5%。多样化种植增加了土壤大团聚体中有机碳和全氮含量,0—20 cm土层,M/S、M-S、M/P和M-P处理>2 mm土壤团聚体有机碳含量分别比CM处理高20.7%、24.3%、18.8%和17.8%;0—20 cm土层,M-S、M/P和M-P处理>2 mm土壤团聚体全氮含量分别比CM处理高13.0%、16.8%和14.8%。M-S和M/P处理0—20 cm土层>2 mm土壤团聚体有机碳和全氮对土壤总有机碳和全氮贡献率高于CM处理,而M/S和M-S处理<0.053 mm土壤团聚体有机碳和全氮对总有机碳和全氮的贡献率则低于CM处理。【结论】东北黑土区多样化种植提高了土壤大团聚体含量,降低了黏粉粒含量,增加了土壤团聚体稳定性及有机碳和全氮含量,有利于促进农田土壤碳氮固持。
东北黑土区;轮作;间作;玉米;大豆;花生;土壤团聚体;土壤有机碳;土壤全氮
0 引言
【研究意义】多样化种植是指通过间作、轮作和覆盖作物的方式在农田中同时或按一定时序种植多种作物的生产方式。与集约化农业相比,多样化种植利用生态位分化和物种互作的正效应提高粮食产量和养分利用效率,被认为是改善耕地质量和保障国家粮食安全的重要措施[1]。【前人研究进展】大量研究表明,多样化种植可以增加农田生物多样性和稳定性[2],提高作物产量[3-4],改善土壤肥力[5-6],提高土地利用率[7],增强作物系统对光、温、水、肥的利用[8]。土壤有机碳与全氮作为衡量土壤肥力的重要指标,与作物养分供给、土壤持水、生物多样性等具有显著的正向作用[9]。种植模式、耕作制度、养分管理等均会影响土壤有机碳和氮含量[10]。前人研究表明,多样化种植由于增加了输入土壤中的凋落物种类和数量,有利于促进土壤有机碳和全氮的积累[11]。也有相关研究认为,多样化种植体系下,进入土壤的有机碳源分解速率高,不利于土壤碳氮的固持[12]。由此可见,有关多样化种植对土壤有机碳和全氮的影响仍存在不确定性,亟需进一步深入研究。土壤团聚体作为土壤结构的基本单元,对维系土壤结构具有重要作用[13]。土壤团聚体含量及粒级分布不仅影响作物生长发育,而且对土壤的一系列物理、化学及生物学过程有重要影响。大量研究表明土壤团聚体对有机碳的固持起着非常重要的作用[14-15]。作物残茬输入土壤后首先在大团聚体中停留[16],土壤大团聚体更容易使微生物接触有机物,从而导致大团聚体有机碳的矿化程度高于微团聚体。有机碳的矿化同时会释放出氮、磷、硫等活性养分[17-18],而微团聚体中的胶体及无机物质与有机碳紧密结合,为有机碳的固持提供了物理保护,使其成为土壤有机碳的重要固持场所[19]。已有研究表明多样化种植可以增强土壤有机碳氮含量及土壤团聚体稳定性[5,11],但受土壤类型、气候特征以及作物搭配种类等影响,不同多样化种植模式对土壤团聚体有机碳和全氮含量的影响及其差异仍需深入研究。【本研究切入点】东北黑土区作为我国最重要的粮食主产区和商品粮供应基地,玉米种植面积和产量均占全国1/3。但长期以来种植制度单一,导致农田生物多样性下降,土壤养分消耗失衡,破坏了土壤生态系统稳定性,土壤中长期大量积累病原物,导致特定土传病虫害加剧,加之缺少耕地质量恢复和保护性措施,黑土耕地质量呈现逐年下降的趋势[20-21]。近年来,随着国家黑土地保护与利用战略的全面实施,以禾本科和豆科作物为主的多样化种植模式在东北黑土区逐渐受到重视,如玉米和大豆、花生等作物的轮作及间作种植模式等。但是多样化种植在保障高产与资源高效利用的同时,其对土壤团聚体结构以及土壤团聚体有机碳、全氮含量的影响仍不清楚,且不同多样化种植模式的差异性也有待研究。【拟解决的关键问题】因此,本研究依托2016年在吉林省公主岭市建立的多样化种植模式田间定位试验,系统分析不同模式下土壤团聚体结构及其有机碳氮含量和贡献率变化特征,以期为东北黑土区不同多样化种植模式下土壤有机碳氮固持特征研究及多样化种植模式推广应用提供依据。
1 材料与方法
1.1 试验地概况
田间试验于2016—2020年在中国农业科学院作物科学研究所公主岭试验站进行(43°31′N,124°48′E,海拔220 m)。试验地所在区域为温带大陆性季风气候,年平均气温5.6 ℃,年平均日照时数为2 710 h,>10 ℃积温为3 180 ℃,年无霜期为144 d,年平均降雨量为562 mm。土壤类型为黑土,0—20 cm土层容重1.38 g·cm-3,有机碳16.3 g·kg-1,pH 7.6,全氮含量1.5 g·kg-1,全磷含量0.6 g·kg-1,全钾含量18.4 g·kg-1,速效氮含量143.3 mg·kg-1,速效磷66.5 mg·kg-1,速效钾150.8 mg·kg-1。
1.2 试验设计及田间管理
田间试验设置玉米连作(CM,对照)、玉米与大豆间作(M/S)、玉米与大豆轮作(M-S)、玉米与花生间作(M/P)、玉米与花生轮作(M-P)5个处理。试验采用随机区组设计,每个处理3次重复。每个小区面积96.0 m2(10.0 m×9.6 m)。每年播种前用旋耕机进行旋耕整地,深度为15 cm。3种作物均采用等行距种植方式,其中单作和轮作处理玉米行距为60 cm,株距24.6 cm,种植密度6.75万株/hm2;大豆行距60 cm,株距15 cm,每穴两粒,种植密度22.5万株/hm2;花生行距60 cm,株距13.9 cm,种植密度12.0万/hm2。玉米与大豆、花生间作处理,玉米行距60 cm,株距18.5 cm,种植密度9.00万株/hm2;间作处理大豆和花生行距、株距和种植密度与单作和轮作处理一致。间作处理采用条带种植的方式,玉米和大豆、花生条带宽度为480 cm。每条带包括240 cm的玉米带(4行)和240 cm大豆或花生带(4行),间作模式中玉米和大豆、花生在年际间进行交替轮作。玉米与大豆、花生间行距为60 cm。不同种植模式每年玉米、大豆和花生均在4月底播种,9月底收获。玉米、大豆和花生品种分别为先玉335、吉育321和吉花19。玉米、大豆和花生施肥量均为165 kg N·hm-2、82.5 kg P2O5·hm-2和 82.5 kg K2O·hm-2。肥料中50%的氮肥和全部的磷钾肥作为基肥在旋耕整地时施用,50%氮肥在6月中下旬作为追肥施用,整个生育期不灌水,玉米、大豆和花生秸秆在收获时均移出田外。
1.3 测定指标与方法
1.3.1 土壤样品采集 在2020年10月,作物收获时采集0—20和20—40 cm土层土壤和土壤团聚体样品。土壤样品采用土钻采集。土壤团聚体采用直径8 cm铝盒采样,将铝盒倒扣在取样位置,缓缓压入土壤中,待铝盒内土壤样品充满后,移出采样铝盒,带回实验室沿自然断裂面掰成直径1 cm土块,在阴凉通风处自然晾干,留待团聚体粒级及有机碳和全氮含量测定。单作和轮作处理,每个小区在作物行间采集3个样点,混为一个土样,间作处理在每种作物行间和两种作物行间分别采集3个样点,混匀成为一个土样。
1.3.2 土壤团聚体测定 土壤水稳性团聚体采用湿筛法测定。将风干后的土壤样品干筛,然后按照干筛各粒级比例称取土壤样品50 g,倒入孔径分别为2、0.25和0.053 mm的筛组中进行振荡筛分5 min(30 次/min),将土壤团聚体分为>2 mm、0.25—2 mm、0.053—0.25 mm和<0.053 mm 4个粒级,最后将各筛子中的团聚体用去离子水洗入铝盒中,60 ℃烘干至恒重后称重,计算各粒级土壤团聚体质量百分比。土壤团聚体根据其粒级的大小可划分为大团聚体(> 0.25 mm)和微团聚体(<0.25 mm)。
1.3.3 团聚体有机碳和全氮 团聚体有机碳含量采用重铬酸钾外加热法测定,全氮含量采用凯氏定氮法测定[22]。
1.3.4 数据计算 土壤团聚体平均重量直径(mean weight diameter,MWD)、平均几何直径(geometric mean diameter,GMD)和>0.25 mm土壤团聚体比例计算公式如下[23]:



式中,MWD为平均重量直径,GMD为平均几何直径,R0.25为>0.25 mm土壤团聚体所占比例,M>0.25为>0.25 mm土壤团聚体质量,Xi为第i级团聚体的质量,Mi为第i级团聚体平均直径,其中>2 mm、2—0.25 mm、0.25—0.053 mm和<0.053 mm粒级的团聚体对应的平均直径分别为2、1.125、0.1515和0.053 mm。
土壤团聚体有机碳贡献率(WSOC)和全氮贡献率(WTN)计算公式如下:


式中,OCi为第i级团聚体有机碳含量,Ni为第i级团聚体全氮含量,Mi为第i级团聚体所占比例,SOC为土壤有机碳含量。TN为土壤全氮含量。
1.3.5 统计分析 采用SAS 9.2软件进行数据统计分析,各处理不同粒级土壤团聚体比例,团聚体平均重量直径、平均几何直径、团聚体有机碳含量、全氮含量,贡献率等均采用SAS软件中的PROC GLM过程进行方差分析,其中处理为固定因子,区组为随机因子,采用最小显著极差法在0.05水平上进行多重比较,采用Excel 2016软件作图。
2 结果
2.1 不同多样化种植模式下土壤团聚体分级
由图1可以看出,在0—20 cm土层,M-S和M/P处理>2 mm土壤水稳性团聚体比例分别比CM处理高8.6%和10.7%。M/S处理0.25—2 mm土壤水稳性团聚体比例则比CM处理高15.3%。各处理0.053— 0.25 mm土壤水稳性团聚体间无显著差异。M/S和M-S处理<0.053 mm土壤水稳性团聚体比例比CM处理低15.0%和15.7%。在20—40 cm土层,M-S处理>2 mm土壤水稳性团聚体比例比CM高7.7%,而其他处理>2 mm土壤水稳性团聚体比例则与CM处理无显著差异。M/S和M-P处理0.25—2 mm土壤水稳性团聚体比例分别比CM处理高5.3%和8.2%。M/S和M-P处理0.053—0.25 mm土壤水稳性团聚体比例则比CM处理低5.0%和6.6%。M/S处理<0.053 mm土壤水稳性团聚体比例则比CM处理低5.4%,而其他处理<0.053 mm土壤水稳性团聚体比例则与CM处理无显著差异。
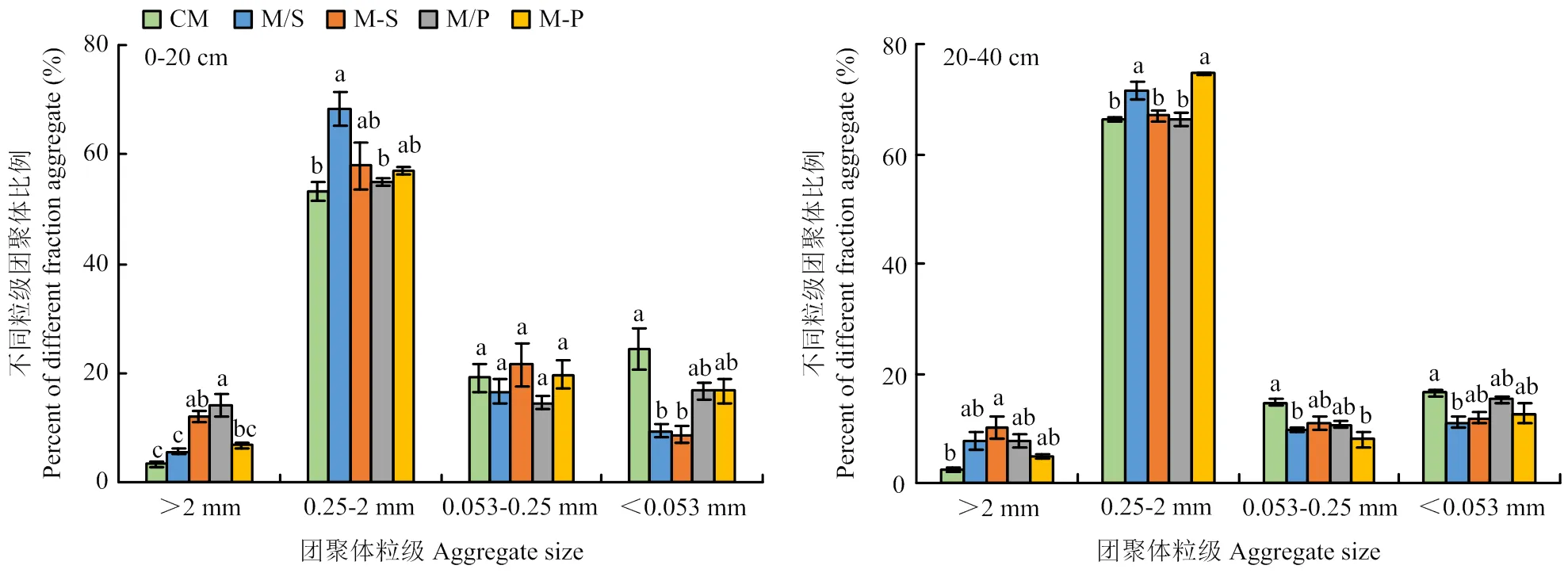
CM:玉米连作;M/S:玉米-大豆间作;M-S:玉米-大豆轮作;M/P:玉米-花生间作;M-P:玉米-花生轮作。不同小写字母表示处理间差异显著(P<0.05)。下同
2.2 不同多样化种植模式下土壤团聚体平均重量直径和平均几何直径
由图2可以看出,在0—20 cm土层,M/S、M-S和M/P处理土壤团聚体平均重量直径分别比CM处理高29.1%、31.2%和31.4%,而在20—40 cm土层,M/S、M-S和M-P处理土壤团聚体平均重量直径分别比CM处理高18.4%、18.5%和15.3%。
在0—20 cm土层,M/S和M-S处理土壤团聚体平均几何直径分别比CM处理高68.7%和62.2%,而M/P和M-P处理土壤团聚体平均几何直径则与CM处理无显著差异。在20—40 cm土层,M/S、M-S和M-P处理土壤团聚体平均几何直径分别比CM处理高34.4%、30.2%和30.3%,而M/P处理土壤团聚体平均几何直径则与CM处理无显著差异。

图2 土壤团聚体平均重量直径和几何直径
由图3可以看出,在0—20 cm土层,M/S和M-S处理>0.25 mm土壤团聚体比例分别比CM处理高17.6%和13.4%,而M/P和M-P处理>0.25 mm土壤团聚体比例则与CM处理无显著差异。在20—40 cm土层,M/S、M-S和M-P处理>0.25 mm土壤团聚体比例分别比CM处理高10.4%、8.3%和10.5%,而M/P处理>0.25 mm土壤团聚体比例则与CM处理无显著差异。

图3 >0.25 mm土壤水稳性团聚体比例
2.3 不同多样化种植模式下土壤团聚体有机碳含量
由图4可知,在0—20 cm土层,M/S、M-S、M/P和M-P处理>2 mm土壤团聚体有机碳含量分别比CM处理高20.7%、24.3%、18.8%和17.8%。M-S、M/P和M-P处理0.25—2 mm土壤团聚体有机碳含量分别比CM处理高13.3%、17.8%和15.0%。各处理0.053—0.25 mm和<0.053 mm土壤团聚体有机碳含量则无显著差异。在20—40 cm土层,M-S处理>2 mm土壤团聚体有机碳含量比CM处理高11.6%,其他处理>2 mm土壤团聚体有机碳含量则与CM处理无显著差异。仅M/P处理0.25—2 mm土壤团聚体有机碳含量比CM处理高23.6%,而其他处理0.25—2 mm土壤团聚体有机碳含量则与CM处理无显著差异。各处理0.053—0.25 mm土壤团聚体有机碳含量与CM均无显著差异。M-S处理<0.053 mm土壤团聚体有机碳含量比CM处理高18.6%,其他处理<0.053 mm土壤团聚体有机碳含量则与CM处理无显著差异。
2.4 不同多样化种植模式下土壤团聚体全氮含量
图5显示,在0—20 cm土层,M-S、M/P和M-P处理>2 mm土壤团聚体全氮含量分别比CM处理高13.0%、16.8%和14.8%,而M/S处理>2 mm土壤团聚体全氮含量则与CM处理无显著差异。各处理0.25—2 mm土壤团聚体全氮含量与CM处理无显著差异。M-P的处理0.053— 0.25 mm和<0.053 mm土壤团聚体全氮含量分别比CM处理高18.9%和10.3%,而M/S、M-S和M/P处理0.053— 0.25 mm和<0.053 mm土壤团聚体全氮含量则与CM处理均无显著差异。在20—40 cm土层,M-S、M/P和M-P处理>2 mm土壤团聚体全氮含量分别比CM处理高13.7%、14.5%和16.7%。M-S、M/P和M-P处理0.25—2 mm土壤团聚体全氮含量则分别比CM处理高23.9%、26.9%和17.2%,而M/S处理>2 mm和0.25—2 mm土壤团聚体全氮含量与CM处理无显著差异。M/S、M-S、M/P和M-P处理0.053—0.25 mm和<0.053 mm土壤团聚体全氮含量均与CM处理无显著差异。

图4 不同粒级土壤团聚体有机碳含量

图5 不同粒级土壤团聚体全氮含量
2.5 多样化种植模式下不同粒级团聚体有机碳对土壤总有机碳的贡献率
表1显示,在0—20 cm土层,M-S和M/P处理>2 mm土壤团聚体有机碳对总有机碳贡献率分别比CM处理高10.0和11.2个百分点,而M/S和M-P处理>2 mm土壤团聚体有机碳对总有机碳贡献率则与CM处理无显著差异。M/S处理0.25—2 mm土壤团聚体有机碳对总有机碳贡献率比CM处理高15.4个百分点,而M/S、M-S和M-P处理>2 mm土壤团聚体有机碳对总有机碳贡献率则与CM处理无显著差异。M/S、M-S、M/P和M-P处理0.053—0.25 mm土壤团聚体有机碳对总有机碳的贡献率与CM处理无显著差异。M/S和M-S处理<0.053 mm土壤团聚体有机碳对总有机碳的贡献率比CM处理低16.2和17.8个百分点,而M/P和M-P处理<0.053 mm土壤团聚体有机碳对总有机碳的贡献率则与CM处理无显著差异。
在20—40 cm土层,M-S处理>2 mm土壤团聚体有机碳对总有机碳贡献率比CM处理高8.3个百分点,而M/S、M/P和M-P处理>2 mm土壤团聚体有机碳对总有机碳贡献率则与CM处理无显著差异。M-P处理0.25—2 mm土壤团聚体有机碳对总有机碳贡献率比CM处理高7.6个百分点,M/S、M-S和M/P处理0.25—2 mm土壤团聚体有机碳对总有机碳贡献率与CM处理无显著差异。M/S、M/P和M-P处理0.053—0.25 mm土壤团聚体有机碳对总有机碳贡献率分别比CM处理低3.9、4.2和5.6个百分点,而M-S处理0.053—0.25 mm土壤团聚体有机碳对总有机碳贡献率则与CM处理无显著差异。各处理<0.053 mm土壤团聚体有机碳对总有机碳贡献率与CM处理均无显著差异。
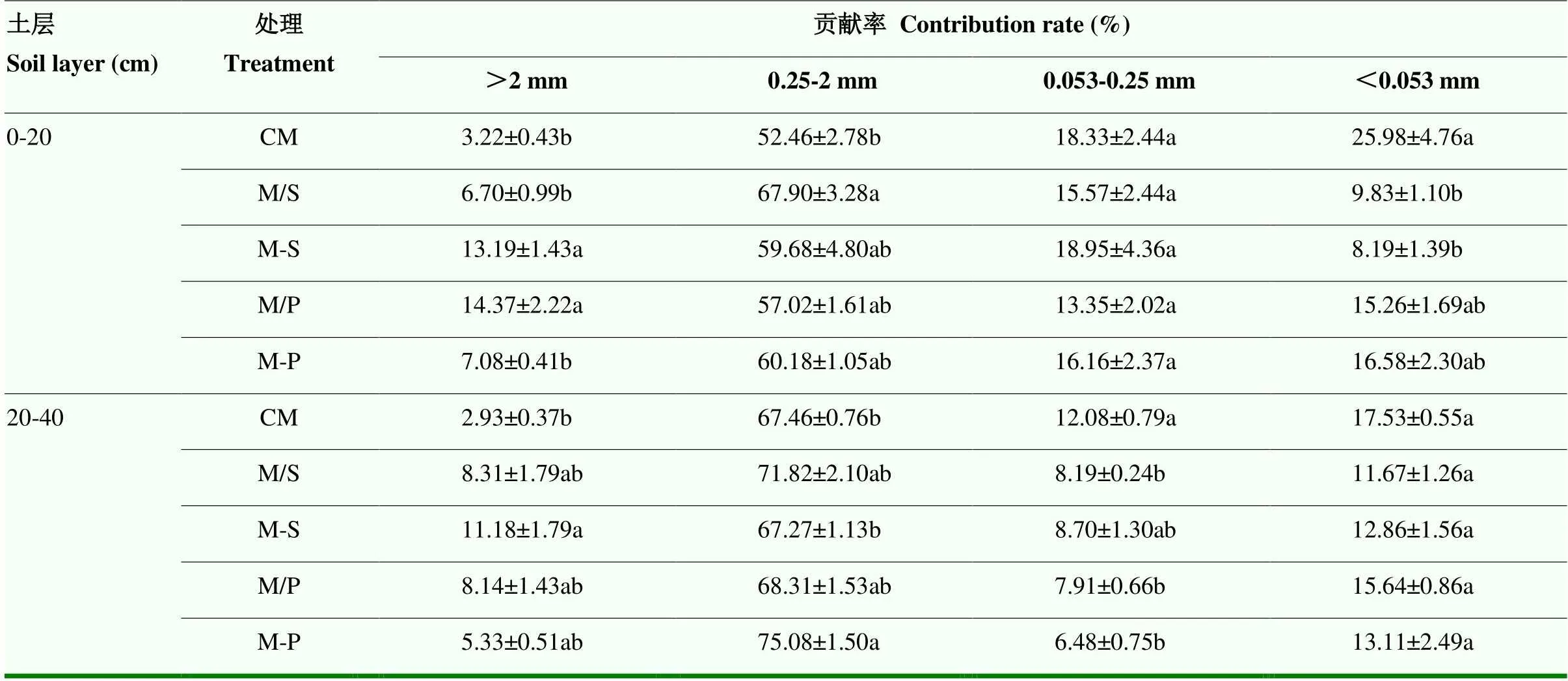
表1 不同粒级团聚体有机碳对土壤总有机碳的贡献率
2.6 多样化种植模式下不同粒级团聚体全氮对土壤全氮的贡献率
表2显示,在0—20 cm土层,M-S和M/P处理>2 mm团聚体全氮含量对土壤全氮的贡献率分别比CM处理高9.2和12.4个百分点,而M/S和M-P处理>2 mm团聚体全氮含量对土壤全氮的贡献率则与CM处理无显著差异。M/S处理0.25—2 mm团聚体全氮含量对土壤全氮的贡献率比CM处理高16.5个百分点,M-S、M/P和M-P处理0.25—2 mm团聚体全氮含量对土壤全氮的贡献率则与CM处理无差异。各处理0.053—0.25 mm团聚体全氮含量对土壤全氮的贡献率均无差异。M/S和M-S处理<0.053 mm团聚体全氮含量对土壤全氮的贡献分别比CM处理低16.1和17.1个百分点,而M/P和M-P处理<0.053 mm团聚体全氮含量对土壤全氮的贡献则与CM处理无显著差异。在20—40 cm土层,M-S处理>2 mm团聚体全氮含量对土壤全氮的贡献率比CM处理高7.1个百分点,而M/S、M/P和M-P处理>2 mm团聚体全氮含量对土壤全氮的贡献率则与CM处理无显著差异。M/S和M-P处理0.25—2 mm团聚体全氮含量对土壤全氮的贡献率分别比CM处理高7.1和8.9个百分点,M-S和M/P处理0.25—2 mm团聚体全氮含量对土壤全氮的贡献率则与CM处理无显著差异。M/S、M-S、M/P和M-P处理0.053—0.25 mm团聚体全氮含量对土壤全氮的贡献率分别比CM处理低6.2、4.4、5.9和6.7个百分点。各处理<0.053 mm团聚体全氮含量对土壤全氮的贡献率则无显著差异。
3 讨论
3.1 作物多样化种植促进了土壤大团聚体的形成
土壤团聚体是土壤结构的基本单元,是土壤微生物的生境及养分储存场所[24]。土壤团聚体含量及粒级分布不仅影响作物生长发育,还对土壤的一系列物理、化学及生物学过程有着重要影响。土壤团聚体结构稳定性可以用土壤团聚体平均重量直径和平均几何直径来表征,土壤团聚体平均重量直径和几何直径越高,说明团聚体稳定性越强[25]。本研究表明,与玉米连作相比,玉米与豆科作物间作、轮作可以增加土壤水稳性大团聚体(>0.25 mm)含量,降低黏粉粒(<0.053 mm)含量,玉米与豆科作物间作和轮作均可增加土壤团聚体平均重量直径和平均几何直径,土壤团聚体结构稳定性高于玉米连作。乔鑫鑫等[26]在华北平原的研究表明,冬小麦–夏玉米||花生复种模式较传统冬小麦–夏玉米复种模式明显增加了土壤大团聚体含量,增强了团聚体的机械稳定性和水稳定性。白录顺等[27]在南方红壤开展的间作模式试验结果表明,玉米/大豆模式可以显著提高水稳性大团聚体(1—5 mm)含量,其中对>2 mm团聚体提高效果最明显,玉米/大豆模式下各土层团聚体平均重量直径和平均几何直径显著高于单作。魏艳春等[28]在黄土高原的研究结果表明,玉米与豆科作物轮作可明显提高土壤水稳性大团聚体含量。LI等[11]通过在甘肃和宁夏设置的间作模式长期定位试验表明,不同作物间作可以增加土壤大团聚体含量,提高团聚体稳定性,这与本研究的结果均一致。分析原因,一方面是由于多样化种植模式根系分泌物种类和数量增加,根际真菌和菌丝活力增强,促进了菌丝的生长和多糖类物质的分泌,增强了微团聚体的胶结作用,从而促进微团聚体转化为大团聚体,增强了土壤团聚体的稳定性[29-30]。另一方面,不同作物的交替更换有利于维持土壤生物多样性,土壤碳氮转化作用增强,产生了大量蛋白质、多糖、木质素等物质,从而间接促进了大团聚体的形成[30-32]。此外,多样化种植(间作和轮作)模式通过间接影响土壤微生物群落组成来增强土壤的聚集性,如提高土壤和丛枝菌真菌(AMF)的相对丰度,或降低的相对丰度[33],从而间接促进土壤团聚体的形成[34]。

表2 不同粒级团聚体全氮对土壤全氮的贡献率
3.2 作物多样化种植可以提高土壤团聚体有机碳氮含量及其贡献率
土壤有机碳是土壤团聚体重要的胶结物质,对土壤团聚体的形成发挥重要作用,而土壤团聚体作为土壤有机碳的重要储存场所,同样对土壤有机碳起到了保护作用[35]。本研究发现,玉米与豆科作物间作、轮作增加了土壤团聚体有机碳和全氮含量。这与前人的研究结果一致[26,28,36]。乔鑫鑫等[26]在华北平原的研究表明,冬小麦–夏玉米||花生复种模式较传统冬小麦–夏玉米复种模式可显著提高土壤团聚体(特别是>0.25 mm粒级团聚体)的碳、氮含量。魏艳春等[28]在黄土高原的研究结果表明,玉米与豆科作物轮作可明显提高土壤团聚体有机碳和全氮含量。李孝梅等[36]研究发现,玉米大豆间作和玉米萝卜间作可以增加土壤团聚体有机碳、氮含量。分析原因,一方面是由于大豆和花生均为豆科作物,具有较强的固氮功能,可以将大气中的氮固定储存到土壤中,不仅可以增加土壤氮含量,同时增加的土壤氮素可以促进作物生长和对碳的固定,从而可以间接增加土壤有机碳含量[37];另一方面,由于间作和轮作促进了作物多样性的增加,而作物多样性的增加可导致土壤微生物活性和多样性增加,从而对土壤碳、氮循环等产生正向影响[38-40]。此外,多样化种植特别是含有豆科作物的地块,由于豆科作物碳氮比较低导致秸秆分解速率较高,较高的固氮能力、较多的残茬输入改善了土壤结构、养分供应和土壤生物功能,增强了生物-非生物相互作用,从而有利于土壤碳氮的积累[41-42]。本研究发现,玉米与豆科作物间作和轮作模式主要是增加了>2 mm和0.25—2 mm土壤团聚体有机碳和全氮含量,这与SIX等[19]提出的“大团聚体周转”概念模型相符,即新输入有机物在胶结物质作用下首先形成大团聚体,土壤有机碳、氮等养分首先在大团聚体中聚集,然后随着颗粒有机物的分解,微团聚体逐渐形成,微团聚体中土壤有机碳、氮养分含量增加,由于大团聚体向微团聚体转化需要较长时间,因此,多样化种植模式对土壤团聚体碳氮含量的影响还需要较长的时间来验证。
3.3 土壤团聚体有机碳氮含量与作物产量的关系
土壤有机碳是评价土壤肥力的重要指标,在一定范围内,土壤有机碳的数量和固存状态与作物生产力存在显著相关性[43]。前人研究表明,作物产量随土壤水稳性团聚体及粉黏粒组分中有机碳含量的增加而呈线性增加的趋势[44]。本研究发现多样化种植模式增加了土壤团聚体稳定性,进而促进了有机碳和全氮含量的增加。本试验的前期结果也表明,多样化种植同样有增加作物产量的趋势[45]。分析其增产机制可能是多样化种植通过增加根系分泌物,促进了土壤团聚体胶结,增加土壤团聚体稳定性,有利于土壤团聚体有机碳和全氮的固持[46-47]。而作物多样化种植下土壤微生物活动显著增加,促进了土壤团聚体养分的储存和释放[40],增加了养分利用率,从而提高作物产量。
本研究重点分析了短期多样化种植对土壤团聚体及碳氮的影响,其长期效应仍有待长期监测与深入分析,此外,多样化种植模式下土壤团聚体碳氮特征与其他土壤指标及作物生产指标间的关系需要进一步探究。
4 结论
东北黑土区短期禾本科与豆科作物多样化种植有增加土壤水稳性大团聚体含量,降低黏粉粒含量的趋势,>0.25 mm土壤水稳性团聚体含量显著高于玉米连作。多样化种植可以增加土壤团聚体稳定性,其中玉米与大豆间作、轮作模式0—20和20—40 cm土层及玉米与花生轮作20—40 cm土层土壤团聚体平均重量直径和几何直径均显著高于玉米连作。多样化种植显著增加了土壤有机碳和全氮含量,主要得益于>2 mm土壤水稳性团聚体有机碳和全氮对土壤总有机碳和总氮的贡献率增加。
[1] TILMAN D, BALZER C, HILL J, BEFORT B L. Global food demand and the sustainable intensification of agriculture. PNAS, 2011, 108(50): 20260-20264.
[2] MESSÉAN A, VIGUIER L, PARESYS L, AUBERTOT J N, CANALI S, IANNETTA P, JUSTES E, KARLEY A, KEILLOR B, KEMPER L, MUEL F, PANCINO B, STILMANT D, WATSON C, WILLER H, ZORNOZA R. Enabling crop diversification to support transitions toward more sustainable European agrifood systems. Frontiers of Agricultural Science and Engineering, 2021: 8(3): 474-480.
[3] ZHAO J, YANG Y D, ZHANG K, JEONG J, ZENG Z H, ZANG H D. Does crop rotation yield more in China? A meta-analysis. Field Crops Research, 2020, 245: 107659.
[4] RENARD D, TILMAN D. National food production stabilized by crop diversity. Nature, 2019, 571(7764): 257-260.
[5] CONG W F, HOFFLAND E, LI L, SIX J, SUN J H, BAO X G, ZHANG F S, VAN DER WERF W. Intercropping enhances soil carbon and nitrogen. Global Change Biology, 2015, 21(4): 1715-1726.
[6] OELBERMANN M, ECHARTE L. Evaluating soil carbon and nitrogen dynamics in recently established maize-soybean inter- cropping systems. European Journal of Soil Science, 2011, 62(1): 35-41.
[7] LIU X, RAHMAN T, SONG C, YANG F, SU B Y, CUI L, BU W Z, YANG W Y. Relationships among light distribution, radiation use efficiency and land equivalent ratio in maize-soybean strip intercropping. Field Crops Research, 2018, 224: 91-101.
[8] 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响. 作物学报, 2022, 48(6): 1476-1487.
YANG H, ZHOU Y, CHEN P, DU Q, ZHENG B C, PU T, WEN J, YANG W Y, YONG T W. Effects of nutrient uptake and utilization on yield of maize-legume strip intercropping system. Acta Agronomica Sinica, 2022, 48(6): 1476-1487. (in Chinese)
[9] VIRK A L, LIN B J, KAN Z R, QI J Y, DANG Y P, LAL R, ZHAO X, ZHANG H L. Simultaneous effects of legume cultivation on carbon and nitrogen accumulation in soil. Advances in Agronomy. Amsterdam: Elsevier, 2022, 171: 75-110.
[10] MIKHA M M, RICE C W. Tillage and manure effects on soil and aggregate-associated carbon and nitrogen. Soil Science Society of America Journal, 2004, 68(3): 809-816.
[11] LI X F, WANG Z G, BAO X G, SUN J H, YANG S C, WANG P, WANG C B, WU J P, LIU X R, TIAN X L, WANG Y, LI J P, WANG Y, XIA H Y, MEI P P, WANG X F, ZHAO J H, YU R P, ZHANG W P, CHE Z X, GUI L G, CALLAWAY R M, TILMAN D, LI L. Long-term increased grain yield and soil fertility from intercropping. Nature Sustainability, 2021, 4(11): 943-950.
[12] DIJKSTRA F A, HOBBIE S E, REICH P B, KNOPS J M H. Divergent effects of elevated CO2, N fertilization, and plant diversity on soil C and N dynamics in a grassland field experiment. Plant and Soil, 2005, 272(1): 41-52.
[13] BRONICK C J, LAL R. Soil structure and management: A review. Geoderma, 2005, 124(1/2): 3-22.
[14] GUNINA A, KUZYAKOV Y. Pathways of litter C by formation of aggregates and SOM density fractions: Implications from13C natural abundance. Soil Biology and Biochemistry, 2014, 71: 95-104.
[15] XIE J, PENG B, WANG R, BATBAYAR J, HOOGMOED M, YANG Y, ZHANG S, YANG X, SUN B. Responses of crop productivity and physical protection of organic carbon by macroaggregates to long-term fertilization of an Anthrosol. European Journal of Soil Science, 2018, 69(3): 555-567.
[16] NYAWADE S O, KARANJA N N, GACHENE C K K, GITARI H I, SCHULTE-GELDERMANN E, PARKER M L. Short-term dynamics of soil organic matter fractions and microbial activity in smallholder potato-legume intercropping systems. Applied Soil Ecology, 2019, 142: 123-135.
[17] BIMÜLLER C, KREYLING O, KÖLBL A, VON LÜTZOW M, KÖGEL-KNABNER I. Carbon and nitrogen mineralization in hierarchically structured aggregates of different size. Soil and Tillage Research, 2016, 160: 23-33.
[18] 张维理, KOLBE H, 张认连. 土壤有机碳作用及转化机制研究进展.中国农业科学, 2020, 53(2): 317-331. doi: 10.3864/j.issn.0578-1752. 2020.02.007.
ZHANG W L, KOLBE H, ZHANG R L. Research progress of SOC functions and transformation mechanisms. Scientia Agricultura Sinica, 2020, 53(2): 317-331. doi: 10.3864/j.issn.0578-1752.2020.02.007. (in Chinese)
[19] SIX J, ELLIOTT E T, PAUSTIAN K. Soil macroaggregate turnover and microaggregate formation: a mechanism for C sequestration under no-tillage agriculture. Soil Biology and Biochemistry, 2000, 32(14): 2099-2103.
[20] 赵兰坡, 王鸿斌, 刘会青, 王艳玲, 刘淑霞, 王宇. 松辽平原玉米带黑土肥力退化机理研究. 土壤学报, 2006, 43(1): 79-84.
ZHAO L P, WANG H B, LIU H Q, WANG Y L, LIU S X, WANG Y. Mechanism of fertility degradation of black soil in corn belt of Songliao plain. Acta Pedologica Sinica, 2006, 43(1): 79-84. (in Chinese)
[21] 于磊, 张柏. 中国黑土退化现状与防治对策. 干旱区资源与环境, 2004, 18(1): 99-103.
YU L, ZHANG B. The degradation situations of black soil in China and its prevention and counter measures. Journal of Arid Land Resources & Environment, 2004, 18(1): 99-103. (in Chinese)
[22] 鲍士旦. 土壤农化分析. 3版. 北京: 中国农业出版社, 2000.
BAO S D. Soil and Agricultural Chemistry Analysis. 3rd ed. Beijing: China Agriculture Press, 2000. (in Chinese)
[23] KEMPER W D, ROSENAU R C. Aggregate stability and size distribution//KLUTE A, CAMPBELL G S, JACKSON R D. Methods of Soil Analysis. Part 1. Physical and Mineralogical Methods. Madison: WI, 1986: 425-442.
[24] WANG F, ZHANG X X, NEAL A L, CRAWFORD J W, MOONEY S J, BACQ-LABREUIL A. Evolution of the transport properties of soil aggregates and their relationship with soil organic carbon following land use changes. Soil and Tillage Research, 2022, 215: 105226.
[25] LIU Z X, CHEN X M, JING Y, LI Q X, ZHANG J B, HUANG Q R. Effects of biochar amendment on rapeseed and sweet potato yields and water stable aggregate in upland red soil. Catena, 2014, 123: 45-51.
[26] 乔鑫鑫, 王艳芳, 李乾云, 包全发, 尹飞, 焦念元, 付国占, 刘领. 复种模式对豫西褐土团聚体稳定性及其碳、氮分布的影响. 植物营养与肥料学报, 2021, 27(3): 380-391.
QIAO X X, WANG Y F, LI Q Y, BAO Q F, YIN F, JIAO N Y, FU G Z, LIU L. Effects of multi-cropping systems on cinnamon soil aggregate stability, carbon and nitrogen distribution in western Henan Province. Journal of Plant Nutrition and Fertilizers, 2021, 27(3): 380-391. (in Chinese)
[27] 白录顺, 范茂攀, 王自林, 王婷, 邓超, 李永梅. 间作模式下玉米/大豆的根系特征及其与团聚体稳定性的关系. 水土保持研究, 2019, 26(1): 124-129.
BAI L S, FAN M P, WANG Z L, WANG T, DENG C, LI Y M. Relationship between root characteristics and aggregate stability in the field with maize and soybean intercropping. Research of Soil and Water Conservation, 2019, 26(1): 124-129. (in Chinese)
[28] 魏艳春, 马天娥, 魏孝荣, 王昌钊, 郝明德, 张萌. 黄土高原旱地不同种植系统对土壤水稳性团聚体及碳氮分布的影响. 农业环境科学学报, 2016, 35(2): 305-313.
WEI Y C, MA T E, WEI X R, WANG C Z, HAO M D, ZHANG M. Effects of cropping systems on distribution of water-stable aggregates and organic carbon and nitrogen in soils in semiarid farmland of the Loess Plateau. Journal of Agro-Environment Science, 2016, 35(2): 305-313. (in Chinese)
[29] 王婷, 李永梅, 王自林, 肖靖秀, 白录顺, 范茂攀. 间作对玉米根系分泌物及团聚体稳定性的影响. 水土保持学报, 2018, 32(3): 185-190.
WANG T, LI Y M, WANG Z L, XIAO J X, BAI L S, FAN M P. Effects of intercropping on maize root exudates and soil aggregate stability. Journal of Soil and Water Conservation, 2018, 32(3): 185-190. (in Chinese)
[30] 向蕊, 伊文博, 赵薇, 王顶, 赵平, 龙光强, 汤利. 间作对土壤团聚体有机碳储量的影响及其氮调控效应. 水土保持学报, 2019, 33(5): 303-308.
XIANG R, YI W B, ZHAO W, WANG D, ZHAO P, LONG G Q, TANG L. Effects of intercropping on soil aggregate-associated organic carbon storage and nitrogen regulation. Journal of Soil and Water Conservation, 2019, 33(5): 303-308. (in Chinese)
[31] FREIXO A A, DE A MACHADO P L O, DOS SANTOS H P, SILVA C A, DE S FADIGAS F. Soil organic carbon and fractions of a Rhodic Ferralsol under the influence of tillage and crop rotation systems in southern Brazil. Soil and Tillage Research, 2002, 64(3/4): 221-230.
[32] WEI X R, SHAO M G, GALE W J, ZHANG X C, LI L H. Dynamics of aggregate-associated organic carbon following conversion of forest to cropland. Soil Biology and Biochemistry, 2013, 57: 876-883.
[33] TIAN X L, WANG C B, BAO X G, WANG P, LI X F, YANG S C, DING G C, CHRISTIE P, LI L. Crop diversity facilitates soil aggregation in relation to soil microbial community composition driven by intercropping. Plant and Soil, 2019, 436(1): 173-192.
[34] GUPTA V V S R, GERMIDA J J. Distribution of microbial biomass and its activity in different soil aggregate size classes as affected by cultivation. Soil Biology and Biochemistry, 1988, 20(6): 777-786.
[35] 邱晓蕾, 宗良纲, 刘一凡, 杜霞飞, 罗敏, 汪润池. 不同种植模式对土壤团聚体及有机碳组分的影响. 环境科学, 2015, 36(3): 1045-1052.
QIU X L, ZONG L G, LIU Y F, DU X F, LUO M, WANG R C. Effects of different cultivation patterns on soil aggregates and organic carbon fractions. Environmental Science, 2015, 36(3): 1045-1052. (in Chinese)
[36] 李孝梅, 李永梅, 乌达木, 范茂攀. 玉米间作大豆、萝卜对红壤不同粒径水稳性团聚体碳氮分布的影响. 中国土壤与肥料, 2022(1): 104-111.
LI X M, LI Y M, WU D M, FAN M P. Effects of maize//soybean and maize//radish inter-cropping on the carbon and nitrogen distribution of water-stable aggregates in red soil. Soil and Fertilizer Sciences in China, 2022(1): 104-111. (in Chinese)
[37] QUINN THOMAS R, CANHAM C D, WEATHERS K C, GOODALE C L. Increased tree carbon storage in response to nitrogen deposition in the US. Nature Geoscience, 2010, 3(1): 13-17.
[38] JIA Y F, ZHAI G Q, ZHU S S, LIU X J, SCHMID B, WANG Z H, MA K P, FENG X J. Plant and microbial pathways driving plant diversity effects on soil carbon accumulation in subtropical forest. Soil Biology and Biochemistry, 2021, 161: 108375.
[39] LANGE M, EISENHAUER N, SIERRA C A, BESSLER H, ENGELS C, GRIFFITHS R I, MELLADO-VÁZQUEZ P G, MALIK A A, ROY J, SCHEU S, STEINBEISS S, THOMSON B C, TRUMBORE S E, GLEIXNER G. Plant diversity increases soil microbial activity and soil carbon storage. Nature Communications, 2015, 6: 6707.
[40] MCDANIEL M D, TIEMANN L K, GRANDY A S. Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecological Applications, 2014, 24(3): 560-570.
[41] PROMMER J, WALKER T W N, WANEK W, BRAUN J, ZEZULA D, HU Y T, HOFHANSL F, RICHTER A. Increased microbial growth, biomass, and turnover drive soil organic carbon accumulation at higher plant diversity. Global Change Biology, 2020, 26(2): 669-681.
[42] ZHANG K L, MALTAIS-LANDRY G, LIAO H L. How soil biota regulate C cycling and soil C pools in diversified crop rotations. Soil Biology and Biochemistry, 2021, 156: 108219.
[43] LAL R. Beyond Copenhagen: mitigating climate change and achieving food security through soil carbon sequestration. Food Security, 2010, 2(2): 169-177.
[44] 曹寒冰, 谢钧宇, 强久次仁, 郭璐, 洪坚平, 荆耀栋, 孟会生. 施肥措施对复垦土壤团聚体碳氮含量和作物产量的影响. 农业工程学报, 2020, 36(18): 135-143.
CAO H B, XIE J Y, QIANGJIU C R, GUO L, HONG J P, JING Y D, MENG H S. Effects of fertilization regimes on carbon and nitrogen contents of aggregates and maize yield in reclaimed soils. Transactions of the Chinese Society of Agricultural Engineering, 2020, 36(18): 135-143. (in Chinese)
[45] SUN T, FENG X M, LAL R, CAO T H, GUO J R, DENG A X, ZHENG C Y, ZHANG J, SONG Z W, ZHANG W J. Crop diversification practice faces a tradeoff between increasing productivity and reducing carbon footprints. Agriculture, Ecosystems & Environment, 2021, 321: 107614.
[46] JOSEPH U E, TOLUWASE A O, KEHINDE E O, OMASAN E E, TOLULOPE A Y, GEORGE O O, ZHAO C S, WANG H Y. Effect of biochar on soil structure and storage of soil organic carbon and nitrogen in the aggregate fractions of an Albic soil. Archives of Agronomy and Soil Science, 2020, 66(1): 1-12.
[47] 张贺, 杨静, 周吉祥, 李桂花, 张建峰. 连续施用土壤改良剂对砂质潮土团聚体及作物产量的影响. 植物营养与肥料学报, 2021, 27(5): 791-801.
ZHANG H, YANG J, ZHOU J X, LI G H, ZHANG J F. Effects of organic and inorganic amendments on aggregation and crop yields in sandy fluvo-aquic soil. Journal of Plant Nutrition and Fertilizers, 2021, 27(5): 791-801. (in Chinese)
Effects of Diversified Cropping on the Soil Aggregate Composition and Organic Carbon and Total Nitrogen Content
SUN Tao1,3, FENG XiaoMin2,3, GAO XinHao1, DENG AiXing3, ZHENG ChengYan3, SONG ZhenWei3, ZHANG WeiJian3
1State Key Laboratory of Nutrient Use and Management/Key Laboratory of Wastes Matrix Utilization, Ministry of Agriculture and Rural Affairs/Institute of Agricultural Resources and Environment, Shandong Academy of Agricultural Sciences, Ji’nan 250100;2Institute of Sorghum Research, Shanxi Agricultural University, Jinzhong 030600, Shanxi;3Institute of Crop Sciences, Chinese Academy of Agricultural Sciences/Key Laboratory of Crop Physiology and Ecology, Ministry of Agriculture and Rural Affairs, Beijing 100081
【Objective】In this study, the composition as well as carbon and nitrogen distribution characteristics of soil aggregates under diversified cropping system of poaceae and legumes in the black soil region of Northeast China were elucidated, which could provide the theoretical basis and technical guidance for promoting the optimization of cropping system of combined use and cultivation in black soil. 【Method】Field experiment was conducted from 2016 to 2020 in Gongzhuling Experimental Station, Institute of Crop Sciences, Chinese Academy of Agricultural Sciences. Four diversified cropping systems were considered, including maize-soybean intercropping (M/S), maize-soybean rotation (M-S), maize-peanut intercropping (M/P), and maize-peanut rotation (M-P), while the maize continuous cropping (CM) system was used as control. Soil samples of 0-20 cm and 20-40 cm layers were collected after harvesting in October 2020. Then, soil aggregates and their soil organic carbon (SOC) and total nitrogen (TN) content under diversified cropping system were analyzed. 【Result】Diversified cropping was beneficial to increase the content of water-stable macro-aggregates in 0-20 cm and 20-40 cm soil (>0.25 mm), and to reduce the content of silt and clay (<0.053 mm), the mean weight diameter (MWD) and geometric mean diameter (GMD) ofsoil aggregates under four diversified cropping systems were significantly higher than those of maize continuous cropping system (<0.05). Soil aggregates ratios of >0.25 mm under M/S and M-S treatments were 17.5% and 13.4% higher than that under CM treatment in 0-20 cm soil layer, respectively. Soil aggregates ratios of >0.25 mm under M/S, M-S and M-P treatments was 10.4%, 8.3% and 10.5% higher than that under CM treatment in the 20-40 cm soil layer, respectively. Diversified cropping increased the SOC and TN content of soil aggregates. In 0-20 cm soil layer, the SOC of >2 mm soil aggregate under M/S, M-S, M/P and M-P treatments was 20.7%, 24.3%, 18.8% and 17.8% higher than that under CM treatment, respectively; the TN of >2 mm soil aggregate under M-S, M/P and M-P treatments was 13.0%, 16.8% and 14.8% higher than that under CM treatment, respectively. When compared with CM treatment, the contribution rates of >2 mm soil aggregate to SOC and TN under M-S and M/P treatments were higher in 0-20 cm soil layer, while the contribution rate of <0.053 mm soil aggregate to SOC and TN under M/S and M-S treatments were lower in 0-20 cm soil layer. 【Conclusion】Diversified cropping increased the content of soil macroaggregates, decreased the content of clay particles, improved the soil aggregate stability, SOC and TN of soil aggregates, which was conducive to promoting the SOC and TN sequestration in black soil region of Northeast China.
black soil region of Northeast China; rotation; intercropping; maize; soybean; peanut; soil aggregate; soil organic carbon; soil total nitrogen

10.3864/j.issn.0578-1752.2023.15.008
2022-08-13;
2022-12-06
中国农业科学院科技创新工程联合攻关重大科研任务(CAAS-ZDRW202202)、国家绿肥产业技术体系(CARS-22-G-16)、国家自然科学基金(31671642)、山东省农业科学院农业科技创新工程(CXGC2022E03)
孙涛,E-mail:suntao1007@126.com。通信作者宋振伟,E-mail:songzhenwei@caas.cn
(责任编辑 李云霞)
