铁肥施用、生物强化与人体铁素营养*
2023-08-03张育维
黄 俊, 张育维, 汪 洪
(1.安徽省桐城市种植业管理中心 安徽桐城 231400;2.中国农业科学院农业资源与农业区划研究所/国家化肥质量检验检测中心〔北京〕/农业农村部农产品质量安全肥料源性因子风险评估实验室〔北京〕 北京 100081)
0 前言
铁是一种金属元素,原子序数为26,平均相对原子质量为55.845,常见价态有0 价、+2 价、+3 价。 Fe(Ⅱ)作为电子供体,Fe(Ⅲ)作为电子受体,参与众多氧化还原的生化反应。 铁是人体必需的微量元素,是人体血红蛋白、肌红蛋白、血红素酶或非血红素化合物等的重要组成,参与氧的运输和贮存及多种金属酶的合成,调节组织呼吸,防止疲劳,促进人体生长、神经发育,增强人体免疫力[1-2]。 联合国在《2030 年可持续发展议程》中提出的可持续发展目标(SDGs) 的第二项(SDG-2)“零饥饿”目标,旨在通过获得安全营养,到2030 年结束所有形式的饥饿和营养不良[3],但粮食不安全和营养不良仍是全球面临的最大挑战。 根据联合国粮食及农业组织等发布的《2022 年世界粮食安全和营养状况》,2021 年全球有7.02 亿~8.28 亿人陷入饥饿,预计到2030 年全世界仍将有近6.7 亿人(占全球总人口的8%)面临饥饿,发展中国家特别是人口大量增加的非洲和南亚地区的情况较为严重[4]。
铁是植物生长发育必需的微量元素,参与植物体内叶绿素合成、光合作用、电子传递等生理代谢过程[5]。 第一次农业绿色革命提高了谷类作物的产量,增加了全球粮食供给,但忽视了粮食品质对环境和人体健康的影响,出现了“隐性饥饿”问题,即微量元素营养无法满足人体需求,尤其是铁、锌、维生素A 等缺乏[6-7]。 发展中国家低收入贫困家庭多以禾谷类籽粒为主要食品,谷类作物的微量元素不仅含量较低,而且大多数铁、锌等聚集在谷物籽粒的糊粉层中,在粮食加工过程中因糊粉层的分离导致损失[8]。 另外,作物籽粒中富含植酸,容易与铁、锌等结合形成不溶配合物,而人体消化系统缺少植酸水解酶,影响对铁等微量元素的吸收。 因此,在以谷物为主食、经济负担能力较弱的发展中国家人群中普遍存在铁、锌等微量元素缺乏问题[9]。 “隐性饥饿”会削弱免疫系统,使人们容易受到腹泻威胁、病毒感染,儿童容易出现发育迟缓、失明、贫血等症状;全球近5 亿15~49 岁的女性患有衰弱性贫血,缺铁是主要原因。 因营养不良(包括蛋白质和热量摄入不足以及微量营养素缺乏)导致整个非洲的经济损失平均占国内生产总值(GDP)的11%[10]。 本文就人体铁素摄入与人体健康、生物强化与作物铁肥施用等方面的研究进行了综述。
1 铁与人类健康
人体铁营养的维持与人体内铁含量密切相关,缺铁会导致人出现疲劳和虚弱、认知和免疫系统受损、智力发育不良等问题[11-12]。 缺铁是人体贫血的主要原因,缺铁性贫血(IDA)是全世界最普遍的营养缺乏症,影响到全球33%的非孕妇、40%的孕妇和42%的儿童[13]。 估计超过60%的全球人口存在缺铁或潜在缺铁的问题,其中约25%的人口存在严重的缺铁性贫血问题[14-15]。 缺铁风险较高的人群是学龄前儿童和育龄妇女,铁营养不良是2019 年全球0 ~9 岁、10~24 岁人群残疾调整生命年(DALYs)的主要风险因素,因素排序分别为第7 位和第8 位[16]。 亚洲开发银行和联合国儿童基金会(UNICEF)的一项研究发现,缺铁性贫血会导致从事繁重体力劳动的工人生产力降低17%,从事中等体力劳动的人群生产力降低5%[17]。 过量铁离子造成人体“铁浓化”,通过Fenton 反应引发氧化性胁迫,发生非凋亡性细胞过度氧化坏死即铁死亡(ferroptosis)[18]。
刘建欣等[19]开展的Meta 分析结果显示:我国0 ~14 岁儿童缺铁性贫血患病率在2006—2010 年为 22.6%, 2011—2015 年为 21.9%,2016—2020 年下降至16.8%;88.7%的患病儿童表现为轻度贫血,11.3%的表现为中重度;西北、西南地区儿童患病率分别为31.9%、28.3%;农村儿童患病率(25.6%) 远高于城市儿童的(9.1%)。 2015—2019 年,国家卫生健康委员会中国疾病预防控制中心、国家癌症中心、国家心血管病中心开展了中国居民慢性病与营养监测,覆盖31 个省(区、市)近6 亿人口,现场调查超过60 万人,根据监测结果编写了《中国居民营养与慢性病状况报告(2020 年)》。 该报告中我国18 岁及以上年龄居民贫血率为8.7%,6 ~17 岁儿童和青少年贫血率为6.1%,孕妇贫血率为13.6%,与2015 年相比均有显著下降。 《国民营养计划(2017—2030 年)》要求到2020 年降低人群贫血率,其中5 岁以下儿童贫血率控制在12%以下,孕妇贫血率下降至15%以下,老年人群贫血率下降至10%以下,贫困地区人群贫血率控制在10%以下;到2030 年,进一步降低重点人群贫血率,其中5 岁以下儿童和孕妇贫血率控制在10%以下。世界卫生大会的目标是期待到2025 年实现育龄妇女贫血率减少50%。
2 人体铁营养摄入
营养是人体维持生命、生长发育和健康的重要物质基础。 世界卫生组织(WHO)推荐的每日饮食铁摄入量:5 岁以下儿童2 mg/kg(体质量,下同),5~12 岁儿童30 mg/kg,育龄妇女60 mg/kg[20]。美国国家科学院医学研究所(IOM)食品和营养委员会(FNB)制定的膳食参考摄入量(DRI)中,提供了铁和其他营养素的摄入量建议[21]。 DRI 是一组参考值的总称,用于规划和评估健康人体的营养摄入,因年龄和性别而异,包括:推荐膳食量(RDA),即每日平均摄入量足以满足几乎所有(97%~98%)健康个体的营养需求,通常用于为个人制定营养充足的饮食;充足摄入量(AI),即假设摄入量达到这一水平以确保营养充足,通常在无法确定RDA 时使用;估计平均需求量(EAR),即估计每日平均摄入量水平,以满足50%健康个体的需求,通常用于评估人群的营养摄入,并为他们制定营养充足的饮食,也可用于评估个体的营养摄入;容许摄入量上限(UL),即不会对健康造成不良影响的每日最大摄入量。 美国居民膳食营养推荐铁摄入量和可耐受摄入量上限见表1。
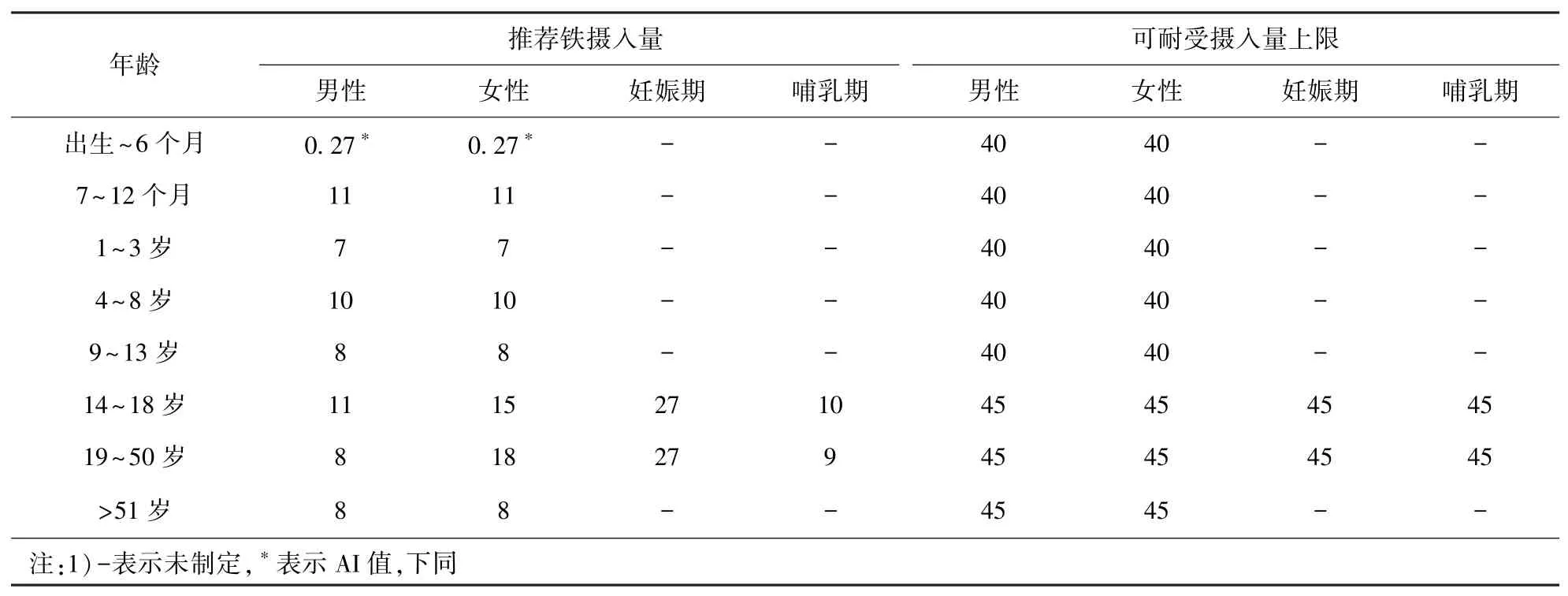
表1 美国居民膳食营养推荐铁摄入量和可耐受摄入量上限mg/d
我国在《国民营养计划(2017—2030 年)》中建议,贫血、消瘦等营养不良人群要在合理膳食的基础上,适当增加瘦肉类、奶蛋类、大豆和豆制品的摄入,保持膳食的多样性,满足身体对蛋白质、钙、铁、维生素A、维生素D、维生素B12、叶酸等营养素的需求;增加含铁食物的摄入或者在医生指导下补充铁剂来纠正贫血。 行业标准《中国居民膳食营养素参考摄入量 第3 部分:微量元素》(WS/T 578.3—2017)于2017 年发布,其中中国居民膳食营养参考铁摄入量见表2。

表2 中国居民膳食营养参考铁摄入量
为改善人体的铁营养健康,发达国家采取的主要策略有饮食多样化、强化食品和膳食补充剂[13,22-25]。 美国联邦政府在《2020—2025 年美国人膳食指南》中指出:食物提供了一系列营养素和其他有益健康的成分,营养需求应主要通过食物来满足。 因此,饮食多样化、多食富铁食物,可增加人体对铁的摄入。 食物中的铁分为血红素铁和非血红素铁两种形式,瘦肉、海鲜等是血红素铁的膳食来源[26],坚果、豆类、蔬菜和强化谷物制品是非血红素铁的膳食来源[21,27-28]。 肉类、海鲜和维生素C 可提高非血红素铁生物利用度,混合饮食中铁生物有效性为14%~18%,素食中铁生物有效性为5%~12%[21,27-28]。 人体吸收非血红素铁通常较血红素铁困难,且受到食物中各种因素的影响,血红素铁生物利用率较高。 抗坏血酸、肉类、家禽和海鲜可增强非血红素铁吸收,植酸盐(存在于谷物和豆类中)和某些非动物性食物(如谷物和豆类)具有相反作用。 与其他铁吸收抑制剂不同,钙可能会降低非血红素和血红素铁的生物利用。 主要农作物和蔬菜是非血红素铁的良好来源,如绿叶蔬菜、豆类、坚果和完整的谷物籽粒。植酸盐、草酸盐、单宁和钙在肠中结合非血红素铁,会降低人体对铁的吸收[29]。 蓝丰颖等[29]采用多阶段整群抽样,抽取全国6 座城市、2 个农村1 600 名3 ~12 岁的儿童进行调查,结果发现:铁摄入量低于平均需要量(EAR)的儿童占17.4%,高于可耐受最高摄入量(UL)的占2.9%;对受访儿童膳食铁贡献最大的食物为谷物和肉类;农村儿童动物源铁的摄入比例显著低于城市的。
(1)膳食补充剂,即在市场上销售的含铁补充剂。 14%~18%的美国人使用含铁补充剂;含铁补充剂的使用因年龄和性别存在差异,12 ~19 岁的青少年使用率为6%,哺乳期妇女的为60%,孕妇的为72%[30]。 补充剂中常用的铁包括亚铁和三价铁盐,如硫酸亚铁、葡萄糖酸亚铁、富马酸亚铁、乙二胺四乙酸铁、柠檬酸铁、硫酸铁等。富马酸亚铁含铁33%,硫酸亚铁含铁20%,葡萄糖酸亚铁含铁12%[31]。 因亚铁盐具有较高的溶解度,故膳食补充剂中亚铁的生物利用率比三价铁的高[27]。 高剂量补充铁(超过45 mg/d)可能会导致胃肠道副作用,如恶心、便秘等[21]。 补充其他形式的铁,如天然植物铁蛋白、乳铁蛋白、血红素铁多肽、羰基铁、铁氨基酸螯合物和多糖铁复合物,对肠胃的副作用可能比亚铁或铁盐小。 钙可能会干扰铁的吸收,尽管这种影响尚未确定,但一些专家建议不要同时服用钙和铁补充剂。
(2)食品强化工程。 美国、加拿大和许多其他国家在食品中添加铁,开展食品强化,销售铁强化的面粉,婴儿配方奶粉中也可添加铁[32]。 不同国家对铁强化食品的规定有所不同。 在中国,可可粉、婴儿食品和盐中允许加入硫酸亚铁的最大剂量分别为11 ~12、300 ~500、3 000 ~6 000 mg/kg。在日本,可添加到食用橄榄油中的葡萄糖酸亚铁的最大剂量为150 mg/kg[25]。
3 作物遗传育种与铁生物强化
药物防治、食品防御及饮食多样化等措施已在发达国家广泛应用并取得了良好的效果。 但发展中国家经济技术相对落后,政策扶持力度不足,食品和药品防治作用有限,人群饮食习惯多以含铁量较低的谷物类为主,导致在发达国家行之有效的措施在发展中国家并不能得到很好的实施。生物强化工程被认为是一种安全、经济、稳定、有效的策略[33-35]。 在 Ross Welch 教授、 Robin Graham 教授和Howarth ″Howdy″ Bouis 博士等科学家的倡议下,2004 年国际农业研究磋商委员会组织启动了HarvestPlus 计划,即生物强化项目。该项目针对发展中国家人群铁、锌、维生素A 等微营养普遍贫乏的状况,旨在培育和推广富含微量营养元素的作物新品种,克服植物性食物中微量营养元素不足的缺陷,提高膳食营养水平。 美国比尔和梅林达·盖茨基金会、美国国际开发署、世界银行和丹麦外交部国际发展署资助该项目,国际热带农业中心和国际农业政策研究所负责协调,在全球特别是在发展中国家建立了研究联盟进行具体实施[35]。 实现生物强化的主要途径是植物育种和农艺栽培措施,提高粮食作物籽粒中微量元素的含量和生物有效性,提高作物可食用部分铁含量,从而改善通过主食提供的铁素营养[34-36]。 遗传育种的生物强化措施涉及基因工程和常规育种。 精米和小麦籽粒中的铁含量生物强化目标分别为13~15、59 mg/kg,而普通水稻的籽粒铁含量低7.5 倍,普通小麦籽粒中的铁含量仅为20~40 mg/kg[37-39]。 HarvestPlus 计划旨在通过作物育种研究项目,研发小麦、水稻和大豆等的新品种,铁含量提高了10 mg/kg[40]。 HarvestPlus项目与国际干旱地区农业研究中心(ICARDA)合作开发了铁强化的扁豆品种[41],2010 年首次在孟加拉国和叙利亚上市,目前孟加拉国、印度、尼泊尔和叙利亚已经发布了12 种富铁扁豆;与国际半干旱热带作物研究所(ICRISAT)合作培育的富铁小米,在印度和尼日尔已经发布了2 种开放授粉品种和9 种小米杂交种,铁含量达到77 mg/kg,珍珠小米中铁生物利用率达8%[42];研发了近60 个富铁豆类品种,铁含量比常规品种的高90%,产量也很高,并具有农艺、口感和烹饪特性,在11 个国家得到推广,定期食用可为育龄妇女和儿童提供80%的日铁需求,迄今已在布隆迪、刚果民主共和国、卢旺达等非洲国家推广了42 个品种,在玻利维亚、巴西、哥伦比亚、萨尔瓦多、危地马拉、洪都拉斯、尼加拉瓜、巴拿马等推广了23 个品种[43]。 在卢旺达开展的一项针对女大学生的生物效能试验发现,高铁豆类改善了人体铁营养,认知能力、大脑功能和工作效率明显增强[44]。 缺铁的女大学生(18 ~27 岁)每天吃富铁豆,坚持4.5 个月后,体内血红蛋白、血清铁蛋白和铁浓度明显升高[45],记忆力、注意力及完成每天体力劳动的能力明显提升[46-48]。
分子标记辅助选择、高通量表型和特定性状的基因型筛选等分子育种与转基因组技术已应用于铁生物强化的遗传育种计划中[49]。 全基因组关联研究(GWAS)加速遗传标记的开发,有助于更快地确定目标基因。 通过数量性状基因座(QTLs)定位和GWAS 鉴定了与水稻籽粒中铁和锌积累有关的遗传位点,在12 条水稻染色体上已鉴定出93 个与铁含量相关的QTLs,其中数量最多的位于第1、3、7 号染色体,17 个稳定的QTLs位于第1~4、6~8、10、11 号染色体[50]。 表达水稻烟酰胺合成酶(OsNAS2)和大豆铁蛋白(SferH-1)基因有助于提高胚乳中铁和锌含量[51-52]。 过表达OsYSL2和铁储存蛋白铁蛋白的转基因水稻品系体内锌和铁的浓度分别增加了1.6 倍和6 倍[53]。 作物籽粒中植酸含量较高,通过增加烟酰胺含量或过表达NAS基因可降低植酸含量,增强小麦和玉米的铁生物利用[38]。 反向育种、寡核苷酸介导诱变、基因组编辑技术和高通量表型等现代技术,为微量营养生物强化育种战略带来全新的变化[54-55]。 遗传育种的生物强化措施能有效提高作物中铁的含量和有效性,但仍面临耗时长等问题,而田间应用效果、与重金属累积相关的健康风险以及在铁供应不足的背景下能否发挥其潜力等还有待验证[40,55-56]。 遗传生物强化手段目前仍处于早期阶段。
4 铁肥施用与作物铁强化
4.1 植物铁吸收运输与生物强化
4.1.1 土壤铁供应与植物铁吸收
谷物中铁的含量与土壤中铁的有效性密切相关。 铁是地壳中第四大元素,在土壤中含量丰富,但是铁在土壤(尤其是碱性土壤)中主要以不溶性的氧化铁和氢氧化铁的形式存在,游离铁总的物质的量浓度一般不超过10-15mol/L,无法满足植物正常生长发育对铁的需求(10-9~10-4mol/L),导致植物缺铁现象普遍存在,影响作物产量和品质[57]。 全球有1/3 的耕地是石灰性土壤,植物缺铁现象较为严重[58]。
为了从土壤中获得足够的铁,植物在长期的进化过程中,形成了2 种不同的铁吸收策略。 双子叶植物和非禾本科单子叶植物属于策略Ⅰ植物,根系向根际释放有机酸、酚类和黄素类化合物等,促进难溶性铁化合物溶解、螯合;根系质膜上H+-ATPase 泵分泌H+,根际土壤酸度降低,土壤中的可溶性铁含量增加[59]。 根系质膜上诱导Fe3+螯合物还原酶的表达,将Fe3+还原成Fe2+,通过高亲和力的Fe2+转运蛋白转入根细胞[5]。 Fe3+螯合物还原酶基因FRO2已在拟南芥、番茄、黄瓜、花生等相继被克隆[60-63]。 Eide 等[64]的研究发现,拟南芥根表细胞质膜上铁转运蛋白基因AtIRT1属于ZIP 转运体家族,具有广泛的底物特性,除了能转运铁外,同时还能转运锌、锰和镉等金属元素[64]。 在豌豆、番茄、花生等植物根表也相继发现了IRT 转运蛋白[65]。 NRAMP 等蛋白也参与Fe2+的转运[66]。
禾本科植物采用策略Ⅱ适应土壤中铁的缺乏。 根系分泌麦根酸类植物高铁载体(PS),与土壤中的Fe3+螯合形成Fe(Ⅲ)-PS,然后在专一性转移蛋白YS1 或YSL 作用下,将Fe(Ⅲ)-PS 运输到根细胞内,释放出Fe3+供植物体内生理代谢利用[67]。 PS 由S-腺苷甲硫氨酸通过一系列的酶促反应合成得到。 参与麦根酸生物合成的酶包括烟酰胺合成酶(NAS)、烟酰胺氨基转移酶和双脱氧麦根酸合成酶[67]。 Nozoye 等[68]在水稻和大麦根中分离和鉴定了PS 转运蛋白TOM1。
植物体内存在复杂的铁稳态基因调控网络,包括转录和翻译水平上的控制来调节铁稳态,既可保证其对土壤中缺铁胁迫进行应答与有效铁的吸收,又可避免积累过量的铁对其自身生长发育产生毒害[69-71]。 拟南芥中基本螺旋-环-螺旋(bHLH)转录因子的关键调控网络包括转录因子FIT、Ib 亚组、IVc 亚组和URI(bHLH121),这些转录因子调节拟南芥对铁的吸收[70]。 植物激素生长素、乙烯、细胞分裂素、脱落酸、油菜素内酯、茉莉酸、褪黑素以及一氧化氮、钙离子信号、锌指蛋白等信号系统和分子被证实参与调控植物对铁的吸收[70,72-73]。 禾本科作物缺铁胁迫诱导的顺式作用元件IDE1 和IDE2 以及特异结合于该元件上的转录因子IDEF1 和IDEF2,对维持体内铁平衡具有重要的作用[74]。 微生物在缺铁或低铁条件下会产生一些低相对分子质量(500~1 000 Da)的有机化合物,称为铁载体(Siderophores)。 Fe3+-Siderophores 螯合物一部分被微生物自身吸收,进入细胞后还原成Fe2+,供微生物生长发育;另一部分被植物吸收利用[75]。
4.1.2 植物体内铁运输及向籽粒转运
植物细胞有一套精细的机制,控制细胞内游离铁离子的平衡[76]。 通过细胞膜系统运输时,Fe(Ⅲ)通常被还原为Fe(Ⅱ)[77],以避免细胞内产生过多的活性氧,Fe(Ⅱ)通过VIT1 蛋白被运输到籽粒液泡中[78]或Fe(Ⅲ)以铁蛋白形式储存[79-81]。 在细胞质中,铁可能与有机酸、烟碱酰胺(NA)形成复合物,生成Fe(Ⅲ)-柠檬酸盐、Fe(Ⅲ)-NA 和Fe(Ⅱ)-NA[76,82-83]。 植物体内的铁通过木质部和韧皮部进行长距离运输,木质部中的铁大多运输到老叶中,韧皮部中的铁运输到新叶,铁从营养组织经韧皮部运输到正在发育的种子。 铁主要在水稻籽粒糊粉层中以Fe(Ⅱ)和Fe(Ⅲ)形式与植酸结合。 小麦中铁的累积与水稻中的类似,60%或更多的铁在种子麸皮的糊粉层中积累,其次是鳞茎,胚乳中只有少量铁积累[84-85]。
木质部中的铁主要以螯合物如Fe3+-柠檬酸复合物的形式存在[86-88]。 在缺铁条件下,木质部中的柠檬酸盐、苹果酸盐和琥珀酸盐的含量升高[89]。 铁进入胚胎组织需要经历韧皮部铁释放和胚胎铁吸收2 个步骤。 在拟南芥等双子叶植物中,铁首先释放到胚囊液中,再由胚胎吸收[84];拟南芥铁还原酶缺陷3 基因FRD3、水稻体内该基因同源基因FRDL1介导柠檬酸盐和铁向木质部运输[90];铁蛋白1(FPN1)负责拟南芥体内铁向木质部装载[91]。 叶片中的铁可通过YSL 转运蛋白从质外体空间装载到细胞中[92];AtYSL2 参与了铁从木质部到地上部细胞内的分布[93],AtYSL1和AtYSL3 参与了Fe-烟碱酰胺复合物从衰老叶片向花序和种子中的转移、韧皮部中铁的运输。水稻YSL 转运蛋白如OsYSL2,可能参与Fe(Ⅱ)-烟碱酰胺复合物向地上部和种子的转移[94];OsYSL16 有助于Fe(Ⅲ)-脱氧麦根酸通过维管束鞘的运输[95];OsYSL18是Fe(Ⅲ)-脱氧麦根酸的转运载体基因,在花粉、花粉管、叶片和叶鞘接头部位的韧皮部中表达,说明该基因很可能在水稻受精和韧皮部的铁运输中起作用[96]。 寡肽转运蛋白家族OPT3 参与韧皮部中铁的运输,介导地上部到根的信号传导[97]。
4.2 铁肥施用
通过施肥包括土壤施用、叶面喷施、种子包衣等农艺措施,可提高作物籽粒中铁的含量[98]。 当前市场上主要的铁肥品种有以硫酸亚铁为主的无机铁肥、螯合态铁和一些有机物与铁复合形成的铁肥,各种铁肥详情可参见文献[99]。 无机铁肥稳定性差,施入土壤中会迅速转化为不溶形式;螯合态铁肥可克服被土壤固定的缺点,但成本高;有机复合铁肥肥效较低。
土施铁肥能够显著提高果树作物的果实产量和品质[100]。 Zulfiqar 等[101]通过2 年的田间试验发现,土施铁肥处理的面包小麦籽粒产量提高了28%。 Gamble 等[102]于2010—2012 年在阿拉巴马州的2 个高酸度土壤上开展了田间试验,结果表明在犁沟中施用Fe-EDDHA 4.5 kg/hm2的处理提高了大豆产量,但施用柠檬酸铁和FeSO4的处理未能提高大豆产量。 世界上约1/3 的土壤呈碱性,土壤pH 较大,施入土壤中的铁肥易被固定或氧化成植物难以利用的形式,总体来说土施铁肥对作物补铁的效果不是十分理想[99,103]。 基于植物高铁载体脱氧麦根酸(DMA)结构合成了一种新型铁螯合剂脯氨酸-2′-脱氧麦根酸(PDMA)[104]。 PDMA 比DMA 稳定,合成成本较低。 PDMA 已被证实在水培和石灰基质中改善了植物铁营养[104-105]。 盆栽试验结果表明,施用PDMA 可使花生根际不溶性铁溶解,AhYSL1表达增强,改善了花生植株的铁营养;此外,缺铁引起的黄化和生长抑制显著减弱。 值得注意的是,在田间条件下,施用PDMA 可提高花生产量和籽粒铁含量,PDMA 具有作为新型铁肥的潜力[106]。
叶面施肥可避免土壤对铁的固定。 在特定生长时期叶面喷施的铁可以通过茎和叶组织内韧皮部或木质部运输[107],提高谷物铁含量[108-109]。 缺铁土壤上种植的豌豆在籽粒灌浆期间叶面喷施Fe-EDDHA 和FeSO4,可显著提高籽粒中铁的含量,开花期间叶面喷施铁肥是提高成熟谷物中铁含量的合适时间[110]。 Fang 等[111]的田间试验结果表明:叶面喷施铁肥(0.90 kg/hm2)不影响水稻产量、蛋白质含量,可显著提高铁含量37.1%;与不进行叶面喷施铁肥的处理相比,最佳叶面施肥处理的精米中铁含量增加了2.6 倍。 Zulfiqar等[101]的研究发现:2017—2018 年和2018—2019 年叶面喷施0.5%铁溶液处理的小麦籽粒铁含量分别比对照提高了65%和58%。 Yuan 等[112]通过试验发现:与对照相比,叶面喷施铁氨基酸(Fe-AA)肥料显著提高了水稻糙米中的铁含量,平均提高了14.5%;在Fe-AA 中添加1%烟酸(NA)时,糙米中铁含量平均提高了32.5%。
浸种、拌种、种子引发和种子包衣等处理也属于铁肥施用的方式。 种子引发技术主要作用于种子吸水膨胀阶段,通过调控种子吸水状态,激活种子生理代谢过程,使种子质量与植物抗逆能力得到提升。 Zulfiqar 等[101]的试验结果表明:以种子引发方式施用铁肥,小麦籽粒产量提高了33%;通过种子包衣方式施用铁肥,小麦产量增加21%。 Wei 等[113]在种子发芽过程中添加质量浓度0.05~2 g/L 硫酸亚铁,发芽糙米中铁含量提高了1.1~15.6 倍。 采用不同质量浓度(25~600 mg/L)氧化铁纳米颗粒浸泡引发小麦种子12 h,结果表明:400、200 mg/L 处理的高铁基因型(IITR26)和低铁基因型(WL711)小麦的发芽率和芽长均显著增加;25 mg/L 处理的IITR26 和WL711 的籽粒铁含量分别显著增加45.7%和26.8%[114]。
耕作措施如轮作或间作可以改善作物的铁营养。 双子叶植物与禾本科单子叶植物间作促进了双子叶植物对铁的吸收。 Zuo 等[115]观察到,在中国河南省的石灰性土壤中,玉米/花生间作能明显减轻花生的缺铁黄化现象,可提高花生中铁含量和产量。 Gunes 等[116]的田间试验结果表明,小麦和鹰嘴豆间作,可使小麦籽粒中的铁含量、鹰嘴豆籽粒中的铁和锌含量增加。
施用氮肥和硫肥可提高谷物中微量元素的含量[117-118]。 在鹰嘴豆开花期叶面喷施2%尿素400 L/hm2,可提高籽粒中锌和铁的含量[119]。 在大麦孕穗期施用尿素20 kg/hm2(折纯),可提高籽粒中锌和铁的含量[120]。 水稻-小麦种植体系施用硫肥,可提高谷物中锌、铁等元素的含量[118]。
纳米颗粒是指粒径至少在一个维度上小于100 nm 的颗粒,具有小尺寸效应、表面效应、量子尺寸效应和宏观量子隧道效应。 铁纳米颗粒已用于提高作物铁含量[121];α-Fe2O3纳米颗粒预处理豆科种子可促进根系生长[98];施用铁纳米颗粒改善了花生的营养质量、生物量、产量、氮磷代谢和铁强化[122];施用γ-Fe2O3纳米颗粒促进了铁从根部向植物其他部位运输,提高了南瓜中铁的浓度,但Fe(Ⅱ)-EDTA 处理的效果较弱[123];南瓜叶面施用碳涂层铁纳米颗粒有助于将铁从叶片转移到其他部位,但不会影响植物生长[124];铁螯合纳米肥料的应用改善了罗勒植物[125]和水稻[126]产量;施用黄铁矿(FeS2)纳米颗粒预处理菠菜种子,可促进菠菜生长[127];施用螯合铁纳米肥料4 kg/hm2,菠菜叶片中的铁和钾的积累增加[128];铁螯合纳米肥料的施用改善了水稻品质,提高了对氮磷钾的吸收[126]。 纳米颗粒的复杂性和不确定性使得施用纳米肥料进行生物强化面临挑战,一些研究报告了纳米颗粒对植物的生理损伤。 Al-Amri等[129]的研究表明,Fe2O3纳米颗粒会损伤小麦植株根尖细胞,这可归因于活性氧的产生,破坏了细胞膜。 与非纳米形式相比,纳米颗粒被认为对生物和环境系统具有毒性风险,纳米颗粒通过食物链转移到可食用植物部位,进而转移给动物和人类,但这一说法仍需要更多毒理学证据和风险评估研究,综合生命周期来评估纳米肥料对环境与人体健康的影响[130-133]。
5 结语
铁与人体营养健康关系密切,生物强化是改善发展中国家人群铁营养状况的重要途径(图1)。 通过育种培育高铁含量作物品种,采取叶面喷施、土壤施用不同类型的铁肥等农艺措施,改善作物铁素营养,提高农产品中铁的含量,再通过食物链传递以增强人体铁素营养摄入。 建议在“医学-营养学-农学”之间建立起跨学科体系,合作研究肥料-土壤-作物-人体系统中铁的运转与利用效率,综合解决人类铁营养健康问题。
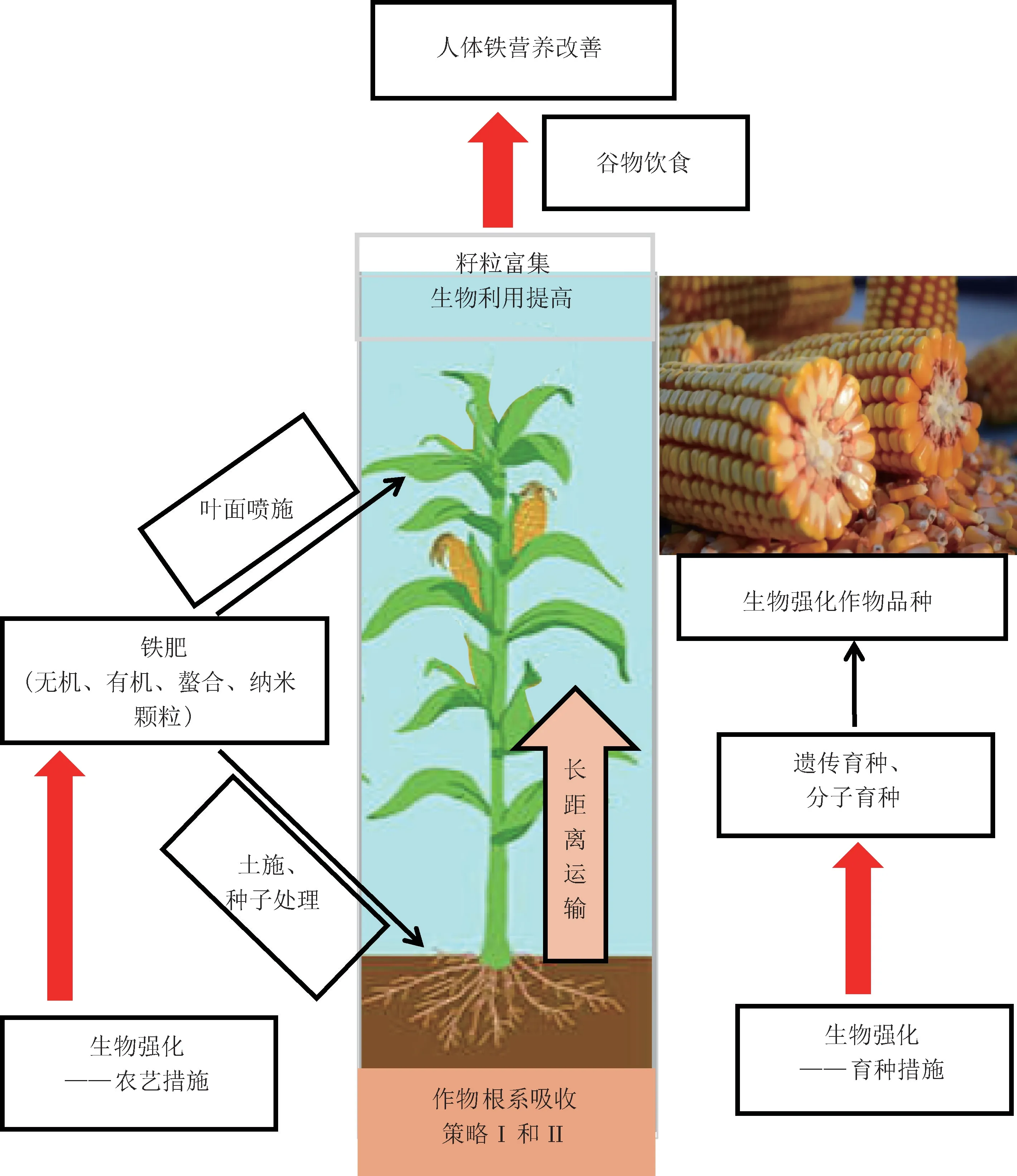
图1 铁肥施用、生物强化与人体铁营养健康
