盐地柽柳(Tamarix karelinii Bunge)的传粉生物学特征及传粉能力差异
2023-07-28赛丽塔娜提赛达克拜尔余润泽
赛丽塔娜提·赛达克拜尔,钟 问,余润泽
(新疆大学生命科学与技术学院/新疆生物资源基因工程重点实验室,乌鲁木齐 830046)
0 引 言
【研究意义】柽柳是柽柳科Tamaririxaceae柽柳属TamarixLinn植物的统称,广泛分布在新疆的准噶尔盆地和塔里木盆地[1],因其抗干旱、耐高温、耐盐碱、耐沙埋和耐贫瘠等特性在荒漠及半荒漠地区始终保持着优势种和建群种地位[2],在沙荒地生态环境改良中发挥了重要作用[3]。有效地保护和利用柽柳对保护与恢复新疆荒漠、半荒漠地区生态具有重要意义。柽柳在自然状态下严重依赖昆虫传粉[4],柽柳(Tamarixspp.)是新疆荒漠植被的重要组成部分,具有重要的生态和经济价值。柽柳自交难以有效完成繁殖,自然状态下严重依赖昆虫传粉,近年来,新疆柽柳分布区由于人类活动和环境恶化而明显缩小,鉴于柽柳的繁殖特性,摸清柽柳的传粉生物学特性并对该特性加以利用是保护野生柽柳资源的重要手段。研究柽柳的传粉生物学特征,分析主要传粉昆虫的传粉行为及传粉能力,对柽柳资源的保护与恢复具有实际意义。【前人研究进展】国内对柽柳属植物的传粉生物学特征及传粉昆虫的研究较为有限,但普遍指出柽柳自交不亲和且不易进行风媒传粉,主要依靠传粉昆虫进行异花传粉[5-6]。新疆作为我国柽柳集中分布并发挥重要生态价值的地区[7]。【本研究切入点】新疆地区柽柳传粉生物学的研究相较而言更加有限,仅零星报道指出膜翅目、双翅目及少数鞘翅目昆虫能够为柽柳提供传粉服务[8],对柽柳属植物的传粉生物学研究主要集中于繁殖特性[9-10]、部分传粉昆虫的行为描述[11]等,缺乏对主要传粉昆虫的确认及传粉能力比较,同时也未对柽柳属植物的传粉生物学特性进行过深入研究。针对新疆地区柽柳属植物的传粉生物学研究较为匮乏。【拟解决的关键问题】以新疆精河县艾比湖自然保护区内的野生盐地柽柳为材料,利用微距拍摄,定点连续观察记录等方法,从花部形态特征、开花动态、花粉活力与柱头可授性、花蜜含糖量昼间变化等方面探究盐地柽柳的花部综合特征;昼间与夜间观察,确定主要传粉昆虫种类,比较传粉行为及携粉能力,描述主要传粉昆虫间的传粉能力差异,研究盐地柽柳的传粉生物学特征,分析盐地柽柳的花部综合特征及主要传粉昆虫的传粉能力差异,从传粉生物学角度为野生柽柳种群的保护与恢复提供科学依据。
1 材料与方法
1.1 材 料
研究于2021年的7~9月进行,供试材料为野生盐地柽柳,地处新疆博尔塔拉蒙古自治州精河县艾比湖湿地自然保护区(新疆准噶尔盆地E 83°6′35″,N 44°37′35″),海拔231 m。气候干燥,干旱少雨,日照充足,气候变化剧烈为特征,属典型大陆性温带干旱气候,年平均气温5.4°,年平均降水量163 mm[12]。伴生的植被类型较为丰富,有梭梭Haloxylonammodendron、白梭梭Haloxylonpersicum、盐穗木Halostachyscaspica、芦苇Phragmitescommunis等。
1.2 方 法
1.2.1 花部形态与结构特征观察
于盛花期(2021年8~9月)随机选取5株盐地柽柳,从中共选取发育良好、开放状态一致且花部结构完整的20朵单花,使用便携式电子游标卡尺测量花瓣、雄蕊及花药长度等指标。
1.2.2 开花动态与花期观测
选取生长状况相似的盐地柽柳5株,对处于花蕾期的花序挂牌标记,每株标记3个花序。花开后,第1~3 d每天8:00~20:00连续观察,从第4 d至第9 d间断性观察[13],记录花朵在形态、颜色、大小、空间排列等变化。
柽柳植株有一个花序开花则视为个体开花开始,其种群开花动态按照Dafni的标准记录开花进程[14]:(1)始花期(25%以下植株开花);(2)盛花期(50%以上植株开花);(3)末花期(只有10%以下植株仍处于花期)。观察并统计植株进入始花期、盛花期以及末花期的日期和花期持续时间。
1.2.3 花粉活力与柱头可授性检测
采用TTC法测定花粉活力[15],从开花第1 d起连续检测3 d。选取少许新鲜花粉置于干净的载玻片上,加1~2滴0.5%TTC溶液,搅匀后盖上盖玻片,置入35℃恒温箱10~15 min后在低倍镜下观察,凡被染为红色的花粉活力强,淡红次之,无色者为失活或不育花粉,观察20张玻片,每片取100粒花粉,统计花粉的染色率,以检验并比较花粉活力。
采用MTT法[16]测定柱头可授性,从开花第1 d起连续检测3 d。将花的雄蕊去除后,摘取柱头置于单凹载玻片的凹陷内,加入MTT染色液,充分浸透每个柱头后,观察柱头的颜色变化。若柱头颜色呈深紫色,则显示柱头有活力,具有可授性;若柱头颜色不变,则显示柱头无活力,已失去可授性。
1.2.4 花蜜含糖量测定
柽柳的花很小,无法使用毛细管法和直接离心的方法提取花蜜直接测量含糖量,参照Malick的方法[17],在待测花序中加入等质量蒸馏水后离心,间接测定花蜜含糖量。在植株开花前选择状态一致的30个花序套袋处理,以排除昆虫等外界因素的干扰。另选取同等数量的花序不套袋作对照。于10:00~20:00,每隔2 h用镊子从花序上取下约7 g的花装入离心管中,再加入等质量蒸馏水,4 000 r/min离心8 min,获得花蜜稀释液,将离心得到的花蜜稀释液使用手持式糖度计检测含糖量,每组重复测定3次,测得的数据进行温度修正,在环境温度下读得的数值加或减温度修正值,获得准确数值。
1.2.5 传粉昆虫种类、访花频率、单花停留时间及其传粉行为
在柽柳的盛花期内,选取开花状态一致的(花序上的单花均开放)10株植株,在昼间08:00~20:00及夜间22:00~04:00,观察目标花序上的访花昆虫,记录访花昆虫种类、数量及访花行为。
采用录制视频及肉眼观察的方式,记录昼间昆虫的访花行为,将在柽柳花上活动的昆虫定义为柽柳的访花昆虫,而将接触柽柳花药后,体表携带盐地柽柳花粉,并且在访花过程中将其传送到柱头上的昆虫定义为传粉昆虫。
在观察夜间访花昆虫的行为时,使用紫外光源(λ=365 nm)或高压汞蒸汽灯(λ=380~800 nm)光源,使用观察与摄影设备的夜视模式或高感光度模式进行观察并拍摄夜间访花昆虫的行为,判断夜间访花昆虫能否完成传粉。
传粉昆虫的单花停留时间是每只传粉昆虫访问单一花朵所用的时间,即昆虫从接触一朵花到离开这朵花所需的时间;访花频率是1h内特定传粉昆虫访问花朵或花序的次数。
1.2.6 花蜜含糖量变化对传粉昆虫数量的影响
在花蜜含糖量的测定同期,统计每小时造访目标花序的传粉昆虫数量,分析花蜜含糖量变化对传粉昆虫数量的影响。
1.2.7 传粉昆虫的携粉能力比较
携粉量的计算参照罗长维[18]方法,用体视显微镜观察干燥保存的传粉昆虫标本,统计不同部位(头、胸、腹、足等)携带花粉的数量。昆虫体表被花蜜或昆虫分泌物粘结形成的花粉块中含有的,无法被转移到柽柳柱头上的花粉则不计入携粉量,每种传粉昆虫各选取10个个体样本加以统计,取其平均值。
1.3 数据处理
所有数据结果均使用IBM SPSS Statistics 26进行处理,采用单因素方差分析(One-Way ANOVA)比较不同开花天数下的花粉活力差异;采用非参数检验中的Kruskal-Wallis H检验法对不同传粉昆虫的携粉量进行差异显著性分析。所有的图表使用OriginPro 2022、GraphPad Prism9.0.2和Excel 2203制作。
2 结果与分析
2.1 盐地柽柳花部形态特征
研究表明,盐地柽柳花序长(5~15) cm,花梗长(1.02±0.16) cm,花瓣倒卵状椭圆形,长(1.8±0.09) mm,宽(0.89±0.06) mm,比花萼(0.99±0.07) mm长一半多,花瓣上部边缘向内弯曲,后侧向外隆起,深红色至深紫色,花后部分脱落,最明显的特征是花冠呈腰鼓状。裂片逐渐变宽延伸花丝基部,雄蕊5,深处花冠之外或与花冠等长,蜜腺组织退化位于花丝基部;花柱3,长圆状棍棒形。图1

图1 盐地柽柳的单花结构
盐地柽柳是夏末开花的种类,于7月下旬(7月29日)进入始花期,约1周后便进入长达约1个月的盛花期(8月7日),花期能一直持续到初秋,直到9月下旬(9月16日)结束开花,种群水平花期约58 d。
2.2 盐地柽柳开花动态
研究表明,盐地柽柳单花密集排列在大型圆锥花序上,于07:30左右从花序基部向顶端次第开放。开花前花冠呈腰鼓状,雌蕊与雄蕊被包在花瓣里,开花后花丝随花瓣展开充分伸展至花瓣周围,花柱完全露出,在11:00左右大量花瓣都已展开,单花花期约2~3 d。盐地柽柳开花后散粉时间较长,约8 h,柱头在开花后1~2 h内活力最强。图2,表1

A:处于花苞期的花序;B:开花前花瓣呈腰鼓状;C:盛花期;D:花末期,花药已脱落,花柱皱缩,少数花瓣宿存;E:结种期;F:夜间花序开放状态,花瓣略闭合
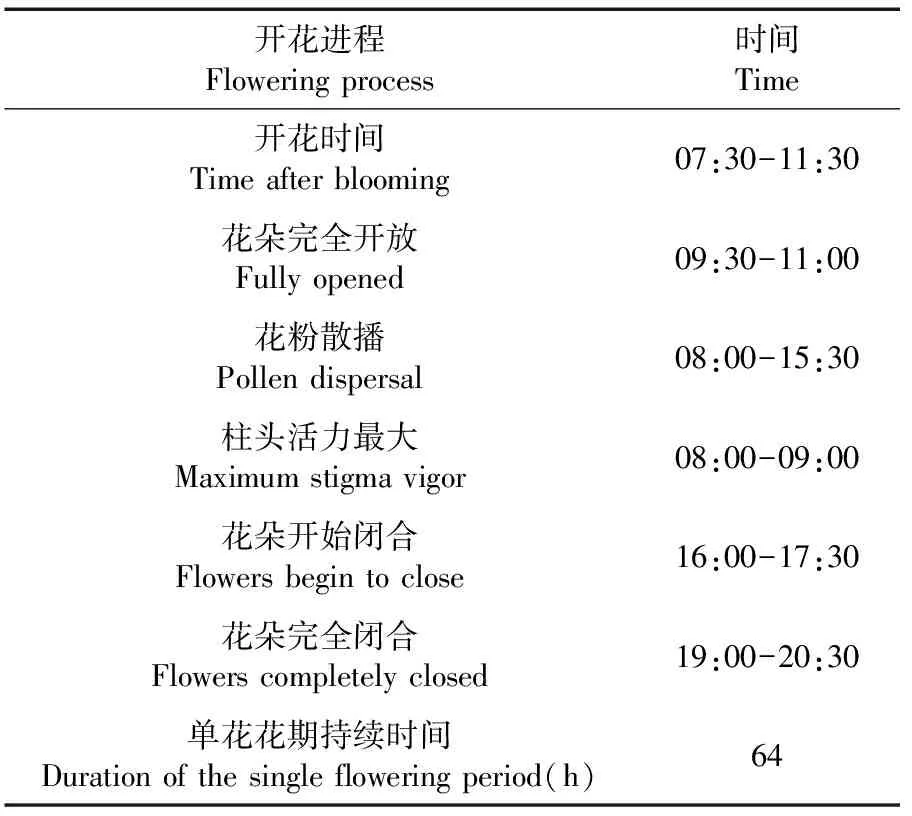
表1 盐地柽柳的单花开花时期
2.3 盐地柽柳花粉活力变化
研究表明,盐地柽柳具有活力的花粉均能被TTC溶液染成红色或浅红色,而失活的花粉则无法被染色。盐地柽柳开花1~3 d的花粉活力差异显著(P<0.05),随着开花天数的延长花粉活力逐渐降低,并于开花第1 d花粉活力达到最高(84.4%),开花后第3 d花粉活力最低(35.85%)。图3
2.4 自然状态下盐地柽柳柱头可授性变化
研究表明,盐地柽柳在单花开花期(开花后1~3 d)柱头可授性随着开花时间的延长而逐渐下降。开花当天柱头均具有100%的极强可授性,柱头形态饱满且直立,与染色液相接触后颜色变化明显;开花2 d起柱头开始轻微皱缩,或出现褐色斑点,与染色液达到反应也较为缓慢,柱头活性(73.33%)低于开花第1 d,但未检测出无可授性的柱头;开花第3 d,柱头发生明显的形态变化,皱缩变小,柱头活性(23.33%)降低幅度较大,且可检测出多个失活柱头。表2

注:图中“*”表示P<0.05,差异显著

表2 盐地柽柳在开花1~3 d的柱头可授性变化
2.5 盐地柽柳花蜜含糖量的日变化规律
研究表明,套袋状态下的花蜜含糖量,显示了不受昆虫觅食行为活动干扰的,植株花蜜含糖量。开花当天,盐地柽柳植株套袋处理的花序的花蜜含糖量首先呈缓慢上升趋势,在10:00~12:00含糖量最低(2.34%),于18:00~20:00达到当日峰值(4.34%)。
未套袋状态下的花蜜含糖量,显示了花朵在自然状态下,被不同昆虫造访后花蜜的含糖量变化。盐地柽柳在自然状态下的花蜜含糖量变化总体呈先上升后下降趋势,于14:00~16:00达到一天中的最高值(3.84%),花序在自然状态下花蜜含糖量日变化更加明显。图4

图4 套袋与自然状态下的花蜜含糖量
2.6 盐地柽柳的传粉昆虫种类及其传粉行为
研究表明,盐地柽柳的昼间访花昆虫隶属于膜翅目、双翅目及鞘翅目,其中双翅目昆虫个体数量最多(44.94%),膜翅目昆虫次之(43%),鞘翅目昆虫个体数量最少(5.30%)。夜间观察到的访花昆虫隶属于鳞翅目夜蛾科,只是长时间停留在花上休息,未见传粉行为。盐地柽柳的主要传粉昆虫共2目4科5种,其中意大利蜂Apismellifera(蜜蜂科Apidae)、稀刻淡脉隧蜂Lasioglossumalbescens(隧蜂科Halictidae)分别占膜翅目访花昆虫个体总数的35.96%和26.97%,长尾管蚜蝇Eristalistenax(食蚜蝇科Syrphidae)、大灰优食蚜蝇Eupeodescorollae(食蚜蝇科Syrphidae)及丝光绿蝇Luciliasericata(丽蝇科Calliphoridae)分别占双翅目访花昆虫个体总数的20.43%、17.20%及15.05%,是盐地柽柳主要的传粉昆虫。图5

图5 盐地柽柳访花昆虫类群及
意大利蜂一般从花序基部爬行至花序顶端并不断吸食花蜜,在此过程中,吸食花蜜的口器及携带大量花粉的附肢和胸腹部喙持续性触碰柱头和花药,从而完成传粉;稀刻淡脉隧蜂在访花时通常径直飞向花瓣并迅速地用前足收集花粉,并通过中足将花粉传递到后足的花粉篮内,在取食花蜜的过程中其口器、头部正面及腹部和附肢的体毛上长可以附着大量的花粉,在造访下一朵花时与花的柱头接触,完成授粉;长尾管蚜蝇访花前常在空中短时间悬停,然后飞向花瓣,访问同一花序不同花朵时常采用爬行而非飞行的方式,通过不断调整身体位置,将口器探入单花花冠内取食开裂的花药表面的花粉并吸食花蜜,在取食过程中其舐吸式口器、复眼、足以及胸腹部的体毛上附着的花粉会有机会接触柱头,完成传粉过程;大灰优食蚜蝇的访花行为与长尾管蚜蝇的相似,但其后足较长,访花时腹部会与花保持一定的距离,故胸腹部与花的雌雄蕊很少有接触,只有头部和口器、胸足等附肢能与花药和柱头相触碰,完成传粉;丝光绿蝇的访花活动十分频繁,访花时会在花序上来回爬动,口器和部分头部伸进花冠舐吸花蜜,在这个过程中胸部腹面及腹部腹面、部分附肢及头部都会粘附花粉,在访问下一朵花时接触柱头完成传粉。图6
2.7 开花当天主要传粉昆虫数量与花蜜含糖量变化间的关系
研究表明,盐地柽柳开花第1 d,主要传粉昆虫数量呈先增后减少的趋势,在14:00~16:00达到最高值,该时间段的花蜜含糖量也达到最高值。图7

注:A:意大利蜂;B:稀刻淡脉隧蜂;C:长尾管蚜蝇;D:大灰优食蚜蝇;E:丝光绿蝇
2.8 主要传粉昆虫的访花频率及单花停留时间比较
研究表明,不同传粉昆虫的访花习性不同,表现在其的单花停留时间和访花频率有显著差异(P<0.05)。主要传粉昆虫在一朵花上停留访花时间的长短依次为意大利蜂>稀刻淡脉隧蜂>长尾管蚜蝇>丝光绿蝇>大灰优食蚜蝇。其中意大利蜂的单花停留时间最长,平均每朵花停留(25.95±3.20) s,大灰优食蚜蝇单花停留时间最短,为(13.25±3.40) s。
5种传粉昆虫访花频率的高低依次为稀刻淡脉隧蜂>意大利蜂>丝光绿蝇>长尾管蚜蝇>大灰优食蚜蝇,稀刻淡脉隧蜂的访花频率最高,为(3.89±0.60)次/h,大灰优食蚜蝇的访花频率最低为(1.89±0.93)次/h。图8
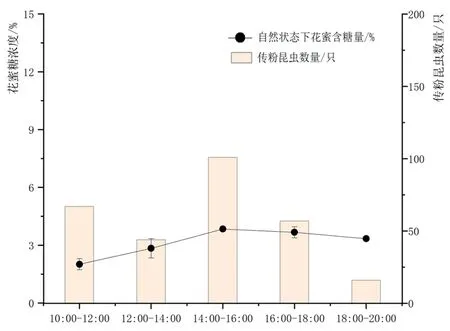
图7 盐地柽柳花蜜含糖量与传粉

图8 主要传粉昆虫的单花停留时间与访花频率
2.9 主要传粉昆虫的携粉能力比较
研究表明,5种主要传粉昆虫体表总携粉量差异显著(P<0.05),总携粉量的高低依次为意大利蜂(490.1±13.55)粒>长尾管蚜蝇(278.5±7.03)粒>稀刻淡脉隧蜂(195.2±9.41)粒>丝光绿蝇(96.6±8.21)粒>大灰优食蚜蝇(22.8±4.66)粒。不同传粉昆虫主要的携粉部位有所不同,意大利蜂、长尾管蚜蝇及丝光绿蝇的主要携粉部位位于胸部与足部,稀刻淡脉隧蜂体表约50%的花粉都黏附在腹部,腹部是主要的携粉部位,大灰优食蚜蝇的携粉量极低,体表仅检测到少量花粉,主要集中在头部与胸部。图9

图9 主要传粉昆虫体表不同部位携粉量
3 讨 论
泛化型的虫媒植物的花部综合特征能够吸引和适应多样化的传粉昆虫,保障传粉效率和繁殖成功率[19]。植物的花部综合特征(floral syndrome)为花部构成(floral design)及开花式样(floral display),前者一般包括花的结构、颜色、气味、传粉(花粉、花蜜)等,是花在单花水平上对传粉昆虫的吸引;后者则是植物在某一阶段花朵开放的数量及其在花序上的结构排列,体现花在种群水平上对传粉昆虫的吸引[20]。盐地柽柳的花药与柱头外露于花冠外的特征在一定程度上增大了传粉者与花生殖部位的接触机率,表现出了对虫媒传粉的适应。盐地柽柳的花序长5~15 cm,开花时多个单花密集排列组成大型的圆锥花序,有助于昆虫在花序上来回爬行以便取食。盛花期持续的时间较长,占开花进程的半数以上,较长的花期为其充分传粉打下了一定的基础,单花花期为3 d,保证了处于盛花期时,植株上每天都有大量的花开放,植物的这种开花模式是为了应对生长环境或传粉者稀缺所形成的特殊适应性机制[21-22]。
开花当天,经套袋处理后,花蜜含糖量在一天中呈上升趋势,并于18:00~20:00达到最高值,这很有可能说明花蜜体积会随着时间的增长逐渐积累,使得相应时间段的花蜜含糖量较高。自然状态下花序含糖量变化结果表明,14:00~16:00自然状态下花序的花蜜含糖量达到最高值,相应的该时间段内访花者的活动也最为频繁,昆虫的访花活动与盐地柽柳花蜜含糖量变化有较高的相关性,昆虫更倾向于去访问花蜜含糖量较高的花朵。花蜜含糖量在一定时间范围内产生波动的原因可能跟环境因子的变化相关,随着环境湿度的降低、环境温度的升高,花蜜中的水分在不断蒸发,很有可能是植物吸引传粉昆虫访花的一种策略。
虫媒植物的传粉过程中,传粉昆虫起着关键作用,不同传粉昆虫之间因访花行为与访花频率的不同,对植物产生的传粉效应也不同。昆虫的传粉效力与其传粉数量及传粉质量密切相关[23-24],传粉效力(即花粉携带转运能)强调传粉能力,是由多个指标来反映的,通常需要对传粉数量和传粉质量同时评估,传粉数量主要取决于访花频率,而传粉质量则取决于传粉者大小、访花行为等[25-26]。研究中,膜翅目与双翅目昆虫均能为其传粉,其中意大利蜂、稀刻淡脉隧蜂、长尾管蚜蝇、大灰优食蚜蝇及丝光绿蝇个体数量占比大、携带有盐地柽柳的花粉并观察到了传粉行为,因此是盐地柽柳较为稳定的传粉昆虫种类。可能与几种类群昆虫的习性相关,如易大面积扩散繁殖、种群数量庞大,其能与荒漠生境炎热、干旱的气候相适应。
稀刻淡脉隧蜂的访花频率最高,在同一时间段里,稀刻淡脉隧蜂相较于其它4种传粉者能访问更多花的可能性较大,这在一定程度上提高了植物的授粉机会。昆虫携粉能力的高低也影响传粉能力,携粉量高的传粉昆虫通常可以带给柱头更多的花粉,即意味着传粉的成功率更高[27]。研究中,5种主要传粉昆虫的携粉量差异显著,其中意大利蜂的携粉量最高,可能与这几种传粉者的形态结构相关,体表越光滑、体毛分布较少且不密集等,使得花粉难以在体表附着,携带的花粉量就越少,意味着携粉能力就越弱。
4 结 论
盐地柽柳花两性,花药与柱头外露于花冠外,分泌花蜜,具有虫媒传粉的花部综合特征;种群花期约58 d,单花花期为3 d,单花花期内花粉和柱头均具有活力,其中开花当日的花粉活力和柱头可授性最高;意大利蜂Apismellifera、稀刻淡脉隧蜂Lasioglossumalbescens、长尾管蚜蝇Eristalistenax、大灰优食蚜蝇Eupeodescorollae及丝光绿蝇Luciliasericata为主要传粉昆虫;主要传粉昆虫的访花高峰与花蜜含糖量最高的时间段一致;意大利蜂的个体数量最多、单花停留时间最长、携粉量最高,是盐地柽柳较为高效的传粉者。
