玉米-大刍草渗入系群体的剑叶遗传基础解析
2023-08-07高沐甜邱冠杰朱通通李瑞莲邓敏罗红兵黄成
高沐甜 邱冠杰 朱通通 李瑞莲,2,3 邓敏,2,3 罗红兵,2,3 黄成,2,3
(1 湖南农业大学农学院,410128,湖南长沙;2 作物生理与分子生物学教育部重点实验室,410128,湖南长沙;3 湖南省玉米工程技术研究中心,410128,湖南长沙)
玉米剑叶是玉米雌穗苞叶顶端的进一步延伸,是玉米长期进化过程中退化的一种器官,是鉴别玉米品种和种质来源的重要性状之一[1]。不同种质来源玉米的剑叶长度差异较大,大部分温带甜玉米具有较长的剑叶,而大部分热带玉米则没有剑叶或剑叶较短[2]。剑叶作为玉米雌穗的重要组成部分,对玉米雌穗的生长发育和产量形成都具有重要作用。Cantrell 等[3]在3 个环境下利用6 个具有长剑叶的不同马齿型玉米材料探究了剑叶与产量之间的相互关系,发现6 个材料中有4 个在完全去除剑叶的条件下显著降低了玉米产量,平均降幅为2.6%。Sawada 等[4]进一步利用水培和大田试验探究了剑叶和茎生叶在光合速率和光合产物转运等方面的特征,发现剑叶与茎生叶具有大致相当的光合速率,但剑叶的光合产物转运效率(96%~97%)显著高于茎生叶(87%~92%),这可能得益于剑叶比茎生叶更接近籽粒。因此,解析玉米剑叶的遗传基础、鉴定控制玉米剑叶的数量性状位点(quantitative trait locus,QTL)和挖掘相关候选基因对培育剑叶性状适中的优良玉米新品种具有重要意义。
玉米剑叶受气候和土壤等环境因素影响,在光照、温度、湿度、水分、肥料和生长空间等环境因素适宜的条件下易出现[2]。同时,玉米剑叶也受遗传因素控制。Cantrell 等[5]利用2 个长剑叶玉米自交系(A639 和A509)和2 个无剑叶自交系(W153R和A634)构建的4 个F2群体对玉米剑叶面积的广义遗传力进行分析发现,玉米剑叶面积的广义遗传力变化范围为30%~76%,表明玉米剑叶面积受遗传因素控制。尽管研究人员围绕玉米剑叶从栽培、生理和遗传等方面开展了相关研究并取得了一定的成果,但利用现代玉米(Zea maysssp.mays)自交系与玉米野生祖先种大刍草(Zea maysssp.parviglumis)构建的渗入系群体解析玉米剑叶遗传基础的研究尚未见报道。
大量考古学和遗传学研究表明,玉米是大约1万年前由分布于墨西哥西南部的大刍草(Zea maysssp.parviglumis)驯化而来[6-8]。在长期选择驯化过程中,现代栽培玉米与大刍草呈现出巨大的形态结构差异[9-11],具体表现为大刍草多分蘖、多穗、丛生、茎秆细、易落粒、籽粒单行互生且外围包裹有坚硬的果壳,而现代栽培玉米则分蘖很少或没有分蘖、茎秆粗壮、不易落粒、籽粒多行且完全裸露。此外,现代栽培玉米的遗传多样性也严重降低。Wright 等[12]利用14 个代表性的玉米(Zea maysssp.mays)自交系和16 个大刍草(Zea maysssp.parviglumis)自交系材料,对774 个基因片段的SNP多样性进行分析,发现现代栽培玉米仅保留了大刍草中约57%的遗传多样性。玉米剑叶也被认为是一种返祖现象。因此,利用现代玉米自交系与玉米野生祖先种大刍草构建的渗入系群体解析玉米剑叶的遗传基础,不仅可以增强对玉米驯化过程的理解,也为玉米剑叶性状的遗传改良提供了基因资源。
本研究利用一套由玉米自交系W22 与玉米野生祖先种大刍草杂交衍生得到的、包含866 个家系的渗入系材料,进一步结合19 838 个SNP 分子标记,对玉米剑叶进行QTL 定位分析,旨在为克隆玉米剑叶相关基因和分子标记辅助选择育种提供参考。
1 材料与方法
1.1 试验材料
试验材料是在美国玉米种质资源中心(Maize Coop Stock Center)获得的以玉米野生祖先种大刍草作为供体亲本、现代玉米自交系W22 作为受体亲本,通过1 次杂交、2 次回交和3 次自交得到的BC2S3渗入系群体。该群体包含866 个家系,且利用简化基因组测序技术获得了覆盖玉米全基因组的19 838 个SNP 分子标记。
1.2 田间种植
2019 年春季,在湖南省浏阳市(113.6°E,28.2°N)国家农作物品种区域试验站种植上述玉米-大刍草渗入系群体。田间试验采用扩增式不完全随机区组设计,每个小区种植2 行,每行15 株,株距25cm。每垄种植2 个家系,垄高15cm,垄宽70cm,沟宽30cm。根据当地常规大田生产管理措施进行水肥管理和病虫草害防治。
1.3 表型调查
参照Wills 等[13]调查玉米多穗性状的方法,每个家系材料从第2 株开始,连续调查10 个单株。如果一个家系材料10 个单株中出现剑叶长度大于2cm 的单株,则将该家系的表型值记为1;如果一个家系材料10 个单株中没有出现剑叶长度大于2cm 的单株,则将该家系的表型值记为0;如果一个家系材料调查的单株数小于3,则将该家系的表型值记为NA(not available)。
1.4 数据处理
利用SAS 软件(v9.1;SAS Institute,Cary,NC)计算每个家系材料表型值的最佳线性无偏估计值(best linear unbiased prediction,BLUP),计算得到的BLUP 值用于QTL 定位分析。利用R 软件(v4.1.2)进行表型相关性分析。
1.5 QTL 定位
利用R/qtl 的多QTL 模型进行QTL 定位分析,具体参照Huang 等[14]的方法。首先利用scanone 命令进行QTL 简单区间定位分析,并采用置换检验(Permutation test)10 000 次的方法确定显著QTL的似然函数比值对数值(logarithm of odds,LOD)阈值(α=0.05)。进一步拟合多QTL 模型,并利用refineqtl 命令优化QTL 位置,然后利用addqtl 命令检测是否存在其他可以显著改善模型的QTL。如果检测到新的QTL,则重新拟合多QTL 模型,重复此过程,直至检测不到新的可以显著改善模型的QTL。最后利用fitqtl 命令计算所有QTL 解释的总表型变异和单个QTL 的表型贡献率。QTL 置信区间定义为QTL 的LOD 峰值下降1.5 对应的区间。依据McCouch 等[15]提出的方法进行QTL 命名。
1.6 基因组织表达特异性分析
玉米苞叶RNA-seq 测序产生的原始数据下载于美国国立生物技术信息中心(National Center for Biotechnology Information,NCBI),数据编号分别为SRR15988094、SRR15988095 和SRR15988097。首先利用fastp 软件对下载的RNA-seq 数据进行质量控制,然后利用Salmon 软件将质量控制后产生的数据比对到玉米B73 参考基因组(RefGen_V4)。唯一比对到参考基因组的数据用于计算TPM(Transcripts Per Million)值。其他9 个组织(根、茎、成熟叶、未成熟叶、茎尖分生组织、花丝、种子、幼嫩的雌穗、幼嫩的雄穗)的FPKM(Fragments Per Kilobase Million)表达数据下载于MaizeGDB(http://www.maizegdb.org)数据库。利用R 软件将FPKM 值转换为TPM 值,转换公式如下,TPMi=(FPKMi/∑jFPKMi)×106。
将校正后的TPM 值进一步换算为以10 为底的对数值,并用TBtools[16]软件的HeatMap 功能绘制基因的表达热图。
2 结果与分析
2.1 渗入系群体剑叶表型分析
调查渗入系群体剑叶表型,发现部分家系材料在雌穗苞叶顶端长有较长的剑叶(大于20cm),而大部分家系材料则完全没有剑叶(图1a)。进一步对雌穗苞叶生长剑叶的位置进行观察,发现有些家系材料并不是最上端的苞叶长有最长的剑叶,而第2~4 片苞叶也可能长有最长的剑叶(图1a)。对渗入系群体866 个家系材料进行表型比例分析(图1b)发现,68%的家系材料不长剑叶,仅有29%长有剑叶,另外3%没有可用的表型值。

图1 渗入系群体剑叶表型分析Fig.1 Phenotypic analysis of the flag leaf in the introgression line population
2.2 剑叶与其他性状相关性分析
剑叶作为玉米雌穗的重要组成部分,可能影响玉米籽粒产量和植株发育。利用R 软件分析剑叶与粒长、粒宽、粒厚、百粒重、株高[17]、穗位高[17]、茎长[18]、茎粗[18]、种子出苗率[19]、叶绿素含量[20]等性状的相关性发现,剑叶与粒宽(r=-0.11)、粒厚(r=-0.01)和百粒重(r=-0.16)等产量性状呈显著负相关(P<0.05),而与株高(r=0.08)、穗位高(r=0.09)、茎长(r=0.11)和叶绿素含量(r=0.11)等性状呈显著正相关(P<0.05),但与粒长(r=0.02)、茎粗(r=0.01)和种子出苗率(r=0.006)等性状呈不显著正相关(P>0.05)(图2)。

图2 剑叶与其他性状相关性分析Fig.2 Correlation analysis between flag leaf and other traits
2.3 QTL 定位分析
利用R/qtl 的多QTL 模型对玉米-大刍草渗入系群体剑叶性状进行QTL 定位分析。首先采用置换检验10 000 次的方法确定检测显著QTL 的LOD阈值为4.28,结果共检测到9 个控制玉米剑叶的QTL(图3),分别位于第1、2、3、4、5、7 和8号染色体,其中第1 和第5 号染色体均含有2 个QTL,表明玉米剑叶属于由多基因控制的数量性状。

图3 渗入系群体剑叶QTL 图谱Fig.3 QTL mapping for flag leaf in the introgression line population
由表1 可知,共检测到9 个控制玉米剑叶的QTL,分别命名为qFL1-1、qFL1-2、qFL2-1、qFL3-1、qFL4-1、qFL5-1、qFL5-2、qFL7-1和qFL8-1,各QTL 置信区间两侧的分子标记分别为m00191 与m01743、m04426 与m05704、m08696 与m12985、m16430 与m16819、m21495 与m22749、m25472与m25799、m29042 与m29724、m39972 与m40321、m42951 与m44940。9 个QTL 共解释30.94%的表型变异。单个QTL 的表型贡献率变幅为1.76%~11.51%,加性效应的变幅为-0.03~0.12,LOD 峰值的变幅为4.49~27.49。检测到的9 个QTL 中仅有qFL3-1的表型贡献率(11.51%)大于10%,其余8个QTL 的表型贡献率均小于5%,表明该套群体中玉米剑叶性状由1 个主效QTL 加多个微效QTL 控制。进一步分析QTL 加性效应方向,发现9 个QTL中的7 个QTL 均来自玉米等位基因降低剑叶表型值,表明剑叶在玉米长期驯化和改良过程中可能受到定向选择,这也与大部分现代栽培玉米不具有剑叶一致。

表1 剑叶QTL 定位结果Table 1 QTL mapping results for flag leaf
2.4 qFL3-1 位点候选基因挖掘
由QTL 表型效应分析可知,在所有检测到的9个QTL 中,qFL3-1具有最大的表型贡献率,可以解释11.51%的表型变异(表1)。qFL3-1置信区间的物理距离为3.14Mb,遗传距离为0.97cM(图4a)。进一步根据玉米B73 参考基因组(RefGen_V4)数据库(http://www.maizegdb.org),在玉米第3 号染色体qFL3-1的置信区间内共检索到31 个注释基因(图4b)。利用玉米在线数据库分析31个基因在玉米苞叶、根、茎、成熟叶、未成熟叶、茎尖分生组织、花丝、种子、幼嫩雌穗和幼嫩雄穗等10 个不同组织器官中表达特异性,发现31 个基因中仅有5 个基因特异性在玉米苞叶中高表达,而在其他组织器官均表达量较低,分别是Zm00001d042099、Zm00001d042104、Zm00001d042117、Zm00001d042134和Zm00001d042135(图4c)。
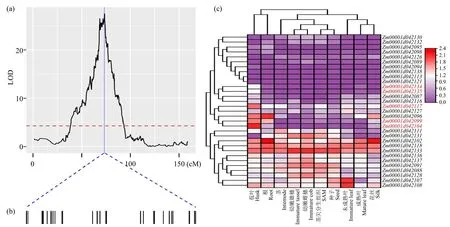
图4 qFL3-1 位点候选基因分析Fig.4 Candidate gene analysis of qFL3-1
基于玉米B73 参考基因组(RefGen_V4)注释信息,对筛选出的5 个基因进行基因功能注释分析,发现Zm00001d042099和Zm00001d042104均编码谷胱甘肽S-转移酶,Zm00001d042117编码一个吡咯啉-5-羧酸还原酶,而Zm00001d042134和Zm00001d042135均编码氨基酸通透酶(表2)。虽然谷胱甘肽S-转移酶、吡咯啉-5-羧酸还原酶和氨基酸通透酶在植物生长发育、次生代谢和逆境胁迫响应过程中起着重要的作用,但相关基因的生物学功能仍有待继续利用基因工程方法进行证实和研究。

表2 qFL3-1 候选基因Table 2 Candidate genes of qFL3-1
3 讨论
玉米剑叶是大部分温带甜玉米雌穗的典型特征,具有较长剑叶的鲜食玉米一般更受广大消费者的青睐,因而具有更大的市场潜力,但目前关于玉米剑叶的遗传基础仍然未知。本研究利用由玉米自交系W22 与玉米野生祖先种大刍草构建的包含866 个家系的渗入系群体,结合均匀覆盖玉米全基因组的19 838 个SNP 分子标记,采用R/qtl 的多QTL 模型对玉米剑叶进行QTL 定位分析,为克隆玉米剑叶相关基因和分子标记辅助选择育种提供参考依据。
Cantrell 等[3]在3 个不同环境下完全去除4 个不同玉米材料的剑叶,发现4 个玉米材料产量的平均降幅为2.6%,表明玉米剑叶对维持玉米高产稳产具有重要作用。本研究对玉米剑叶与粒长、粒宽、粒厚、百粒重、株高、穗位高、茎长、茎粗、种子出苗率、叶绿素含量等性状的相关性进行分析发现,剑叶与粒宽、粒厚和百粒重等产量性状呈显著负相关,而与株高、穗位高、茎长和叶绿素含量等性状呈显著正相关,但与粒长、茎粗和种子出苗率等性状呈不显著正相关,结果表明玉米剑叶与玉米产量性状呈负相关,这与Cantrell 等[3]研究结果相反,可能主要有以下两方面的原因,首先玉米产量是复杂的数量性状,由单位面积有效穗数、穗粒数和百粒重等3 个因素构成,本研究只分析了剑叶与百粒重等籽粒性状的相关性,没有分析剑叶与单位面积有效穗数和穗粒数的相关性,可能是造成结果不同的主要原因;其次是Cantrell 等[3]的研究利用了6 个不同的玉米材料,但仅有其中的4 个玉米材料在完全去除剑叶的条件下显著降低了玉米产量,而其他2 个玉米材料则对产量无显著影响,表明不同遗传背景的玉米材料对分析玉米剑叶与产量的相关性影响较大,这可能是造成结果不同的另一个重要原因。
数量性状呈连续性变异,由多基因控制并易受环境影响。数量性状的遗传结构特征一般可以分为以下3 种类型[21],由少数主效基因控制、由1 个主效基因与多个微效基因控制和由多个微效基因控制。本研究共检测到9 个控制玉米剑叶的QTL,总共可以解释30.94%的表型变异。单个QTL 的表型贡献率变幅为1.76%~11.51%,加性效应的变幅为-0.03~0.12。检测到的9 个QTL 中仅有qFL3-1的表型贡献率(11.51%)大于10%,其余8 个QTL的表型贡献率均小于5%,表明该群体中玉米剑叶的遗传结构是由1 个主效基因加多个微效基因控制。qFL3-1是本研究唯一检测到的控制玉米剑叶的主效QTL,这有利于针对qFL3-1位点开发紧密连锁的功能性分子标记,并利用分子标记辅助选择育种技术定向改良现代栽培玉米的剑叶性状。进一步对9 个QTL 的加性效应方向进行分析,发现9 个QTL 中的7 个QTL 均来自玉米等位基因降低剑叶表型值,包括qFL3-1位点,表明剑叶在玉米长期驯化和改良过程中可能受到定向选择,这也与大部分现代栽培玉米不具有剑叶的现象一致。
根据玉米B73 参考基因组(RefGen_V4)信息,在qFL3-1位点置信区间内共检索到31 个注释基因。进一步利用玉米在线数据库分析31 个基因在玉米苞叶、根、茎、成熟叶、未成熟叶、茎尖分生组织、花丝、种子、幼嫩的雌穗和幼嫩的雄穗等10个不同组织器官中表达特异性,发现31 个基因中仅有5 个基因特异性地在玉米苞叶中高表达,分别是编码谷胱甘肽S-转移酶的Zm00001d042099和Zm00001d042104,编码吡咯啉-5-羧酸还原酶的Zm00001d042117,编码氨基酸通透酶的Zm00001d042134和Zm00001d042135。谷胱甘肽S-转移酶是植物体内普遍存在的一类多功能蛋白,在植物生长发育、次生代谢以及应对生物和非生物胁迫过程中发挥重要作用[22]。吡咯啉-5-羧酸还原酶是植物体内脯氨酸最后合成的关键酶[23]。研究[24-26]表明,植物体内游离脯氨酸除了响应生物和非生物逆境胁迫外,还可以调控植物发育、开花和繁殖等过程。氨基酸通透酶参与植物体内氨基酸的转运并在调控植物生理过程中具有重要作用[27]。
4 结论
利用866 份玉米-大刍草渗入系材料结合19 838 个SNP 分子标记,对玉米剑叶进行了高精度的QTL 定位分析,共检测到9 个控制玉米剑叶的QTL,单个QTL 的表型贡献率变幅为1.76%~11.51%,表明玉米剑叶属于典型的由多基因控制的数量性状。玉米剑叶与株高、穗位高、茎长和叶绿素含量呈显著正相关,与百粒重、粒宽和粒厚呈显著负相关,表明剑叶与玉米其他重要性状之间呈现复杂的相互关系,共同构成了玉米植株的表型特征。进一步利用转录组数据对最大效应QTL(qFL3-1)进行候选基因挖掘,筛选出Zm00001d042099、Zm00001d042104、Zm00001d042117、Zm00001d042134和Zm00001d042135这5 个候选基因。
