环境因子及内源物质对果树中花青素调控的研究进展
2023-07-29郑秀文张萌萌
王 硕,郑秀文,刘 冠,张萌萌
(1黑龙江大学现代农业与生态环境学院,哈尔滨 150080;2黑龙江大学/农业微生物技术教育部工程研究中心,哈尔滨 150500;3黑龙江大学生命科学学院/黑龙江省寒地生态修复与资源利用重点实验室,哈尔滨 150080)
0 引言
花青素是植物各组织器官内糖基化的多酚类化合物,又称为花青素,其分子结构为3,5,7-甲基-2-苯基苯并吡喃,是自然界中普遍存在于植物叶、花和水果中的水溶天然色素,其使植物呈现不同颜色。目前已知的花青素有20多种,可供人类食用的有6种,根据结构的不同分为天竺葵素(pelargonidin)、矢车菊素(cyanidin)、芍药花素(peonidin)、飞燕草素(delphinidin)、矮牵牛素(petunidin)和锦葵素(malyidin)[1-2]。
花青素功能多样,它在植物抵抗逆境时发挥着重要作用[3],既可以抵抗病原菌侵染、虫害侵染等生物逆境,也可以抵抗紫外线辐射、低温、旱灾等非生物逆境;花青素还有利于植物着色,使植物具有观赏价值,其色彩可以吸引昆虫传粉、促进植物繁殖[4];此外花青素还具有抗氧化、清除[5]、抗炎、抗感染、防癌、防止心血管疾病等作用。
花青素在苹果、杨梅、李子和梨等果实中积累较多,因此果实不仅色泽鲜艳,而且营养丰富,所以花青素的含量在果树的分子改良育种研究中是一个重要指标。本研究主要从环境因子和内源激素对花青素积累影响的研究进行总结及展望,旨在为果树育种以及花青素的深入研究和利用提供一些参考依据。
1 环境因素对花青素积累的调控作用
1.1 光照
光照强度对花青素的合成影响显著。已有一些研究表明,杨梅、苹果、梨等果树在种植过程中如果经过套袋处理,会导致果实没有颜色,而在阳光下则会呈现红色的果皮[6-9]。而当光强的阈值较低时,花青素的含量会随着光合强度的增强而增多[10]。柳蕴芬等[11]研究了不同水果套袋处理对桃子花色苷含量转变的调控,结果表明套袋处理会降低红肉桃内花青素的积累。也有一些研究表明,苹果果皮的花青素含量与光照强度呈正相关[12]。
在光强增加时,与花青素合成、转运控制有关的基因表达明显提升。例如R2R3MYBs就是重要的正调控因子,其直接影响到花青素生物合成基因的表达[13]。而对于一些负调控因子,如COP1(constitutive photomorphogenic 1),在强光条件下,植物会通过COP1/SPA 酶复合物进行正常的光信号传导[14],以防止花青素的过度积累。此外,花青素合成酶(anthocyanidin synthase,ANS)作为花青素合成通路下游的关键酶,对强光胁迫也可起到抵御作用。有研究表明,植物体对强光的抗性和ANS基因的表达水平之间存在正相关关系[15]。
光质则通过调控花青素合成中的有关基因表达来调控花青素合成,且不同光质对花青素的调控机制不同。Çiğdem等[16]通过对石榴汁液的研究发现,在经过UV-C的处理之后,相比于单一热处理,石榴汁的花青素合成量增加。Kim 等[17]发现,蓝光+UV-B 共照的条件是诱导番茄花青素生物合成的最有效方法:蓝色+UV-B通过高度诱导硝酸还原酶的产生增加一氧化氮的生成,进而调控花青素合成通路中某些基因的表达水平并导致花青素的有效累积。也有研究结果表明[18],在白光、红光与蓝光的照射下,植物愈伤组织的花青素总积累会显著提高。
针对于中华猕猴桃的研究表明,红光下中华猕猴桃中的花青素含量降低,其通过抑制查尔酮合成酶(chalconesynthase,CHS)、二 氢 黄 酮 醇-4-还 原 酶(dihydroflavonol-4-reductase,DFR)、花 青 素 合 成 酶(anthocyaninsynthase,ANS)、糖基转移酶(flavonoid 3-O-glucosyltransferase,UFGT)等酶基因的表达得到抑制,而高蓝光可以提高CHS、DFR等基因的表达水平,进而促进愈伤组织花青素的积累。解潇冬等[19]在蓝光:红光为3:1 环境下发现猕猴桃内遗传基因MYB110的表达量增加,从而推动了鲜红色猕猴桃果实的产生。
1.2 温度
温度是影响花青素积累的显著环境因子,其通过影响植物组织中花青素的稳定性和生物合成路径中相关酶的活性来调控花青素的合成[20]。低温使花青素积累升高,而高温使花青素积累降低。金周雨等[21]通过对蓝靛果的研究发现,当以4℃为基准时,随温度的升高花青素的含量逐渐降低。此外Fang 等[22]通过对李子进行研究发现,在对李子进行20℃处理后,花青素积累相关酶基因的转录水平升高(包括2-氨基苯丙酸解氨酶、查尔酮合酶),进而诱导李子果皮中花青素的积累;而30℃以上温度会导致酶的活性降低,花青素的含量下降。而关于高温下花青素积累减少的原因,也考虑与花青素自身的降解有关[23]。
温度还可以通过影响花青素合成途径中调节基因的表达来调控花青素的合成。Bu 等[24]在苹果中发现一种名为MdLBD37的基因,此基因的表达在高温下急剧上调且对苹果花青素的积累有显著抑制作用。Yu等[25]在对苹果进行低温处理后,发现其ROS1基因的表达受 到 抑制,进 而导致MdCHS、MdCHI、MdF3'H、MdANS、MdUFGT和MdMYB10的启动子甲基化水平降低,增加了花青素的积累。
1.3 矿质元素
矿质元素主要通过影响花青素的稳定性来调控花青素的含量,大多数的矿质元素对花青素的稳定性具有保护作用,并能促进花青素的合成,少部分矿质元素无明显作用,极小部分甚至会抑制花青素的合成。
李炎林等[26]研究发现,低浓度的Al3+能提高花青素的含量,而高浓度的Al3+降低了花青素的含量。潘芝梅等[27]通过研究N、K、P、Fe、Ca 5种元素及其与李子表皮颜色形成的关系,发现K、Ca、P、Fe促进李子果皮花青素的合成;在荔枝果皮着色的研究中也发现,P、K、Ca元素能够促进花青素的合成,使果皮着色[28]。
高宇等[29]根据对软枣猕猴桃花青素稳定性的研究,发现K+、Na+、Ca2+对花青素生产加工中稳定性的影响没有很大差别,它们可对花青素产生保护作用,使得花青素的保存量较高。而Cu2+则会通过加速醌式碱的产生来破坏花青素的稳定性,导致其积累量降低。在黑果枸杞中也发现Cu2+能对其花青素饱和溶液有显著的褪色功效;此外其他金属离子如Al3+可明显提色花青素饱和溶液,Fe2+则使花青素溶液变成黑紫色沉积,Fe3+可直接破坏花青素的结构[30]。而赵立仪等[31]在对蓝莓花青素的研究中发现,蓝莓花青素在被酰胺化后稳定性增强,K+、Na+、Cu2+和浓度值小于0.05 mol/L 的Fe2+对其没有显著影响。
在高等教育阶段,确保家庭经济困难学生报到时可通过学校“绿色通道”直接办理入学手续,缓交学费和住宿费。入学后,所在学校将根据学生实际情况采取不同方式予以资助。
此外花青素是弱二酸,作为一种亲核试剂容易发生π堆叠相互作用,进而容易结合金属离子[32]。
1.4 水分
果树在生长过程中的花青素积累与环境中水分的含量息息相关。王虹[33]研究发现,在夏季干旱胁迫下,红叶桃中花青素含量呈现先升后降的趋势,而在秋季干旱胁迫下,花青素含量大幅度增加。此种现象也说明水分可能与温度协同作用从而对花青素的含量产生影响,且适度的干旱可以促进花青素的合成。还有研究发现水分含量过多会促进花青素苷的分解,从而促使花青素的含量升高[34-35]。
1.5 pH
花青素的色调受pH影响,其在偏碱环境下呈淡黄色,在强酸性环境下呈鲜红色,在弱酸性环境下呈深蓝色[36]。此外,花青素结构的稳定性也受pH 影响,在弱酸环境下花青素的稳定性较高;在弱碱环境下稳定性略有下降,而在强酸强碱环境下稳定性显著变差[37-38]。李媛等[39]通过对红莓稳定性和色差的变化研究发现,红莓的色泽受pH变化的影响,随着酸度的降低,红莓由红色变为黄色;且在低pH 条件下的花青素含量最高、降解量最少。而庞学群等[40]提出花青素的颜色随吸收光谱的改变而改变,且低pH能够保持花青素的稳定性并能够促进花青素的合成[41]。
以3 种被不同的糖基修饰的花青素为实验材料,研究不同酸碱环境条件对花青素稳定性的影响。结果表明,所有实验材料均在酸性条件下呈鲜红色,且较稳定[42]。
由于花青素具有在不同的pH 下会改变颜色的特性,已有一些学者对其作为酸碱指示剂的可行性展开了研究。如国外学者在对火龙果皮中花青素进行的酸碱滴定实验中发现[43],火龙果皮花青素的酸碱滴定准确度高,适合作为酸碱滴定实验中常用合成指示剂的替代品。此外亦有使用紫甘蓝与黑枸杞中花青素作为酸碱指示剂的研究[44-45]。
2 内源物质对花青素积累的调控作用
2.1 糖类
糖类在大部分植物中对花青素的积累起正调控,其在果皮颜色的变化中起重要作用。而糖能调节花青素的合成主要有2 个因素:一是糖参与了花青素的合成,通过糖基化生成稳定的花色苷;二是合成花青素的前体物质是通过莽草酸生成的,而莽草酸的生成取决于戊糖的呼吸分解,因此,花青素合成中需要大量的糖分进行呼吸分解[46-47]。而糖类在花青素合成通路的作用并不局限于物质层面,糖类也可通过对信号机制的调控作用于花青素的合成[48],如Huang 等[49]就发现杏树中一些糖可作为花青素合成中的信号分子,正调控其花青素含量。
孙倩怡等[50]研究发现蓝莓花青素的积累会随糖类浓度的增加而增加,两者呈现正相关的趋势,但只有个别糖会产生此种结果,此种现象在梨品种‘红台洋’中也有体现,该梨品种中花青素与果糖含量呈显著正相关,与蔗糖含量相关性不显著,与葡萄糖含量无相关性[51]。此外,当蓝莓和浓度较高的葡萄糖、蔗糖和果糖并存时,花青素的稳定性可得到维持,但与较低浓度的葡萄糖、蔗糖和果糖并存则加速花青素的溶解[52]。
Xiong等[53]在对杨梅的研究中也发现,‘东魁’杨梅的花青素含量较高是因为其可溶性糖的含量(包括蔗糖、葡萄糖和果糖)增加。
2.2 内源激素
生长激素常通过调整花青素合成方式中一些基因的表达来调控花青素的合成[54]。研究结果显示,生长素对花青素的合成具备两重性,即低浓度促进、高浓度抑制[55-57]。Moro 等[58]通过对红树莓果树的研究发现,在采摘后的几天内,使用IAA(3-indoleacetic acid)处理后的红树莓果实中的花青素和总酚类含量均低于对照组,且这些效应与MYB10和ANS基因的转录受到抑制有关,其影响了花青素前体修饰及花青素的上色(图1)。而Cheng 等[59]在对苹果果树的研究中发现,经生长素处理可诱导MdIAA121的表达并释放出MdARF13,而MdARF13作为花青素代谢途径的负调节因子抑制了花青素的积累。也有研究以红肉苹果为材料[57],当在其培养过程中添加生长素后,发现花青素调控基因(MdMYB10和MdbHLH3)和结构基因受0.6 mg/L浓度的2,4-D(2,4-dichlorophenoxyacetic acid)的影响而被显著抑制[57]。
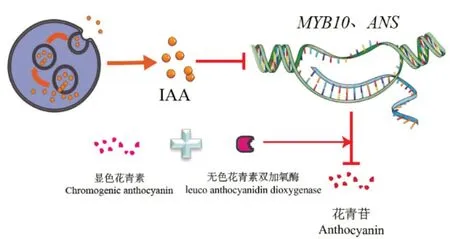
图1 IAA对花青素合成通路的影响
脱落酸对花青素的合成有正调控作用,但不同果树中脱落酸的调控机制存在差异。Qu 等[60]通过对荔枝褐变的研究发现,用ABA(abscisic acid)处理荔枝可上调参与花青素合成途径的关键酶2-氨基苯丙酸解氨酶(2-aminophenylalanine ammonia lyase,PAL)、CHS和UFGT的活性,促进了花青素的积累(图2),但赤霉素(gibberellins,GA)则起到相反的作用,如Li 等[61]在对苹果的研究中发现GA 与花青素的积累呈负相关。一些对蓝莓的研究也表明[62],外源ABA 对蓝莓果实生长并无明显影响,但其可通过加速花青素(主要是锦葵苷、飞燕草苷和矮牵牛苷)的积累来刺激果实的着色,此外ABA 也能够通过刺激枸杞中一氧化氮合酶的转录促进花青素的积累和果实着色[63];而尹金华等[64]则认为ABA 促进花青素的合成是由于促进了乙烯的合成。
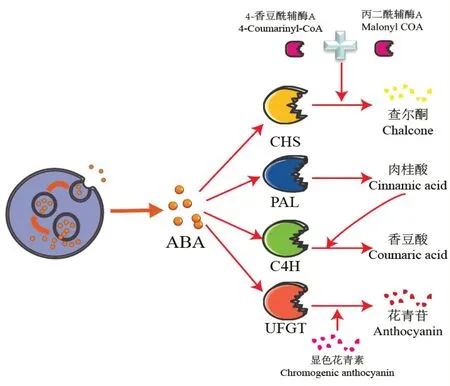
图2 ABA对花青素合成通路的调控作用
乙烯对花青素的合成有重要影响。Ni等[65]对红梨果实的研究表明,乙烯通过抑制转录因子PpMYB10和PpMYB114表达,抑制编码酶的结构基因的表达,从而降低花青素的合成。对蓝莓的研究中则发现ABA 和乙烯是糖类的重要诱导因子,协同促进果实成熟和花青素的积累与合成[66]。而乙烯与光照调控花青素合成途径也存在关联,Yu 等[67]研究发现MdACO1和MdLNC610在苹果果实和愈伤组织中存在过表达现象,MdLNC610作为促进MdACO1表达和乙烯生物合成的正调节剂,参与高光诱导花青素产生的调节。此外也有研究表明[68],乙烯可诱导转录因子MYBL2的表达,抑制花青素的合成。
细胞分裂素对花青素的积累也有促进作用。Das等[69]认为细胞分裂素可提高结构基因的表达,进而促进花青素生物合成。
茉莉酸盐与激素或植物生长调节剂具有许多共性。有研究发现[63],茉莉酸盐对果树花青素的作用主要是通过茉莉酸盐对MYB 和bHLH 转录因子家族的调控作用,诱发花青素积累。此外茉莉酸盐也会诱发梨中的丁二烯合成,进而降低花青素的转化,产生负调控作用。
3 展望
天然食用色素花青素具有来源广、用途多、安全强的特点,日益受到人们的广泛关注。此外随着近年来生物技术的发展,花青素的研究也取得了很大的进展。在应用价值方面,花青素在医学、环境、园艺、经济、作物产量等都具有显著作用;在分子结构方面,通过代谢组和转录组学等研究方法,探究了花青素的合成机制,揭示了花青素生物合成的调控网络;在影响因子方面,花青素稳定性差,初步明确了光照、温度、水分等环境因子以及内源激素对花青素的合成有一定的影响。然而花青素合成的调控机理是一个复杂的调控系统,环境因子和内源激素在调控网络中的效应和机制尚不明确,故缺乏对该调控网络全面系统的解释。因此,还需要借助分子生物学、表观遗传学等生物技术进行深入研究。此外,由于基因技术的提升,后续可以从如下方向对果树中花青素进行研究:(1)借助分子生物技术,探究各类影响因素在花青素合成中的作用模式;(2)利用基因编辑、转基因等分子技术和遗传研究方法,明确各类影响因素在花青素合成中的综合效应和机制;(3)通过添加辅色剂、花青素酰基化、微胶囊技术等技术手段,深入研究花青素在食品加工、运输和贮藏过程中,控制其稳定性的作用机理。
