曹妃甸湿地自然保护区植物多样性研究*
2023-07-24胡爱双郭文静邢春强马旺孙宇丁冯洁张薇
胡爱双 ,郭文静 ,邢春强 ,马旺 ,孙宇** ,丁冯洁 ,张薇
(1.河北省农林科学院滨海农业研究所/河北省盐碱地绿化技术创新中心 唐山 063299;2.中国林业科学研究院生态保护与修复研究所 北京 100091;3.河北省水文工程地质勘查院 石家庄 050021)
陆地、海洋和湿地是世界上最大的3 个生态系统,包含草原、森林、荒漠和江河湖泊等多个生态子系统[1]。沿海湿地生态系统属于湿地生态系统的一种,介于海陆交汇地带,同时受陆地和海洋双重作用的影响[2],具有独特的生态系统结构和复杂多样的生态景观,从而造就了其与众不同的生物多样性和环境敏感性。对沿海湿地生态系统的功能研究显示,湿地在物质循环、能量流动、信息传递等过程中发挥着重要作用,为近海岸人类提供海岸防护、净化水质、调节气候、保护淡水资源、航运、教育和科研等多重保障和服务,是地球上生产力最高、生态服务价值最大的生态系统[3]。因此,调查研究、保护、修复滨海湿地具有重要意义。
生物多样性是衡量湿地生态系统结构和生态系统稳定性的重要指标,集物种、遗传、功能和生态系统多样性于一体[4]。湿地植物多样性是该地区物种多样性的重要组成部分,对维持生态系统功能和服务、指示生态环境质量状况、湿地保护及修复有至关重要的作用[5-6]。湿地植物多样性受气候变化、土壤理化性质、微生物种类和水环境质量等因素的综合影响[7-10]。其中,土壤作为湿地植物生长的空间载体,为植物提供水分和养分,对湿地植物的多样性有较为明显的影响[11]。如多项研究显示土壤理化性质优越的地方,植物生长比较茂盛,生态结构比较稳定[12-13]。同时,土壤理化性质的恶化也可能会对植物生长产生不利影响,如湿地表层土壤盐分的积累可能会降低植株微量元素的吸收,抑制植物生化酶的活性,最终造成植物多样性降低[10]。王杰等[14]对滦河河口地区植物的多样性研究结果显示,该区域植物的多样性受到了土壤盐分的影响。李艳红等[15]对艾比湖湿地植物的多样性研究结果显示,植物的多样性也与土壤全盐含量显著相关。此外,植物对土壤理化性质也会产生一定的作用,如植被覆盖能拦截降水、减少地表径流和减少土壤侵蚀[16];植物的枯枝落叶腐烂后可以促进土壤微生物的发展使土壤更具活力[17];豆科(Fabaceae)植物根部共生的根瘤菌可以固氮,改善土壤结构,保持土壤肥力[18]。土壤环境和植物多样性的相互作用和良性循环共同维持区域生态系统的稳定和可持续发展,反映了植物群落的适应能力和生态环境的可调节机制[19]。因此,为了科学管理湿地生态系统,有必要对湿地植物的多样性进行调查,并对其与土壤理化性质的关系进行研究。
曹妃甸湿地生态系统处于陆地生态系统和海洋生态系统的过渡地带,是具有国际意义的沿海湿地生态敏感区。近些年来随着沿海经济的高速发展,受气候和人为因素的影响[2],曹妃甸湿地普遍出现了鱼塘遍布、土壤返盐和板结,度假村和酒店林立等现象,对植物多样性保护和生态系统功能保障带来了巨大挑战。目前对于曹妃甸湿地的研究主要集中在湿地生态服务价值空间分异[20]、生态健康评价[21]、旅游产业发展[22]等方面。而关于土壤环境因子与植物多样性相关关系的研究较少。本文以曹妃甸湿地和鸟类省级自然保护区为研究区域,通过对该湿地的植物空间分布、土壤理化性质进行实地调查和分析,探究湿地植物资源现状、土壤养分和盐分空间分布特征并分析植物群落与土壤环境因子的关系,以期为湿地生态系统修复、植物多样性保护等湿地综合系统的科学管理提供理论支撑。
1 研究区概况与研究方法
1.1 研究区概况
曹妃甸湿地和鸟类省级自然保护区是2005 年9 月,经河北省人民政府批准建立的省级自然保护区。位于河北省唐山市曹妃甸区南部(118°15′~118°23′E,39°09′~39°14′N),区域平均海拔2.5 m,属温带半湿润气候,总面积11 064 hm2。其中核心区面积3504 hm2,占全区31.7%;缓冲区面积1503 hm2,占全区13.6%;实验区面积6057 hm2,占全区54.7%。保护区北侧距沿海公路南侧800 m,南与南堡盐场相邻,东靠青林公路以西1500 m 处的斗渠,西以三排干为界(图1)。保护区属温带半湿润气候区,四季分明,年平均气温11.2 ℃,年降水量648.1 mm。保护区生物多样性十分丰富,以海淡水湿地生态系统和国家一、二级鸟类为主要保护对象。
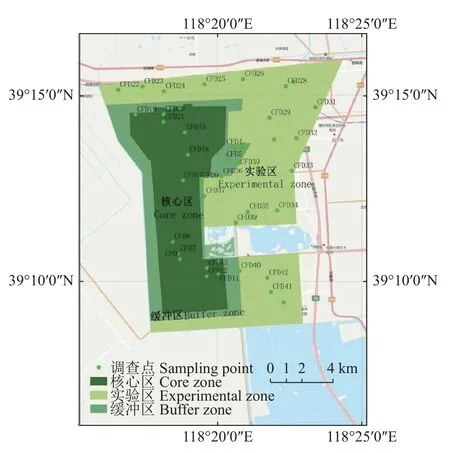
图1 研究区位置及样方分布示意图Fig.1 Location and sampling points distribution of the study area
1.2 研究方法
1.2.1 植被调查
于2021 年8 月开展调查,主要以双龙河为主线,采用生态学样地调查方法[14]对研究区植物群落结构及土壤化学性质进行调查,沿河岸、海岸、垂直于河岸、海岸方向选取具有典型群落特征的区域设置样地(10 m×10 m) 42 个。若样地内存在乔木,则算作一个乔木样方;对于有灌木存在的样地,随机选取5 m×5 m 样方1~2 个;草本植物样地随机选取1 m×1 m样方1~3 个。共计样方93 个,每个样方采用GPS 定位。现场鉴定样方中的植物物种,记录个体数、胸径、高度、冠幅等指标及周边环境特征。
1.2.2 土壤样品采集与分析
沿样方对角线方向选取5 个取样点,用土钻分层采集0~20 cm 和20~40 cm 的土样分别混合后装入自封袋中带回实验室,自然风干、去杂、研磨处理后进行土壤化学性质测定。土壤全盐含量采用电导法(水土比5∶1 浸提)测定,速效氮采用碱解扩散法测定,速效磷采用NaHCO3浸提-钼锑抗比色法测定,速效钾采用NH4Ac 浸提-火焰光度法测定,有机质采用重铬酸钾容量法-外加热法测定[23]。
1.2.3 植物多样性指标
文中涉及指标主要包括Margalef 指数(R)、Shannon-Wiener 指数(H)、Simpson 指数(D)和Pielou均匀度指数(E)[24]。R值反映群落物种丰富度,表示生物群聚中种类丰富度程度;H值越大,群落所含的信息量越大,代表了群落的复杂程度越高;D值越大,表示群落中种数越多,各种个体分配越均匀,物种多样性程度越高;E值表示群落物种分布的均匀度。相关计算公式如下:
式中:S为样方中物种总数,N为样方中记录的个体总数,Ni为样方中第i个物种的个体数。
运用SPSS 22.0 软件对植被多样性指数和土壤环境因子进行相关性分析,运用ArcGIS 10.2 空间分析功能中反距离权重差值(IDW)方法对土壤盐分和养分进行空间分析,生成空间分布图。
2 结果与分析
2.1 研究区植物区系特征
2.1.1 植物区系基本组成
统计分析结果表明:研究区内种子植物共有23 科、47 属、54 种。被子植物有22 科、46 属、53 种,分别占科属种的95.7%、97.9%、98.1%,其中双子叶植物有20 科、44 属、51 种,分别占科属种的87.0%、93.6%、94.4%,单子叶植物有2 科、2 属、2 种,分别占科属种的8.7%、4.3%、3.7%。裸子植物只有1 科、1 属、1 种,被子植物在区系中占绝对优势。研究区植物的生活型以草本植物为主(39 种),如草木樨(Melilotus officinalis)、翅果菊(Lactuca indica)、盐地碱蓬(Suaeda salsa)等,共占研究区植物总种数的72.2%;灌木类6 种,如白刺(Nitraria tangutorum)、紫穗槐(Amorpha fruticosa)、柽柳(Tamarix chinensis)等,占研究区植物总种数的11.1%;乔木类10 种,主要为人工栽植的树种,如垂柳(Salix babylonica)、桑(Morus alba)、槐(Styphnolobium japonicum)等,占研究区植物总种数的18.5%;草质藤本只有鹅绒藤(Cynanchum chinense) 1 种。研究区植物生态水分类型以中生植物为主(46 种),如白茅(Imperata cylindrica)、萹 蓄(Polygonum aviculare)、地肤(Kochia scoparia)等,占植物总种数的85.2%;湿生植物有5 种,如稗(Echinochloa crus-galli)、碱蓬(Suaeda glauca)、芦苇(Phragmites australis)等,占植物总种数的9.3%;旱生只有白刺、中华补血草(Limonium sinense)、二色补血草(Limonium bicolor)3 种,占植物总种数的5.6%。中生生态类型中大部分为中中生植物,占比为75.9%,其次是湿中生和旱中生,占比分别为5.6%和3.7%。
2.1.2 区系数量结构
优势科是指在植物区系中种类众多,且在植物群落中起着建群作用的科[25]。以科所包含种数为依据进行统计分析[26],研究区前4 科所包含种数大于平均值3,在区系总种数占比为59.3%,菊科(Asteraceae)、禾本科(Poaceae)、藜科(Chenopodiaceae)和豆科是该区系的优势科(表1)。在植物区系中种类众多,且在植物群落中起到建群的作用,如藜科的碱蓬、地肤、盐地碱蓬群落,禾本科的芦苇群落等。表征科是某科在该区系中的种数与该科在全世界分布的种数对比,比值越高,代表性越强[27]。将研究区植物科(除单种科、仅含有 1 种的科)与所对应的世界植物区系做对比,比值范围为 0.03%~0.50%,平均值为0.21%。由此确定藜科、白花丹科(Plumbaginaceae)、杨柳科(Salicaceae)是该区系的表征科(表1)。

表1 研究区植物优势科和表征科Table 1 Dominant families and representative families of vegetation in the study area
与科的统计相似对属一级进行统计分析,结果表明藜属(Chenopodium)、莴苣属(Lactuca)、碱蓬属(Suaeda)、蒿属(Artemisia)、补血草属(Limonium)是区系的优势属(表2),莴苣属和碱蓬属是该区系的表征属。

表2 研究区植物优势属及表征属Table 2 Dominant genera and representative genera of vegetation in the study area
2.1.3 区系地理成分分析
植物区系是某一地区或某一时期所有植物种类的总称,是植物界在特定自然地理环境条件下发展、演化的结果,可在一定程度上反映植被特征和地理环境变化[28]。以吴征镒等[29]对植物科、属区系地理成分的划分为依据,对研究区植物区系地理成分统计分析。结果表明:研究区区系地理成分在科一级以世界分布为主,有13 科,占区系总科数56.5%;温带分布有3 科,占区系总科数13.0%;热带分布型有6 科,占区系总科数的26.1%;古地中海分布只有1 科,占区系总科数4.3% (表3)。在属一级划分为5个分布大类、8 个分布型和6 个分布变型。其中世界分布型有11 属,这类分布型植物没有明确的起源中心,无法表现出区系的起源与演替(故不做统计),如蒿属植物几乎遍布全国,常组成植物群落,为荒地、草地、部分湿地的建群种、优势种。多生长在荒坡、旷野及路旁,少数种也分布到海边滩地。鬼针草属植物几乎遍布全国各地,多为荒野杂草等。热带分布类以泛热带分布型为主,有6 属,占非世界分布属的16.7%,如补血草属、狗尾草属、马齿苋属等。温带分布类以泛温带为主,有9 属,占非世界分布属的25.0%,如白茅属、萹蓄属、地肤属等。亚洲分布类以东亚分布型及其变型为主,总计5 属,占非世界分布属的13.9%,如侧柏属、葎草属、茜草属等(表4)。

表3 研究区植物科级区系的类型Table 3 Types of family level flora in the study area
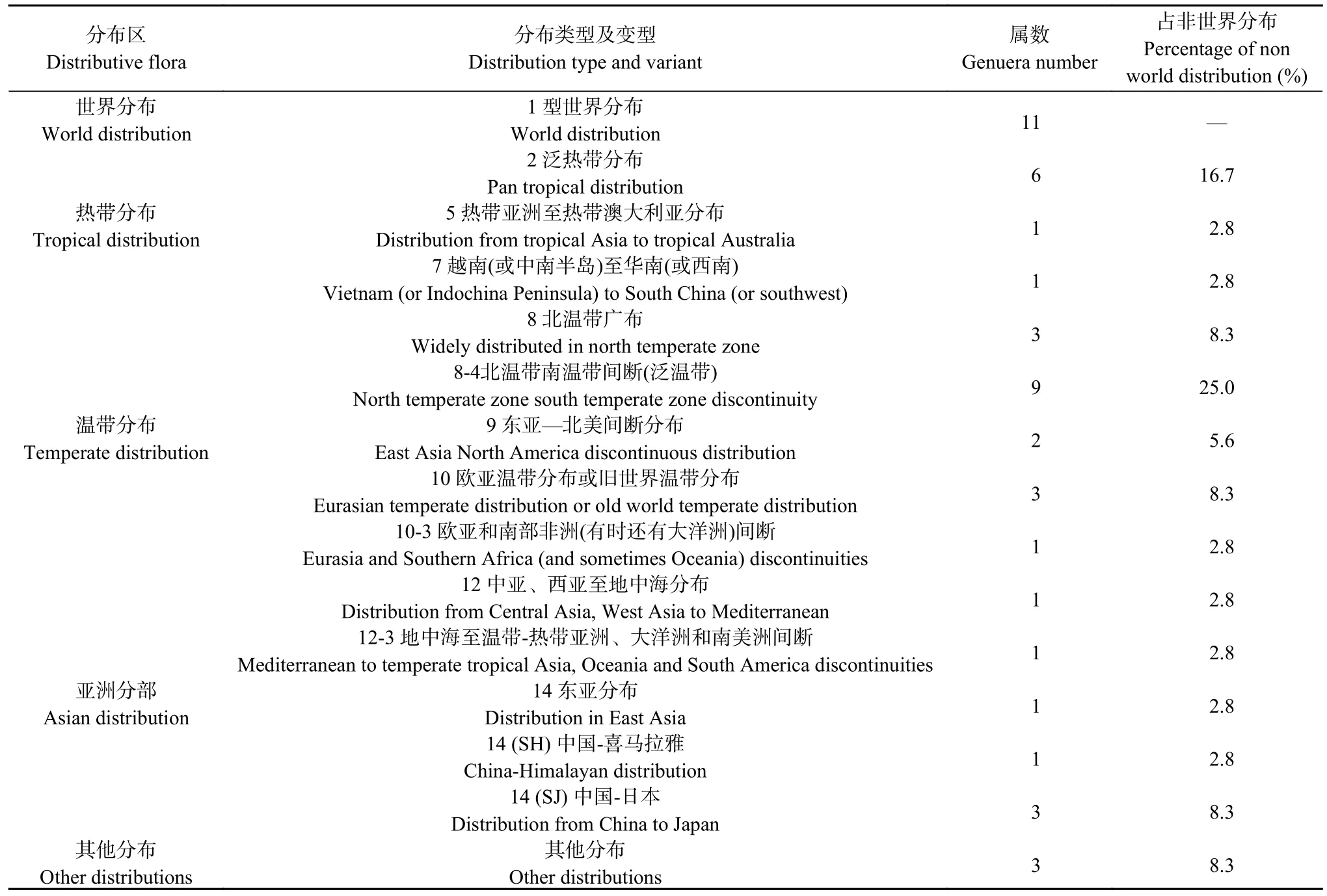
表4 研究区植物属一级区系的类型Table 4 Types of genus level flora in the study area
2.2 研究区植物群落多样性分析
2.2.1 乔灌层群落多样性分析
研究区乔木灌木群落多样性单一,灌木主要是白刺和柽柳,为海滨、滩头、潮湿盐碱地等区域的常见种;乔木层主要是刺槐(Robinia pseudoacacia)、榆树(Ulmus pumila)等栽培种。
2.2.2 草本层群落多样性分析
研究区植被以草本为主,依据实地调查结果及相关指数计算,研究区群落类型有碱蓬、地肤、黄花蒿(Artemisia annua)、盐地碱蓬、芦苇、葎草(Humulus scandens)、三棱草(Carex phacota)、婆婆刺(Bidens pilosa)、茵陈蒿(Artemisia capillaris) 9 种(表5)。群落盖度范围为59.6%~99.9%,最高值是葎草群落,最低值是碱蓬群落;群落植物种数为3~13,最高值是葎草群落,最低值是碱蓬群落;多度和范围为46~222,最高值是三棱草群落,最低值是地肤群落;H值范围为0.77~2.18,最高值是葎草群落,最低值是碱蓬群落;D值范围为0.44~0.86,最高值是茵陈蒿群落,最低值是芦苇群落;E值范围为0.53~0.92,最高值是茵陈蒿群落,最低值是芦苇群落;R范围为1.78~9.64,最高值是葎草群落,最低值是碱蓬群落。研究区芦苇和碱蓬群落的多样性较差,而葎草、茵陈蒿群落结构更具有稳定性。
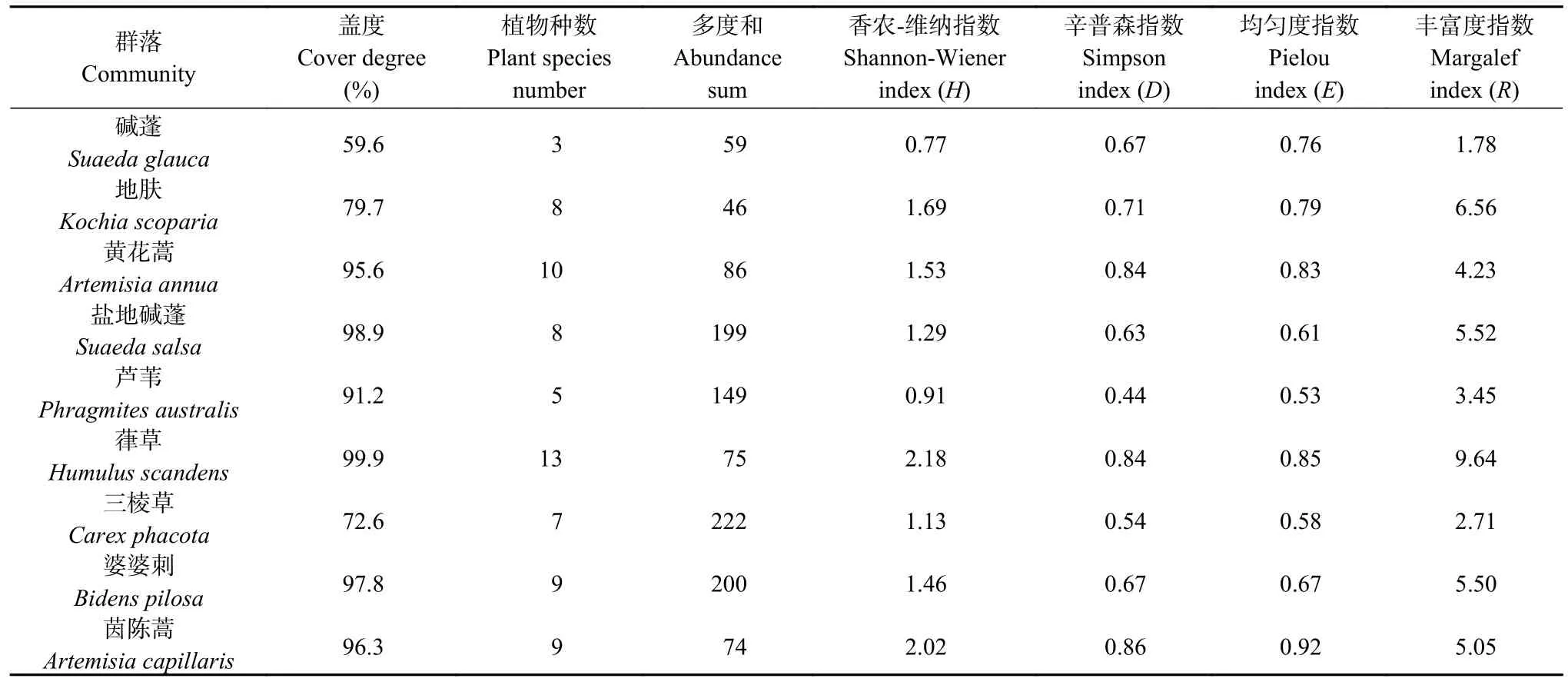
表5 研究区主要植物群落及其主要特征Table 5 Main plant communities and their main characteristics in the study area
2.3 土壤含盐量和土壤养分条件分析
2.3.1 土壤盐分和养分空间分布特征
根据地理数据统计分析,研究区土壤盐分空间分布如图2 所示,0~20 cm 和20~40 cm 土壤盐分在水平空间上分布较为相似,主要呈现沿西南向东北递减的趋势。重度盐渍土(>5 g∙kg−1)主要分布在研究区的西南部地区,该区西部和南部地区临近晒盐池和渤海,受海水入渗共同影响导致西南部地区土壤盐分含量较高。中度盐渍土(3~5 g∙kg−1)零星分布于中部和东北部地区,主要为临近池塘、鱼池和水系地区。轻度盐渍土和非盐渍土主要位于中部和东北部地区,离海岸线较远,土壤盐分含量较低。0~20 cm、20~40 cm 土层非盐渍土样地数量占全部样地的51%和37%,轻度盐渍土分别占19%和35%,中度盐渍土分别占12%和7%,重度盐渍化样地数量分别占18%和21%。在垂直空间分布上,随着土壤深度增加,轻度盐渍土和重度盐渍土土壤面积呈增加趋势,非盐渍土和中度盐渍土面积呈减少趋势,同时土壤盐分含量降低。
对研究区42 块样地中土壤速效氮、速效磷、速效钾和有机质等土壤养分含量进行空间插值,结果如图3 所示,土壤速效氮含量为5.26~89.24 mg∙kg−1,不同土层深度速效氮含量空间分布较为相似,高值区主要位于西北和中部地区,其余地区速效氮含量较低。垂直空间分布上,速效氮高值面积在减少,低值面积在增加,同时随着土层深度增加速效氮含量呈减少趋势。土壤速效磷含量为5.94~73.87 mg∙kg−1,在空间分布上,0~20 cm 土层深度土壤速效磷含量高值区主要位于研究区西北部和中部部分地区,20~40 cm 土壤速效磷高值区主要位于西北和西南部地区。在垂直空间分布上,随着土层深度增加,速效磷高值区在增加,低值区在减少,速效磷含量整体在减少。土壤速效钾含量为10.62~109.82 mg∙kg−1,空间分布上,0~20 cm 土壤速效钾高值区主要分布在西南部地区和中部地区,20~40 cm 土壤速效钾高值区主要分布在西南部地区。随着土层深度增加,速效钾高值区范围在减少,低值区范围在增加,速效钾含量整体减少幅度较小。土壤有机质含量为1.13~28.98 g∙kg−1,空间分布上,0~20 cm 土壤有机质在西北部、东北部和东南部地区较高,在西南部地区和北部地区较低;20~40 cm 土壤有机质分布较为集中,高值区主要分布在西南部和北部的部分地区。随着土层深度增加,土壤有机质高值区范围在减少,低值区在增大,同时土壤有机质整体含量呈减少趋势。
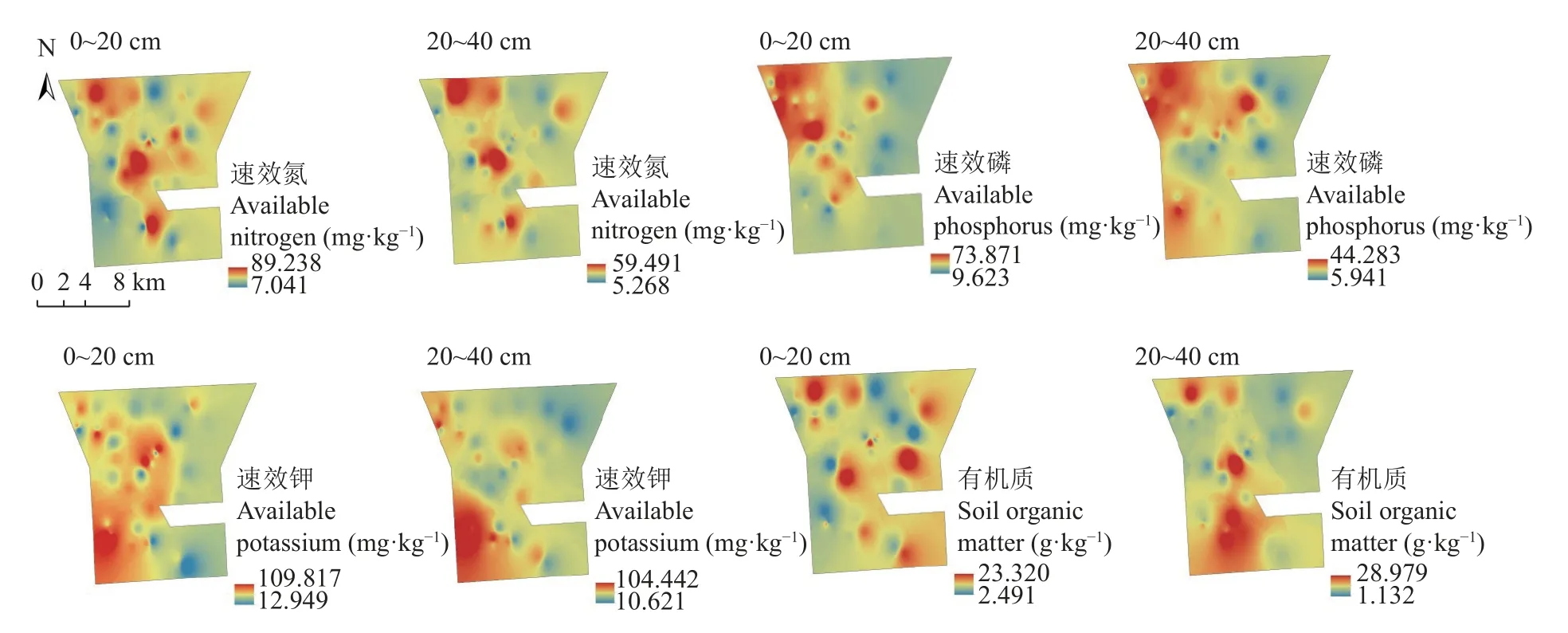
图3 研究区0~20 cm 和20~40 cm 土层土壤养分空间分布Fig.3 Spatial distribution of soil nutrients of 0−20 cm and 20−40 cm layers in the study area
2.3.2 盐分梯度及土壤因子特征
由于土壤盐渍化空间分布呈现由西南向东北减少的趋势,因此选取距离西南角0~5 km、5~10 km和10~15 km 处作为样带1、样带2 和样带3 区域,其中样带1、样带2 和样带3 区域分别包括8 个、17 个和17 个样地。不同样带土壤盐渍化程度及养分含量如表6 所示。0~20 cm 土壤,不同样带间的土壤盐分和养分差异均不显著。20~40 cm 土壤,样带1 土壤盐分显著高于样带2 和样带3。土壤速效氮和速效钾含量在不同样带和土层间均不显著。0~20 cm和20~40 cm 土壤速效磷和有机质含量不同样带间均不显著。样带2 的0~20 cm 土壤速效磷含量显著高于20~40 cm。

表6 研究区不同样带的土壤盐渍化程度及养分含量Table 6 Salinization degrees and nutrients contents of soil in different zones in the study area
2.4 盐碱区植被多样性特征与土壤因子的关系
对不同区域不同土层土壤养分和植被多样性指数进行相关性分析,结果如表7 所示。整个区域0~20 cm 土壤的盐分与植被的Margalef 指数(R)、Shannon-Wiener 指数(H)、Simpson 指数(D)和Pielou指数(E)呈负相关关系。

表7 曹妃甸湿地植物群落多样性与土壤环境因子的相关系数Table 7 Correlation coefficient between plant community diversity and soil environmental factors in Caofeidian wetland
对于样带1,20~40 cm 土壤盐分含量与R多样性指数呈显著负相关。速效磷和速效钾含量与各植物多样性指数主要呈负相关关系。速效氮含量与各植物多样性指数呈正相关关系。土壤有机质含量与R和H呈正相关,与D和E呈负相关关系。不同深度土壤速效氮与H呈显著正相关,20~40 cm 速效氮与D呈显著正相关,20~40 cm 速效钾与R呈显著负相关,不同深度土壤有机质与R呈显著正相关。
对于样带2,土壤盐分和土壤养分含量与植物多样性指数相关系数为−0.274~0.42,未检测到显著相关性。对于样带3,速效氮、速效磷和速效钾与各植物多样性指数整体呈正相关关系。其中,20~40 cm速效氮含量与H达到显著正相关,不同深度土壤速效磷含量与R和H呈显著正相关,20~40 cm 土壤有机质含量与R和H指数呈显著正相关。与0~20 cm相比,20~40 cm 土壤养分与各植物多样性指数相关性整体较高。
3 讨论
研究区湿地临海,部分土壤盐碱化严重,同时存在裸露地面和废弃地,导致植物种类较少,仅有23科47 属54 种,被子植物占绝对优势,与渤海湾周边湿地调查相一致[14]。四季分明的温带气候及较为复杂的生态环境使得植物生活型以草本植物为主,生态水分类型以中生植物为主。优势科是菊科、禾本科、藜科和豆科,表征科是藜科、白花丹科、杨柳科。特别是藜科,既有灰绿藜(Oxybasis glauca)、猪毛菜(Salsola collina)等该地的常见种,也有盐地碱蓬、碱蓬等海边、盐碱地常见的盐生植物。
研究区植物区系相对复杂,受地理位置影响,以温带分布为主,但在科及属水平均出现热带分布,表明该区与热带分布有一定联系,同时存在一定古地中海成分。这与前人对渤海湾湿地植物区系的研究结果相似[14,30]。区系数量结构表明单种科、单种属占比较高,植物演替有极大的潜力,体现了科级和属级水平上的多样性[31]。
研究区植物群落以草本群落为主,茵陈蒿、葎草群落更为稳定,芦苇群落多样性最差。芦苇群落中芦苇集中大量分布,使得物种多样性较低,若生存环境发生变化,不适宜芦苇生存,则整个群落稳定性就会遭到极大破坏;而葎草与茵陈蒿群落物种丰富,生态幅度广泛,存在滩涂特色的植物[32]如碱蓬、盐地碱蓬等,能更好地适应环境。
土壤环境条件是影响植物生长的重要因素,土壤盐分和养分的空间分布差异在一定程度上决定着植物多样性的多寡[33]。土壤盐分是滨海湿地植物多样性的重要影响因素,通过对植物多样性指数和土壤环境因子进行相关性分析,本研究发现研究区内植物的R、H、D和E指数与土壤含盐量整体上呈负相关。其中样带1 深度为20~40 cm 土壤的盐分与R指数呈显著负相关,说明土壤盐分积累对植物多样性产生了不利影响。这可能是因土壤中较多盐分会抑制植物生长,甚至使一些耐盐能力较弱的甜土植物不能存活,进而降低了该区的物种丰富度[34]。该结果与张林静等[35]对新疆阜康地区、赵敏等[10]对黑河中游湿地以及Basharat等[36]对沙特阿拉伯沿海湿地植物多样性与盐分的研究结果一致。
植物生长过程也是对土壤环境适应和改造的过程[33]。植物多样性和土壤养分之间的关系前人已经做了大量研究,由于地区间土壤环境的差异,所得到的结果差异明显。刘俊娟[37]对我国丹江口湿地植物多样性进行研究表明,土壤全磷、全氮和有机质含量和植物多样性指数呈显著正相关。郭舜等[38]指出土壤养分对闽江河口湿地植被多样性贡献为正。而王琳等[39]指出随着有机质和氮含量增加,历山草甸区植被多样性指数在减少。湿地植物群落分布呈影响因素多样性和复杂性等特点。本文以土壤中有机质、速效氮、速效磷和速效钾等作为测量目标,分析土壤养分和植物多样性指数的相关性。结果表明,对于样带1 和样带3,R、H与D指数与速效氮、速效磷、速效钾和有机质存在显著相关性,氮素含量可以直接影响植物的生产力和植被类型[40]。本研究结果显示,研究区内土壤速效氮含量与植物多样性指数整体上呈正相关关系,尤其是与H指数在样带1 和样带3 内存在显著正相关关系,该研究结果与前人[40-41]对辉河和广佛地区典型湿地类型的植被研究结果相似,但与肖德荣等[42]对滇西北高原纳帕海湿地植物研究结果不同,可能是由于不同水域环境和气候条件对植物分布也有较大影响造成的。磷对作物生长有促进作用[43],关于速效磷对植物多样性的影响在同类湿地中得到强调[10,15],对于样带3,土壤速效磷含量越高,群落丰富度和复杂程度越高。有机质含量一定程度上代表了该地区土壤肥力的大小,是土壤养分的重要指标[44]。在盐度较高的样带1,R指数与土壤有机质存在显著正相关,可能是因为在土壤中增加有机质含量,有利于改善土壤透气性、渗透性和养分导致的[45-46]。
4 结论
1)研究区种子植物共有23 科47 属54 种,被子植物占绝对优势,植物中绝大部分是中生草本。区系地理成分特征相对复杂,数量结构特征表现出科级和属级水平上的多样性。
2)研究区植物群落植被以草本为主,可分为9种群落,其中茵陈蒿、葎草群落更为稳定,芦苇群落多样性最差。
3)土壤盐分和养分空间分布表明:随着土壤深度增加,土壤盐分呈增加趋势,速效氮和速效磷呈减少趋势,速效钾和有机质在不同样带区表现不一致。土壤盐分和速效钾高值区主要分布在研究区西南部地区,土壤速效磷高值区主要分布在西北部地区,而速效氮和有机质含量高值区空间分布较为分散。
4)研究区内,植物的丰富度指数、香农-维纳指数、辛普森指数以及均匀度指数与土壤含盐量整体上呈负相关关系,与土壤速效氮、速效磷和有机质多数样点呈正相关关系。样带1 丰富度指数与20~40 cm 土壤盐分含量呈显著负相关关系,对于样带1 和样带3,丰富度指数、香农-维纳指数和辛普森指数与速效氮、速效磷和有机质存在显著正相关性。该研究结果对于明确该区植物多样性影响因素具有重要意义,同时可为曹妃甸湿地植物多样性保护和管理提供一定科学依据。
