蚕豆亚有限生长型新种质的发现与鉴定
2023-07-10周仙莉滕长才郑栋刘玉皎
周仙莉 滕长才 郑栋 刘玉皎


摘要:蠶豆是重要的生物固氮和植物蛋白源作物,在改良土壤培肥地力、改善温带农业区持续蛋白生产方面尤为重要。然而,蚕豆品种的无限生长习性是导致机械化程度低的主因之一,极大地制约了蚕豆的规模化种植。为了探讨亚有限型在蚕豆育种中的应用及其对合理株型矮化育种研究的推动作用,通过查阅相关文献和多年的选择鉴定,对亚有限生长型新种质的发现过程和主要特征进行了总结归纳。对3个具有不同生长习性的种质与青蚕17号×RF25 F2分离群体主要农艺性状的表现差异进行了分析。结果表明,在F2群体中,亚有限型在开花层数、株高、结荚层数上与有限、无限型均存在显著差异,无限型与亚有限型的单株荚数、单株粒数、单株产量差异不显著,二者均显著高于有限型。说明亚有限生长习性蚕豆在品种改良后,能兼顾产量与机械化收获,在生产中应用优势更明显。
关键词:蚕豆;矮化育种;合理株型;亚有限生长习性
中图分类号:S529 文献标志码:A 文章编号:2097-2172(2023)06-0515-06
doi:10.3969/j.issn.2097-2172.2023.06.006
Discovery and Identification of New Germplasm in Vicia faba with
Semi-determinate Growth Habit
ZHOU Xianli 1, TENG Changcai 1, 2, ZHENG Dong 1, LIU Yujiao 2
(1. Academy of Agricultural and Forestry Science, Qinghai University, Xining Qinghai 810016, China; 2. Qinghai University
State Key Laboratory of Plateau Ecology and Agriculture, Xining Qinghai 810016, China)
Abstract: Vicia faba L. is an important biological nitrogen fixation and plant protein source crop. It is critical in improving soil fertility and sustainable protein production in temperate agricultural areas. The indeterminate growth habit of Vicia faba varieties is one of the major reasons for the low level of mechanization, which greatly restricts the large-scale cultivation of Vicia faba. In order to elucidate the application value of semi-determinate type in Vicia faba breeding and to promote research on dwarf breeding of ideotype in Vicia faba, this paper summarized the discovery process and main characteristics of the new germplasm with semi-determinate growth habit. Three Germplasms with different growth habits and a Qingcan 17×RF 25 F2 segregating population were used to analyze the main agronomic traits. Significant differences in the number of flowering layers, plant height, and number of podding layers between three growth habits were detected, while no significant differences in the number of pods per plant, seed number per plant, and yield per plant between the indeterminate growth habit and the semi-determinate growth habit in F2 population were found, and they were significantly higher than those of the determinate growth type. Therefore, semi-determinate growth habit of Vicia faba can benefit both yield and mechanized harvesting, its promotion and application values in production are obvious.
Key words: Vicia faba L.; Dwarf breeding; Ideotype; Semi-determinate growth habit
蚕豆是豆科巢菜属一年生或越年生作物,具有养人、养地、养畜的“三养”功能。蚕豆又是第四大冷季豆类作物,与其他豆类作物相比,可以适应不同的气候和土壤条件,在中国、埃塞俄比亚、澳大利亚、英国、摩洛哥等国家广泛种植[1 ]。据FAO统计,2020年世界蚕豆种植总面积为267.2万 hm2,总产量为566.9万 t。中国蚕豆种植面积和总产量均居世界首位,分别占世界的30.95%与30.41%。蚕豆营养丰富,因其蛋白含量高、氨基酸均衡及固氮能力强[2 - 3 ],在优化植物蛋白供应、丰富豆类蔬菜、保障区域饲草(料)、改良土壤结构和培肥地力方面发挥着重要作用,同时具有一定的药用和保健作用,市场前景较广[4 - 7 ]。适度增加蚕豆生产规模有利于统筹化肥减量与区域粮食安全,然而,蚕豆种植面积受劳动投入多、生产效率低而下降,导致供需差距较大[8 ]。
蚕豆生长习性与产量以及株型、荚分布、倒伏性、熟性等机械化收获指标密切相关。蚕豆的生长习性分为无限生长与有限生长[9 - 10 ]。目前,主栽蚕豆品种所具有的无限生长习性导致营养生长与生殖生长并进期过长,影响了产量的进一步提高,结荚分散,成熟不一致,限制了机械化收割,最终限制了生产效率的进一步提高,以及生产成本增加,直接制约着蚕豆的规模化种植[11 ]。有限型蚕豆具有的顶生花序、结荚集中、成熟期一致特性[12 ],是期望新品种所具备的优良特性,有助于促进蚕豆的规模化生产,节本增效。然而,有限型蚕豆由于开花与结荚层数的限制,增产潜力有限。亚有限型蚕豆品种顶生花序,能有效抑制顶端优势,成熟期相对一致,适应机械化收获。相对于有限型品种,其株高适中、株型紧凑、不易倒伏,同时开花和结荚层数较多,具有增产潜力。有限型、亚有限型相对于无限型较为早熟,将亚有限基因型导入抗旱品种中,也可扩大蚕豆在高寒农牧区的种植规模。以亚有限生长习性作为品种改良目标,统筹产量与机械化生产,塑造理想株型,培育既高产又能够适应机械化收获的优良品种,对于蚕豆生产节本增效、适度扩大生产规模,充分发挥其在轮作体系构建中的土壤改良作用和发挥其植物蛋白源作用具有重要借鉴意义。
1 蚕豆亚有限生长型新种质的发现
2004年,青海大学农林科学院豆类研究团队在国际干旱地区农业研究中心(ICARDA, The International Center for Agricultural Research in the Dry Areas)资源圃的试验中,筛选出一批蚕豆有限生长型种质。在对这些资源持续鉴定筛选中过程中,于2010年发现一单株与传统无限型、有限型均有显著差异,且国内无相关标准,或将其与有限型归为同一类[9 - 10, 13 ]。经过多年的鉴定筛选,发现亚有限生长习性能够稳定遗传,并形成了多个品系。该类种质植株顶生花序,大部分顶端花序畸形,无结荚能力;开花层数多为6~12层,分枝数多为1~3个,结荚主要集中在中下部。株高高于有限生长型,株型紧凑。参考大豆茎生长习性的分类[14 ],将此类种质的生长习性命名为亚有限生长。经过多年的选择鉴定,发现亚有限生长习性能够稳定遗传。2020年,将亚有限型种质RF25种植于云南,发现其主要特征未发生变化,说明亚有限生长习性在不同环境中表现稳定。
2 亚有限生长习性与无限、有限生长习性的比较
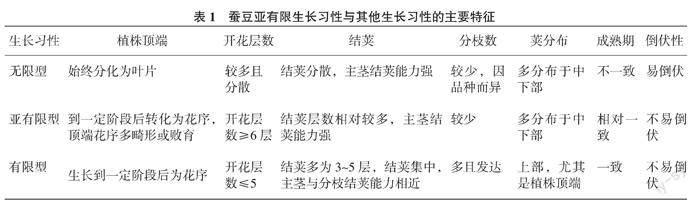
通过对不同生长习性的种质进行多年的观测,总结了3类蚕豆生长习性的主要特征(表1、图1)。无限生长型植株顶端始终分化为叶片,在肥水充足的条件下生育期延长,具有较大的增产潜力;有限型和亚有限型植株在发育到一定阶段后,其顶端均会转变为花序,有效地抑制了顶端优势,但亞有限型顶端花序多畸形败育。有限型与亚有限型生育期均较短,较为早熟,成熟期一致且结荚集中。
亚有限型在株型上与无限型相似,这也是其与有限型的主要区别。有限型与亚有限型的区别主要在开花和结荚层数、结荚部位、分枝数等方面,亚有限型开花层数较多(6层及以上),结荚多集中在中下部,分枝数少;有限型开花(≤5层)和结荚层数(3~5层)较少,集中在上部,尤其是植株顶端,分枝数较多。
3 不同生长习性种质的主要特征及农艺性状表现
3.1 主要特征观测
在网棚隔离田间栽培条件下,于2022年5 — 8月对青蚕17号(无限型)、RF22(亚有限型)、青蚕16号(有限型)主要农艺性状进行观测,结果见表2。开花层数、株高、结荚层数、单株荚数、单株粒数、单层荚数和单株产量等性状均以青蚕17号最大,RF22次之,青蚕16号最小。其中开花层数、株高、结荚层数、单株荚数、单株粒数品种间差异显著;单层荚数青蚕17号与RF22差异不显著,与青蚕16号差异显著;单株产量青蚕16号与RF22差异不显著,与青蚕17号差异显著。分枝数、有效分枝数、百粒重均以青蚕16号最大,青蚕17号次之,RF22最小。分枝数和百粒重青蚕17号与RF22差异不显著,均与青蚕16号差异显著。有效分枝数品种间差异不显著,可能是由于在网棚隔离条件下通风性较差,有限生长型部分分枝上的花序未发育成有效荚。
3.2 F2群体的主要农艺性状表现
2019年在网棚隔离条件下种植青蚕17号×RF25(亚有限型)F2群体,田间管理按常规进行,对群体内单株主要农艺性状进行观测,结果见表3。结果表明,在无限型品种与亚有限型种质杂交产生的F2群体中,出现了无限型、有限型、亚有限型3类生长习性的分离。
开花层数、株高、结荚层数、单株荚数、单株粒数、单株产量和百粒重等性状的均值均以无限型最大,亚有限型次之,有限型最小。无限型、亚有限型、有限型之间的开花层数、株高、结荚层数差异显著;无限型、亚有限型的单株荚数、单株粒数、单株产量差异不显著,与有限型的单株荚数、单株粒数、单株产量差异显著。百粒重无限型与亚有限型差异不显著,与有限型差异显著。分枝数有限型最大,亚有限型次之,无限型最小。有效分枝数亚有限型最大,有限型次之,无限型最小。分枝数和有效分枝数有限型与亚有限型差异不显著,与无限型差异显著。有限型与亚有限型分枝数分别为2.92、2.86个,较无限型略高;有限型分枝数变幅最大,为1.00~10.00个,有效分枝数为1.00~7.00个。
有限型的开花层数、株高、结荚层数、单层荚数、单株荚数、单株粒数、单株产量7个性状的标准差最小,开花层数的变幅最小。百粒重有限型的变异系数高于无限、亚有限型。在F2群体中,单株荚数、单株粒数、单株产量3个产量相关性状亚有限型显著高于有限型,说明亚有限型相对于有限型的增产潜力大,有利于选育相对高产的亚有限型优良品种。
4 亚有限生长型蚕豆种质的研究利用展望
生长习性是豆科作物重要的驯化特征,在形态特征、生态适应性育种研究中尤为重要[15 ]。Specht等[16 ]和Zhang等[17 ] 的研究表明,茎生长习性影响株高、花期、花和荚的空间分布、节点产量、成熟期、水分利用效率、环境适应性,并最终影响产量。亚有限生长习性类似于谷物“绿色革命”的半矮秆性状。亚有限型大豆的产量几乎与无限型品系相等,优于有限型品系[18 - 20 ]。鹰嘴豆的亚有限型抗倒伏,其产量潜力也与无限型相似,对蚕豆亚有限生长习性研究也得出了类似的结果。我们分析发现,亚有限型与无限型单株在单株荚数、单株粒数、单株产量、百粒重4个性状上不存在显著差异,说明以亚有限生长习性为选育与改良目标,在适应机械化收获的同时可以兼顾产量。亚有限型蚕豆品种在改良后,可抑制蚕豆无限生长,促进丰产和成熟一致、结荚相对集中,在生产中应用优势更明显。因而,对亚有限生长习性遗传及相关基因表达调控模式进行研究具有迫切性。
Bernard[14 ]和Liu等[21 ]发现,大豆亚有限生长习性受2对核基因(Dt1、Dt2)控制,2对基因间存在隐性上位作用。大豆Dt1基因与拟南芥TFL1基因同源,编码GmTFL1b蛋白,被定位于GLL连锁群(19 号染色体)上的Sat_099和Satt229标记间[22 - 25 ]。Dt2基因是AP1/SQUA亚家族的一个显性MADS域因子,通过Dt2蛋白Keratin(K)-box功能域与GmSOC1蛋白相互作用,从而抑制Dt1在茎尖分生组织中表达,促进茎尖分生组织向生殖生长转化,进而促进无限型大豆向亚有限型的转变[26 ]。Hegde[27 ]发现,鹰嘴豆的茎生长习性由2个具有显性上位性的基因控制,无限型基因型为(Dt1_Dt2_或Dt1_dt2dt2),亚有限型为(dt1dt1Dt2_),有限生长型基因型为(dt1dt1dt2dt2),为在早期区分鹰嘴豆无限型与亚有限型,鉴定了3个与Dt1基因座有关的SSR标记(TA42、Ca_GPSSR00560、H3DO5)。以上结果可为研究蚕豆亚有限生长习性相关基因提供借鉴。我们前期以蚕豆无限生长习性品种(青蚕17号)与亚有限生长习性品系(RF25、RF22)杂交构建了F2和F2∶3群体,遗传规律分析表明,蚕豆亚有限生长习性由2 对隐性核基因控制,控制生长习性的2对基因间存在显性上位效应,并初步定位了2 个与生长习性相关的基因,其遗传规律与大豆、鹰嘴豆不同,相关基因及其调控机制有待进一步研究[11 ]。
目前,利用亚有限型种质与其他品种/种质(羊眼豆、沙珠玉紫蚕豆、LF18、VF系列菜用品系、通蚕鲜7号、GF96等)进行杂交,将亚有限生长习性基因型导入不同的品种中,主要以亚有限+(早熟、抗病、圆粒、小粒、紫皮、菜用)等性状为选育与改良目标。在亚有限型品种选育、改良过程中,发现至少在F2世代及以上、植株开花末期才能对亚有限生长型进行选择,所需群体较大,选择效率较低。通过分子育种可以有效提高蚕豆育种效率,缩短育种年限[19, 28 ]。在发掘蚕豆亚有限生长习性相关基因后,需进一步利用基因全长序列及侧翼序列,开发与亚有限生长习性共分离或紧密连锁的分子标记,以期为利用分子标记辅助选育亚有限型优良蚕豆品种及创制优异种质提供依据。
参考文献:
[1] FAO. Statistical Database, Food and Agriculture Organization(FAO) of the United Nations[R]. Rome: URL, 2020.
[2] 趙克旺,张万伟,韩 宏,等. 临夏高寒阴湿区蚕豆不同种植方式比较试验[J]. 甘肃农业科技,2018(4):44-46.
[3] 余 莉,王昭礼,卢 云,等. 贵州省毕节市秋播蚕豆品种比较试验[J]. 甘肃农业科技,2015(1):36-38.
[4] 王姣姣. 冷季豆品质性状近红外模型建立及区域分析[D]. 北京: 中国农业科学院,2014.
[5] ROCHESTER I J, PEOPLES M B, HULUGALLE N R, et al. Using legumes to enhance nitrogen fertility and improve soil condition in cotton cropping systems[J]. Field Crops Research, 2001, 70(1): 27-41.
[6] JENSEN E S, PEOPLES M B, HAUGGAARD-NIELSEN H. Faba bean in cropping systems[J]. Field Crops Research, 2009, 115(3):203-216.
[7] RUBIALES D. Faba bean in sustainable agriculture[J]. Field Crops Research, 2009, 115(3): 201-202.
[8] 滕长才,刘玉皎. 青海省蚕豆未来产业发展探讨[J]. 青海农林科技,2022(2):50-52.
[9] 宗绪晓,包世平,关建平,等. 蚕豆种质资源描述规范和数据标准[M]. 北京:中国农业出版社,2006.
[10] 中华人民共和国农业部. 植物新品种特异性、一致性、稳定性测试指南:蚕豆NY/T 234 — 2013[S]. 北京:中国农业出版社,2013.
[11] 周仙莉,滕长才,张红岩,等. 蚕豆亚有限生长习性遗传规律分析及其基因初步定位[J]. 分子植物育种,2021,19(8):2660-2667.
[12] 郎莉娟,SAXENA M C,ROBERTSON L D. 有限花序蚕豆研究初报[J]. 作物品種资源,1990(3):13-15.
[13] 王金陵. 大豆遗传育种[M]. 北京:科学普及出版社,1957.
[14] BERNARD R L. Two genes affecting stem termination in soybean[J]. Crop Science, 1972, 12: 235-239.
[15] 高凤菊,张书良,赵同凯. 大豆结荚习性的研究进展及育种展望[J]. 大豆科技,2010(4):12-16.
[16] SPECHT J E, CHASE K, MACRANDER M, et al. Soybean response to water: A QTL analysis of drought tolerance[J]. Crop Sci, 2001, 41: 493-509.
[17] ZHANG D, WANG X, LI S, et al. A post-domestication mutation, Dt2, triggers systemic modification of divergent and convergent pathways modulating multiple agronomic traits in soybean[J]. Molecular Plant, 2019, 12: 1366-1382.
[18] HARTUNG R C, SPECHT J E, WILLIAMS J H. Modification of soybean plant architecture by genes for stem growth habit and maturity[J]. Crop Science, 1981, 21: 51-56.
[19] KATO S, SAYAMA T, TAGUCHI-SHIOBARA F, et al. Effect of change from a determinate to a semi-determinate growth habit on the yield and lodging resistance of soybeans in the northeast region of Japan[J]. Breeding Science, 2019, 69: 151-159.
[20] AMBIKA, HEGDE V, NIMMY M S, et al. Unraveling genetics of semi-determinacy and identification of markers for indeterminate stem growth habit in chickpea (Cicer arietinum L.)[J]. Scientific Reports, 2021, 11, e 21837.
[21] LIU Y, ZHANG D, PING J, et al. Innovation of a regulatory mechanism modulating semi-determinate stem growth through artificial selection in soybean[J]. PLoS Genet, 2016, 12(1): e1005818.
[22] LIU B H, WATANABE S, UCHIYAMA T, et al. The soybean stem growth habit gene Dt1 is an ortholog of Arabidopsis TERMINAL FLOWER1[J]. Plant Physiology, 2010, 153 (1): 198-210.
[23] LIU B H, FUJITA T, YAN Z H, et al. QTL mapping of domestication-related traits in soybean(Glycine max)[J]. Annals of Botany, 2007, 100(5): 1027-1038.
[24] TIAN Z X, WANG X B, LEE R, et al. Artificial selection for determinate growth habit in soybean[J]. Proceedings of the National Academy of Sciences, 2010, 107(19): 8563-8568.
[25] SCHUMUTZ J, CANNON S B, SCHULUETER J, et al. Genome sequence of the palaeopolyploid soybean[J]. Nature, 2010, 463(7294): 178-183.
[26] PING J, LIU Y, SUN L, et al. Dt2 is a gain-of-function MADS-domain factor gene that specifies semi-determinacy in soybean[J]. The Plant Cell, 2014, 26(7): 2831-2842.
[27] HEGDE V S. Morphology and genetics of a new found determinate genotype in chickpea[J]. Euphytica, 2011, 182(1): 35-42.
[28] DHAWAN G, KUMAR A, DWIVEDI P, et al. Introgression of qDTY1.1 governing reproductive stage drought tolerance into an elite basmati rice variety ‘Pusa Basmati 1 through marker assisted backcross breeding[J]. Agronomy, 2021, 11(2): 202.
收稿日期:2023 - 04 - 03
基金项目:国家现代农业产业技术体系专项(CARS-08)。
作者简介:周仙莉(1995 — ),女,山东济宁人,博士在读,研究方向为蚕豆种质资源研究与利用。Email: 18797001590@163.com。
通信作者:刘玉皎(1974 — ),男,青海互助人,研究员,研究方向为蚕豆遗传育种。Email: 13997058356@163.com。
