日粮添加精氨酸对家禽脂代谢和脂质沉积的影响
2023-07-14韩笑瑛董晓丽高玉田
韩笑瑛 董晓丽 高玉田
(希杰上海商贸有限公司,上海 200000)
为满足现代商品肉鸡生长需求,通常在生产中饲喂高能量和高营养水平日粮[1]。受品种选育特点和侧重于日增重和料重比的生长性能评价体系的影响,实现肉鸡体重快速增长的同时,富余的营养物质进一步转移至颈部皮下、腿部和腹部组织,造成以上区域脂肪沉积量增加[2]。研究表明,现代品系肉鸡体脂肪含量占机体总重的15%~20%,其中约85%的脂肪并非机体代谢必需[3]。沉积于肉禽腹部的非必需脂肪,因食用价值较低,往往在屠宰分割过程中被舍弃,直接造成能量饲料的浪费和胴体产量的降低,也影响了肉禽以活鸡或整鸡售卖时消费者的接受度,进而影响其经济价值[4]。解决腹部脂肪的过度积累是肉禽行业养殖加工一体化进程中亟待解决的问题[5]。
影响家禽脂代谢的主要因素包括机体脂肪合成能力和沉积两方面[6]。肉鸡脂质合成集中于肝脏,受CREB/CRTC2 转录复合物、胆固醇调节元件结合蛋白SREBP1等成脂关键蛋白以及mTOR信号通路调控[7]。家禽脂肪合成与日粮组成关联密切。最新研究表明,日粮中额外补充晶体精氨酸能够调节肉鸡肝脏脂肪合成,降低脂肪沉积[8]。精氨酸作为家禽的必需氨基酸,广泛参与机体蛋白质合成过程,具有免疫调节和抗氧化功能,其代谢通路也与谷氨酸、多胺、脯氨酸和谷氨酰胺等多种物质紧密相关[9]。本文从精氨酸对家禽脂代谢调节作用的角度入手,对精氨酸调控家禽机体脂肪沉积的相关机制和应用效果进行综述,为在生产中合理利用精氨酸调节家禽脂代谢、提高禽肉制品生产效率提供参考。
1 精氨酸代谢途径
1.1 精氨酸
精氨酸分子式为C6H14N4O2(见图1)。和主链最为接近的侧链部分是有机且疏水的,另一侧的侧链为一个胍基,因此该结构无论在酸性、中性或碱性环境中都带正电荷。双键和N 孤立电子对之间的共轭关系导致正电荷离开原位,最终胍基便能形成多重的氢键,在蛋白质外围产生稳定的相互作用,是机体内最丰富的蛋白氮载体[10]。研究表明,鱼类及鸟类的肝脏、肾脏、胰腺、肠道和脾脏中缺乏参与尿素循环和精氨酸代谢的氨甲酰磷酸合成等关键酶,不能通过尿素循环合成精氨酸,只能通过日粮获得[11-13]。精氨酸作为家禽的必需氨基酸,在以玉米-小麦-豆粕为基础的常用日粮结构下,往往占据第三或第四位限制性氨基酸的重要位置[11]。

图1 精氨酸分子式
1.2 代谢途径
精氨酸经采食进入体内,主要于小肠中段吸收。肠道上皮细胞是精氨酸分解代谢的主要场所,能够在结构性一氧化氮合成酶(cNOS)作用下生成具有生理调节功能性的一氧化氮(NO)。由cNOS 合成的NO能够作为内皮舒张因子,维持血管通透性、增加血流量、改善肠道缺血缺氧环境,进一步发挥细胞保护作用[14]。肠道上皮细胞能够分解生成鸟氨酸,最终以尿素的形式排出[15-16]。鸟氨酸是多胺类合成的前体物质,合成的腐胺能够继续生成亚精胺和精胺,促进细胞发育增殖[16]。腐胺、脯氨酸和谷氨酰胺统称为多胺,谷氨酰胺一方面作为碳源氧化供能,另一方面可通过细胞增容作用,促进肌细胞的生长分化[17]。研究表明,谷氨酰胺作为家禽机体D型蛋氨酸转化的氨基供体,与D型蛋氨酸转化成L型蛋氨酸的效率密切相关[18]。同时精氨酸也可经由胍丁胺和瓜氨酸合成多胺[19]。精氨酸在家禽体内通过鸟苷酸循环后分解为氨进而参与合成嘌呤,最终以尿酸形式排出体外,精氨酸代谢途径见图2。在图2中,ASL为精氨酸琥珀酸裂解酶,ADC为精氨酸脱羧酶,AGAT为精-甘脒基转移酶,ARG 为精氨酸酶,ASS 为精氨酸琥珀酸合成酶,DDAH 为二甲基精氨酸二甲基水解酶,OAT 为鸟氨酸转氨酶,ODC为鸟氨酸脱羧酶,OTC为鸟氨酸氨甲酰转移酶,P5C 为吡咯-5-羧酸还原酶,PRMT 为蛋白质-精氨酸甲基转移酶,NOHA为羟基-L-精氨酸。
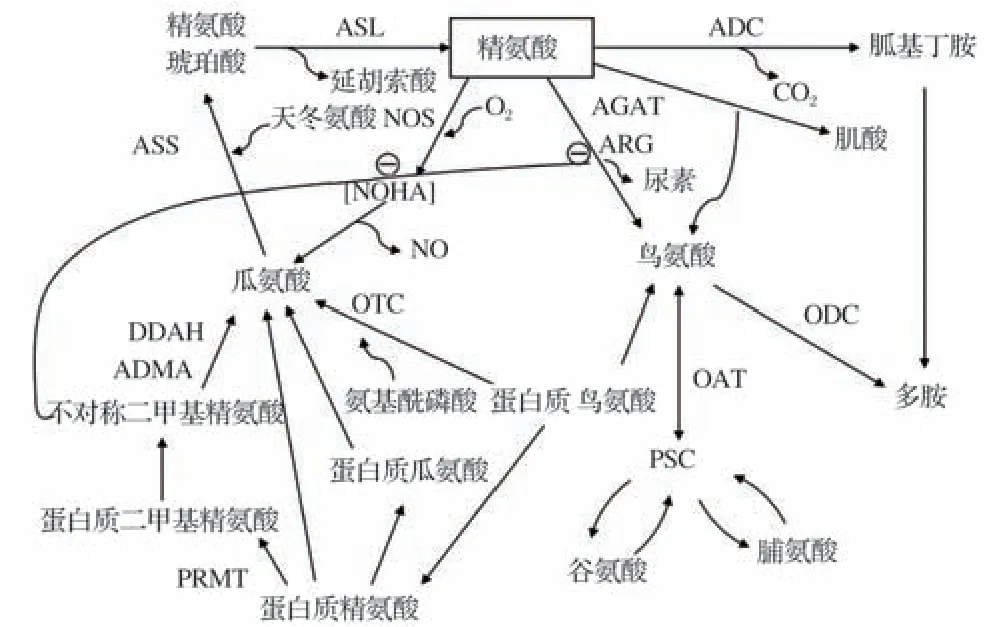
图2 精氨酸代谢途径
2 家禽脂代谢
日粮中的能量物质除用于维持家禽正常日常生长代谢外,其余部分在肝脏和脂肪组织中以脂质形式存留沉积。肝脏和脂肪组织是脂肪酸从头合成的主要位点,在家禽中,70%的脂肪酸于肝脏内合成[20-21]。脂肪酸提供能量并参与细胞膜合成,具有抗氧化损伤、抗炎症反应、调节肠道健康等多种生物学功能[22]。肝脏是脂质合成的主要场所,包括甘油三酯(TG)、磷脂和胆固醇等。血浆TG水平是影响家禽脂肪组织发育、沉积及蛋黄形成的主要因素。TG在肝细胞中合成,储存于脂肪细胞。相较于哺乳动物,生长阶段家禽的血浆富含TG载脂蛋白,即极低密度脂蛋白(VLDL)。其中VLDL主要由TG和载脂蛋白B组成[23],能够与肝细胞内源合成的TG结合,由肝脏经血浆转运至全身,并经过脂蛋白脂肪酶等作用先后异化产生中密度脂蛋白(IDL)和低密度脂蛋白(LDL),与在肝脏合成的高密度脂蛋白(HDL)一同将身体组织中的胆固醇转运至肝脏代谢,并在肝脏内再转化为胆汁酸排出[24-25]。研究表明,肉鸡70%的脂肪酸合成源于肝细胞,仅5%发生于脂肪组织。鸡肝细胞中脂肪生成及其在转录和转录后水平上的调节见图3。肝脏合成的VLDL、HDL运送至靶组织后(如脂肪组织),在脂蛋白脂肪酶(LPL)作用下水解以供立即利用或沉积[26-27]。

图3 鸡肝细胞中脂肪生成及其在转录和转录后水平上的调节
在图3 中,Dietary carbohydrate 为膳食碳水化合物,glycolysis pathway 为糖酵解途径,HKDC1 为己糖激酶结构域1,PK 为丙酮酸激酶,TCA cycle 为三羧酸循环,Pyruvate为丙酮酸,Pyruvate dehydrogenase为丙酮酸脱氢酶,Acetyl-CoA为乙酰辅酶A,Citrate synthase为柠檬酸合酶,Citrate 为柠檬酸,ACLY 为ATP-柠檬酸裂解酶,Oxaloacetate 为草酰乙酸,ACC 为乙酰辅酶A 羧化酶,Malate 为苹果酸,MD 为苹果酸脱氢酶,NADPH 为烟酰胺腺嘌呤二核苷酸磷酸,ME为苹果酸酶,SREBP1为固醇调节元件结合蛋白1,Insulin 为胰岛素,malonyl-CoA为丙二酰辅酶A,Estrogen 为雌激素,FASN 为脂肪酸合酶,C16∶0 palmitic acid 为棕榈酸,C18∶0 stearic acid为硬脂酸,ELOVL6为超长链脂肪酸延伸酶6,SCD为固醇辅酶A 去饱和酶,FADS1 为脂肪酸去饱和酶1,FADS2 为脂肪酸去饱和酶2,FA-CoA 为脂肪酸-酰基-辅酶A,ACSL 为长链酰基-辅酶A 合成酶,G3P 为3-磷酸甘油酯,Lysophosphatidic acid(LPA)为溶血磷脂酸,phosphatidic acid(PA)为磷脂酸,diacyglycerol 为二酰基甘油,triaylglycerol 为三酰基甘油,GPAT 为甘油磷酸酰基转移酶,AGPAT 为酰基甘油磷酸酰基转移酶,liver lipid secretion 为肝脏脂质分泌物,lipid droplets 为脂滴,LPIN 为磷酸酯磷酸水解酶,DGAT 为二酰基甘油酰基转移酶;MTTP为微粒体甘油三酯转运蛋白,VLDL为极低密度脂蛋白,SFA 为饱和脂肪酸,USFA 为不饱和脂肪酸,MUFA 为单不饱和脂肪酸,PUFA 为多不饱和脂肪酸,FFA为游离脂肪酸。
2.1 脂肪合成
肝脏作为家禽脂肪合成的主要器官,通过多种途径主导脂肪生成及葡萄糖-TG 转化[28]。肝细胞脂肪合成的上游过程为糖酵解途径,即通过己糖激酶结构域1 将葡萄糖磷酸化为葡萄糖6-磷酸的己糖激酶同种型。在糖酵解途径中,通过丙酮酸激酶在肝细胞的细胞质中合成丙酮酸。通过激活编码丙酮酸脱氢酶复合物(3种酶的复合物)的丙酮酸脱氢酶β PDHB和二氢硫辛酰胺乙酰转移酶基因,在线粒体内进行丙酮酸向乙酰辅酶A(CoA)的转化。在肝细胞线粒体中,草酰乙酸和乙酰CoA 通过介导柠檬酸合成酶转化为柠檬酸[29]。此外,草酰乙酸能够经由苹果酸脱氢酶基还原为苹果酸[30]。脂肪酸去饱和酶FADS2 作为脂质代谢途径中的关键限制性酶之一,能够将亚油酸和α-亚麻酸转化为多不饱和脂肪酸(PUFAs)。研究表明,家禽肌内脂肪富含PUFAs,FADS2 作为脂质代谢的调节剂,能够刺激胸肌和腿肌的肌内脂肪合成,改善嫩度、多汁性和风味等肉品质指标[31]。
2.2 脂肪沉积
肝脏中重新合成的脂肪酸能够与TG结合并储存。其中,TG以VLDL形态转运至脂肪和肌肉组织,在LPL催化下分解为脂肪酸和单酸甘油酯,能够穿过脂肪细胞膜并再次被合成[32]。研究表明,家禽80%~85%的脂肪沉积来自血脂,其脂肪沉积速率受血浆TG 浓度影响[33]。因此,脂质合成、脂质运输和脂质利用3 方面因素共同决定了家禽脂肪沉积的速率和总量[34]。鸡的沉积脂肪主要由腹部脂肪、皮下脂肪和肌内脂肪(IMF)组成。其中鸡的IMF含量依赖脂肪细胞的数量和脂质沉积能力[35]。
2.3 脂肪分解
家禽体内的脂质分解主要在线粒体中以β 氧化的形式完成。当脂肪酸被细胞吸收,细胞质中的脂肪酸会被运送入线粒体基质,进行β氧化供能[36]。活化的脂酰辅酶A 不能穿透线粒体内膜,需要经肉碱脂酰转移酶(CPT)催化。通过与线粒体膜外膜上的肉碱结合成脂酰肉碱,进入线粒体外膜[37]。在线粒体内膜上进一步转化为游离肉碱和脂酰辅酶A,进入线粒体基质后在脂肪酸氧化酶如长链脂肪酸氧化酶(LCHAD)的作用下进一步氧化[38]。整个过程从β位的碳原子开始,经脱氢、水合、脱氢和硫解,最终产生的乙酰辅酶A 进入柠檬酸循环并产生能量[39]。
3 精氨酸对家禽脂质合成和腹脂沉积的影响
3.1 精氨酸对家禽脂质合成的影响
活跃的脂质合成会加速家禽体脂沉积。已有研究表明,日粮中添加精氨酸可有效降低家禽体脂合成代谢活跃度[40-41]。Wu等[42]研究表明,与对照组相比,肉鸭日粮中添加1.0%的L-精氨酸显著降低了苹果酸脱氢酶(MDH)、葡萄糖6-磷酸脱氢酶和脂肪酸合酶的酶活,表明日粮添加精氨酸与肉鸭体内脂质合成紧密相关。相较于DL-精氨酸中的D型精氨酸需要在体内酶的作用下转化为L型精氨酸才能够被机体利用,L-精氨酸具有更高的生物效价。Fouad等[43]发现,与未添加精氨酸日粮组相比,饲喂添加0.25%和1.00%L-精氨酸的日粮显著下调了肉鸡肝脏中参与脂肪酸合成的脂肪酸合成酶(FAS)和3-羟基-3-甲基戊二酰辅酶A 还原酶的转录组表达,进而调控肉鸡脂肪合成。Tan等[44]研究表明,猪日粮中添加1.0%精氨酸显著增加了肌肉脂肪酸合成酶的mRNA表达水平,同时显著降低了脂肪组织中脂蛋白脂肪酶、葡萄糖转运蛋白4 和乙酰辅酶A 羧化酶-α 的mRNA 表达水平。这表明精氨酸对肌肉组织和脂肪组织中脂肪生成相关基因的表达调节存在差异,精氨酸能够促进肌肉组织中的脂质生成同时降低脂肪组织中的脂质合成。日粮添加精氨酸能够调节脂肪合成,减少体内脂肪沉积[45-46]。
3.2 精氨酸对家禽腹脂沉积的影响
家禽脂肪沉积集中于腹部、腿部和颈部皮下,其中腹部脂肪沉积速度最快[47]。研究表明,生长鸡中腹脂的增重速率高于体重增加速率,出栏前后腹脂沉积量约占总脂肪的20%[48]。腹脂率具有中等以上遗传力,受多基因调控,日粮组成是影响腹脂沉积的主要因素[49]。Filho等[5]研究表明,日粮添加6.87 g/kg精氨酸显著提高了29~42 日龄肉鸡饲料转化率,降低腹脂率和血浆甘油三酯水平。Fouad等[43]研究表明,日粮添加0.25%和1.00%L-精氨酸能够显著降低肉鸡腹脂率。Ebrahimi 等[48]研究表明,日粮中添加6.1~7.5 g/kgL-精氨酸显著提高了肉鸡10、24、46 日龄体重并降低腹脂率;显著提高了肌肉组织中肌间脂肪生成相关基因FAS和LPL的表达,降低了肝脏和腹部脂肪组织中该基因的表达。Corzo 等[50]研究表明,与基础日粮(精氨酸与赖氨酸比例0.90)相比,在大型罗斯肉鸡日粮添加精氨酸(精氨酸与赖氨酸比例1.45)减少了腹脂重量,显著提高了56 日龄胴体率。Khalaji等[51]研究表明,日粮添加8.4、12 g/kg精氨酸显著抑制了高能日粮对肉种鸡血浆TG、胆固醇和糖化红蛋白浓度的升高,避免了腹脂前体物质过度积累,并通过激活血浆过氧化物酶体增殖物激活受体-α、过氧化物增殖物活化受体-γ 通路,减少高能日粮引发氧化应激对肉鸡生长速率的负面影响。Jannatar 等[52]研究表明,日粮添加0.25%精氨酸能够降低饲喂高油高能日粮肉鸡的腹脂率和血浆胆固醇水平,增加回肠上皮IgG和IgM的蛋白表达水平,改善胴体性能的同时提高肉鸡肠道免疫功能。Nishchal等[53]研究表明,精氨酸能够显著降低蛋白不平衡日粮组肉鸡的42日龄腹脂率。与肉鸡相似,日粮添加10 g/kg精氨酸降低了北京肉鸭腹脂率和腹部脂肪细胞的大小(直径和体积),显著增加胸肌肌内脂肪含量和胸肌质量[54]。Fernandes等[55]研究表明,肉鸭21日龄胸肌重随日粮精氨酸水平(1.39%、1.49%、1.59%、1.69%、1.70%)增加呈线性增加,腹脂率则随日粮精氨酸水平增加呈下降趋势,提示日粮添加精氨酸能够有效降低肉鸭的脂肪沉积,促进肌肉增长。
4 展望
肉鸡和肉鸭是普遍认可的优质动物蛋白来源,其肉质和脂肪含量往往是评价其生产效率和经济价值的重要因素。高能量高蛋白日粮容易使家禽腹部脂肪过多沉积,降低经济效益。推广低蛋白日粮离不开多种必需氨基酸的添加补充。精氨酸作为一种必需氨基酸能够通过多种途径调节家禽体内脂肪代谢,有效减少脂肪堆积,提高饲料利用率。此外,精氨酸在降低家禽腹部脂肪堆积的同时,还能增加肉中蛋白质含量。精氨酸可作为优良的功能性氨基酸产品广泛应用于家禽饲料生产中。
