稀土矿区不同土地利用类型土壤碱性磷酸酶基因细菌多样性及其群落特征
2023-06-28杨贤房万智巍陈永林王远东
杨贤房,郑 林,陈 朝,万智巍,陈永林,王远东
(1.江西师范大学地理与环境学院,江西 南昌 330022;2.赣南师范大学地理与环境工程学院,江西 赣州 341000;3.广东省科学院生态环境与土壤研究所/ 广东省农业环境综合治理重点实验室,广东 广州 510650)
离子型稀土是国家重要的战略性资源,而长达半世纪的稀土开采造成了生态破坏、水土污染等问题。常用的原地浸矿工艺需沿注液井灌注大量浸矿剂〔(NH4)2SO4、NH4HCO3〕,残留在土壤中的 SO42-和 NH4+导致水体氨氮等指标超标[1]。矿区耕地因尾水灌溉导致作物生长困难、产量减少,矿区草地、林地、农田等土壤酸化。已有研究表明,矿区开采对周边农田土壤养分、重金属含量等造成影响[2],在酸性土壤中,磷缺乏是限制农业发展的主要因素之一[3]。探讨酸性矿山周边不同土地利用类型土壤酸化、有效磷含量及其解磷微生物响应问题,对于土壤磷调控及可持续利用具有现实意义。
土壤中磷主要以植物难以吸收的有机磷形态存在,有机磷必须转化为无机磷才能被大量吸收,而磷肥进入土壤后被钙、铁、铝等离子固定,形成无效态磷,其利用率通常不超过25%[4-5]。磷的高效利用与根系分泌物、微生物等有关[6],解磷细菌是土壤磷循环的主要驱动者[7]。细菌分泌的磷酸酶主要包括酸性磷酸酶[8-9]、碱性磷酸酶和植酸酶[10-11],是矿化土壤中有机磷的重要来源,其中phoD是细菌中最关键的碱性磷酸酶基因,在陆地生态系统中占主导地位[12]。研究耕地和草地土壤中phoD基因发现,含phoD基因的微生物可促进碱性磷酸酶分泌,进而使土壤中有机磷得到矿化[13]。由于根系分泌物的种类、数量及生长环境差异,不同植物解磷细菌的种类不同[14-15],休耕地、耕地和草地间phoD基因的细菌群落存在显著差异[16]。林地转耕地后phoD基因的细菌多样性和群落结构也会发生显著变化[17]。此外,土壤营养物质含量等是影响phoD基因群落结构的重要因素,较高pH值和较低的有效磷含量可以提高土壤中有机磷的矿化[18]。尽管当前国内外对phoD基因细菌多样性报道较多,但对酸性矿山尾水影响下的不同土地利用类型phoD基因细菌多样性及群落结构的研究较少。基于此,笔者采集尾水灌溉的弃耕地、农田以及泉水灌溉的蔬菜地、油茶地、脐橙地等土壤样品,采用高通量技术分析phoD基因细菌群落特征,目的在于探索赣南丘陵地区油茶地、脐橙地、耕地等用地类型土壤phoD基因细菌多样性及群落特征,揭示稀土矿区土壤phoD基因细菌多样性及群落结构对尾水灌溉的响应,以期为矿区土地利用可持续发展和生态重建提供科学指引。
1 材料与方法
1.1 样地设置及样品采集
样品采集区为江西省赣州市龙南足洞稀土矿区,赣州有着“稀土王国”“脐橙之乡”的美誉,同时是我国重要的油茶种植区。龙南足洞稀土矿区是我国最大的重稀土矿区,矿区周边分布有成片的油茶、脐橙、耕地等。采样区年平均气温18.9 ℃,年平均降水量1 526.3 mm,年平均蒸发量1 064.2 mm,取样时间为2021年7月中旬,天气晴朗,气温33 ℃。选择弃耕地(矿区内尾水灌溉)、农田(矿区内部分尾水灌溉)、蔬菜地(矿区内山泉水灌溉)、油茶地(矿区边缘山泉水灌溉)、脐橙地(矿区边缘山泉水灌溉)等样地(表1),设置20 m×30 m处理小区,对每个处理小区采用多点采样法,采集深度为0~20 cm的根际土壤,5个土芯混合为1份土壤样品,每份样品约100 g,每种土地利用类型共采3个样品,分成2份,一份样品用保温箱保存送往深圳微生物测序公司,另一份样品用于土壤理化性质测定。
1.2 研究方法
1.2.1土壤理化指标的测定
土壤有机质(SOC)含量采用重铬酸钾氧化-外加热法测定,有效磷(AP)含量采用钼锑抗比色法测定,土壤中铵态氮(NH3-N)和硝态氮(NO3-N)含量分别采用KCL浸提靛酚蓝比色法和双波长紫外分光光度法测定,土壤pH值采用pH计测定(水土比为1∶2.5),以上方法均参照文献[19]测定。
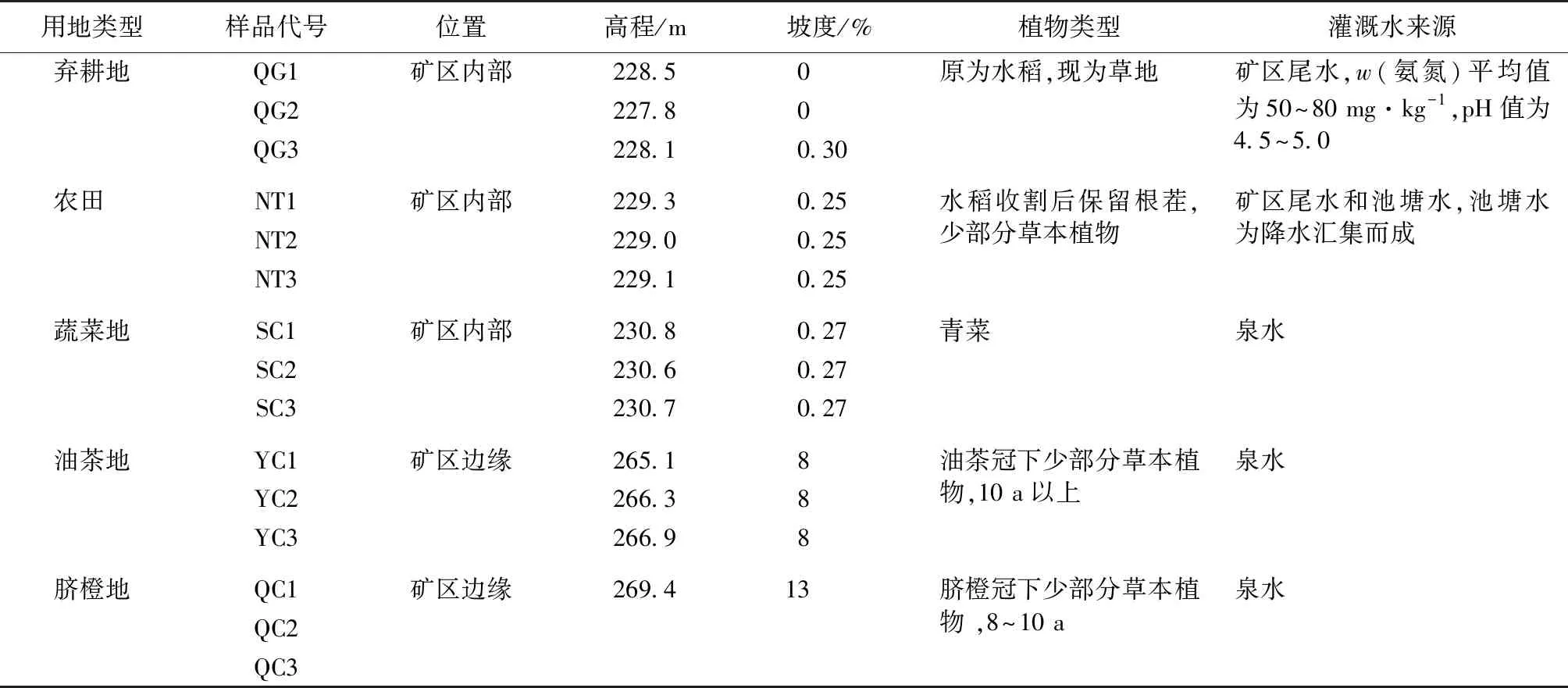
表1 样品采集信息
1.2.2土壤DNA提取与phoD基因细菌高通量测序
根据E.Z.N.A.® soil试剂盒(Omega Bio-tek,美国)说明书进行总DNA抽提,DNA浓度和纯度采用NanoDrop 2000进行检测,采用1%琼脂糖凝胶电泳检测DNA提取质量;phoD基因引物为phoD-733(5′-TGGGAYG ATCAYGARGT-3′)和phoD-1083 (5′-CTGSGCSAK SACRTTCCA-3′),片段长度约为350 bp[10],反应体系和条件为Phusion Master Mix(2×)15 μL,正反向引物(0.2 μmol·μL-1)1 μL,gDNA(1 ng·μL-1)10 μL,H2O 2 μL。98 ℃预变性1 min;30个循环(包括98 ℃变性10 s;50 ℃退火30 s;72 ℃延伸30 s),72 ℃延伸5 min。随后进行MiSeq文库构建及Illumina上机测序,测序由深圳微科盟科技集团有限公司完成。
1.2.3数据分析及方法描述
基于扩增子分析工具(QIIME)分析平台对测序原始数据进行拼接、过滤,以97%的一致性进行操作性分类单元(OTUs)聚类,并对有效序列进行物种注释。使用QIIME2 core-diversity插件计算多样性矩阵,α多样性指数包括OTUs数量、Chao1指数、Shannon指数等。使用多重柱形比较图寻找组间显著差异的物种,采用Duncan法对土壤理化性质和细菌多样性进行差异显著性检验。运用主成分分析法(principal component analysis,PCA)分析样本组之间的物种差异,通过冗余分析(redundancy analysis,RDA)和皮尔逊相关系数(Pearson correlation coefficient)分析物种与环境因子的关系。
2 结果与分析
2.1 土壤理化性质分析
由表2可知,不同土地利用类型的土壤pH值均呈一定的酸性,其pH值排序为油茶地>脐橙地>蔬菜地>农田>弃耕地。

表2 稀土矿区不同土地利用类型理化性质
其中弃耕地pH值仅为4.3,且同蔬菜、油茶地等用地存在显著差异(P<0.05)。蔬菜地铵态氮、硝态氮含量明显高于其他用地(P<0.05),其次为脐橙地,而弃耕地和农田的铵态氮和硝态氮含量最低。有机质含量为弃耕地土壤最低,其次为脐橙地,蔬菜地有机质含量显著高于其他用地(P<0.05)。全磷和有效磷含量为弃耕地最低,农田次之。总体上,弃耕地和农田的全磷、有效磷、铵态氮、硝态氮含量要低于蔬菜地、油茶地和脐橙地。
2.2 phoD基因细菌群落多样性与结构分析
不同土地利用类型土壤中phoD基因细菌多样性分析见表3。其中Chao1指数用来估计物种总数,值越大代表物种总数越多。Shannon指数用来反映物种的多样性程度,值越大表明群落物种的多样性越高[20]。结果显示,文库平均覆盖度均在0.98以上,表明测序深度能较全面地反映样品中的微生物信息。Chao1指数、OTUs数量、Shannon指数排序均为蔬菜地>油茶地>脐橙地>农田>弃耕地(表3)。OTUs数量和Shannon指数分别为116~286、5.4~6.9。其中农田和弃耕地OTUs数量仅为蔬菜地土壤的55.9%和41.3%,且弃耕地OTUs数量显著低于蔬菜地、油茶地和脐橙地(P<0.05)。蔬菜地和油茶地Shannon指数显著高于弃耕地(P<0.05)。

表3 稀土矿区不同土地利用类型phoD基因细菌α多样性
通过属水平主成分分析(PCA)发现,轴1的贡献率为22.00%,轴2的贡献率为19.55%,累计贡献率为41.55%(图1)。农田和弃耕地距离较近,表明两者具有相类似的phoD基因细菌群落结构,油茶地和蔬菜地距离近,显示不同土地利用方式的植物类型也是影响phoD基因细菌群落结构的重要因素。此外,蔬菜和脐橙地较近,表明2种用地类型phoD基因细菌群落结构存在一定程度的相似性。
不同土地利用类型phoD基因细菌在门、纲、属水平上的相对丰度如图2所示。在门水平上,变形菌门(Proteobacteria)均为不同土地利用类型的优势细菌(38.1%~79.5%),酸酐菌门(Acidobacteria)为脐橙地、农田、蔬菜地的主要优势细菌(26.5%~39.2%),浮霉菌门(Planctomycetes)为油茶地和蔬菜地的优势菌门(13.3%和15.3%)。在纲水平上,农田以γ-变形菌纲(Gammaproteobacteria,48.2%)为主要优势细菌,脐橙和蔬菜地以放线菌纲(Actinobacteria,34.1%和47.3%)、α-变形菌(Alphaproteobacteria,29.4%和23.5%)为主,弃耕地和油茶地以α-变形菌纲(82.7%和63.2%)为主。在属水平上,弃耕地主导性菌属为慢生根瘤菌(Bradyrhizobium,64.7%),农田优势菌属为假单胞菌属(Pseudomonas,47.7%)、链霉菌属(Streptomyces,24.5%)。脐橙优势菌属为链霉菌属(17.1%),油茶地主要由慢生根瘤菌(23.4%)、Stella(22.0%)、Brevundinonas(13.1%)等菌属组成,蔬菜地优势细菌属丰度较低,但种类较为丰富。

图1 稀土矿区不同土地利用类型含phoD基因细菌群落结构的主成分分析
组间差异分析结果显示,农田、弃耕地的多数菌属均显著低于蔬菜、脐橙、油茶用地(图3),不同土地利用类型组间假单胞菌属、链霉菌属、Gemmata、Kineococcus、Paludisphaera、Stella、未分类planctomycetes、Rhodoplanes等菌属相对丰度存在显著性差异(P<0.05)。其中尾水灌溉的农田、弃耕地土壤Gemmata、Kineococcus、Limnoglobus、Stella、Paludisphaera等菌属相对丰度均显著低于泉水灌溉土壤(P<0.05)。
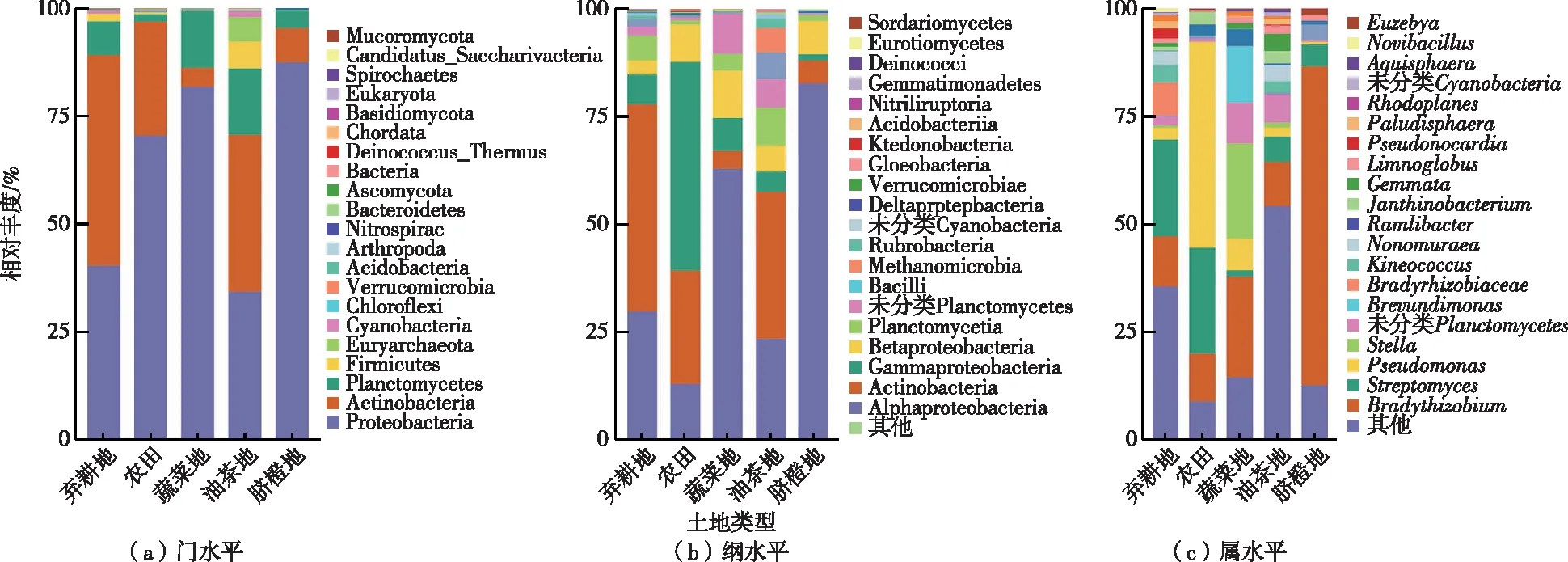
图2 phoD基因细菌在不同土地利用类型土壤中的相对丰度
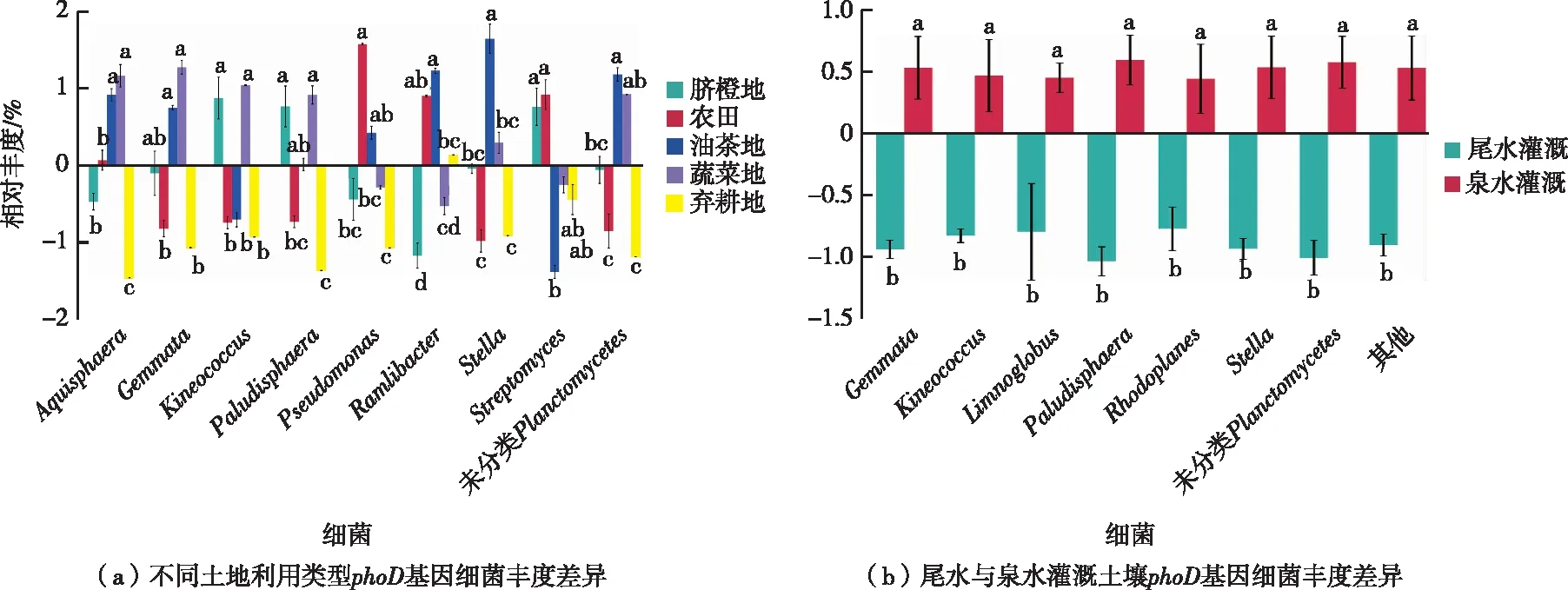
同一组直方柱上方英文小写字母不同表示不同土地利用类型间某指标差异显著(P<0.05) 。
2.3 phoD细菌群落的影响因子分析
将细菌群落α多样性与理化性质进行相关性分析(表4)发现,pH值、硝态氮含量、有效磷含量、有机质含量等均与phoD细菌Chao1指数呈显著相关性,但与Shannon指数未见显著相关性,其中pH值与Chao1指数相关性最高,表明pH值是影响phoD基因细菌丰富度的重要因素,硝态氮、有效磷、有机质含量与Chao1指数呈显著正相关,铵态氮含量未与Chao1指数呈显著相关性。RDA分析结果发现,pH值、硝态氮含量、铵态氮含量等能解释phoD基因细菌结构变化的49.11%。含phoD基因细菌优势类群与土壤理化性质相关性分析热图(图4)显示,硝态氮含量、pH值、有效磷含量是影响优势细菌的重要环境因子,其中硝态氮含量与Kineococcus、Paludisphaera、Gemmata相对丰度呈极显著正相关,与Limnoglobus呈显著正相关。pH值与Gemmata、Stella、未分类planctomycetes、Rhodoplanes相对丰度呈极显著相关,有效磷含量与Paludisphaera、Gemmata相对丰度呈极显著相关。
3 讨论
3.1 不同土地利用类型土壤理化性质差异及原因分析
矿区内尾水灌溉的弃耕地、农田pH值明显低于蔬菜地、油茶地和脐橙地。一般情况下,农业施肥措施在短期内较难观察到土壤pH值下降,而尾水浇灌的土壤pH值低于其他土壤,表明长期尾水浇灌降低了耕地土壤pH值,其中弃耕地土壤pH值仅为4.3,这与已有研究结果相同[21]。此外,尾水浇灌的农田和弃耕地的全磷、有效磷含量均低于蔬菜地、油茶地和脐橙地,这可能由土壤酸化、施肥管理等多种因素造成。有研究表明,由于酸性土壤中磷极易与Fe3+和Al3+结合,形成难溶态磷酸盐[22],土壤pH值与有效磷含量呈显著正相关[23]。此外,弃耕地和农田土壤铵态氮、硝态氮含量显著低于蔬菜等用地。一方面,此次采样为水稻收割后的农田和弃耕地土样,农田缺少后续施肥,而叶菜类蔬菜通常施氮肥较多。另一方面,尽管尾水灌溉可能增加土壤铵态氮等含量,但由于土壤反硝化易导致脱氮、养分流失等,尤其是农田闲置时期铵态氮、硝态氮含量变化较大,因此农田铵态氮含量并不比其他用地更高。此外,土壤中湿度、温度等也可能是影响铵态氮、硝态氮含量的重要因素。

表4 含phoD基因细菌α多样性与土壤理化性质的相关系数

图4 含phoD基因优势类群与土壤理化性质的RDA及Pearson相关性分析
3.2 不同土地利用类型phoD基因细菌多样性差异及原因分析
不同土地利用类型土壤中的phoD基因细菌多样性均存明显的差异,其中农田、弃耕地phoD基因细菌丰度仅为蔬菜地、油茶地和脐橙地的50%左右。一方面可能同土壤植被类型和覆盖率有关,通常情况下不同植被类型土壤多样性均存在差异性,此次采样时间为水稻收割后,而弃耕地多为草本植物,植物覆盖率相对其他土壤更低。另一方面,土壤理化性质是影响细菌多样性的重要因素。有研究发现,耕地长期施肥导致土壤酸化[17],进而影响phoD基因丰度下降。该研究中农田、弃耕地均采用稀土矿区尾水灌溉,土壤pH值较其他土壤更低,而phoD基因细菌的最适pH值在8~10范围内[24],这可能是弃耕地和农田土壤phoD基因细菌丰度较低的重要原因。此外,农田、弃耕地土壤硝态氮、有效磷等含量明显低于其他土地类型,硝态氮、有效磷含量与phoD基因细菌多样性呈显著正相关,pH值、速效磷和硝态氮含量对phoD基因细菌多样性的影响作用大于总磷和铵态氮含量[17]。
此外,土壤phoD基因细菌多样性与有效磷含量旦显著正相关,与当前学术界对phoD基因细菌多样性与有效磷含量关系的研究结果存在一定差异。有研究表明,有效磷含量与phoD基因细菌丰度呈负相关,施加磷肥导致phoD基因细菌丰度下降,而不施肥处理的根际土中phoD基因细菌丰度显著高于施肥处理[25];但也有研究显示,随着供磷水平的提高,含phoD基因的细菌群落α多样性还会提高[26-27]或无显著变化[28]。可见,有效磷含量与phoD基因细菌丰度的相关性同时受土壤pH值、土壤养分等多种因素影响。土壤pH值可能是影响有效磷含量与phoD基因细菌丰度关系的重要因素。一方面,尽管土壤低磷含量可能会促使微生物分泌更多的碱性磷酸酶,然而由于phoD基因细菌更适合在碱性环境中生存,phoD基因的细菌群落对pH值的响应更为强烈[29]。因此,低磷条件下phoD基因细菌丰度并未增加;另一方面,在酸性土壤中虽然酸性磷酸酶含量可能较高,但是相关研究表明碱性磷酸酶是最主要的有机磷水解酶[30],能水解90%以上的有机磷[25]。孙锋等[31]研究华南地区3种蟛蜞菊酸性土壤有机磷组分特征发现,有机磷矿化主要受碱性磷酸酶影响,而受酸性磷酸酶影响较小。郑曼曼等[32]研究酸性红壤解磷微生物丰度特征发现,phoD基因拷贝数同有效磷含量呈显著正相关,而phoC基因拷贝数未与有效磷含量呈现相关性。当然,关于酸性土壤中phoC和phoD基因细菌丰度同有效磷含量的相关性仍需开展更多的研究。就该研究而言,在酸性土壤环境中,土壤中的难溶性磷较难转化成易于吸收利用的有效磷,推测土壤pH值是影响稀土矿区磷源胁迫下有机磷矿化的重要因素,这可能是该研究中土壤有效磷含量与phoD基因细菌丰度呈正相关的重要原因,同时与该研究弃耕地、农田有效磷含量低,phoD基因细菌丰度也较低这一结果相吻合。
3.3 不同土地利用类型phoD基因细菌群落结构差异及原因分析
对解磷功能基因phoD进行高通量测序,全面分析了5种土地利用类型phoD基因细菌群落结构。在属水平上,慢生根瘤菌属是不同土地利用类型土壤优势phoD基因细菌,且弃耕地土壤中慢生根瘤菌属丰度高达64.7%,其他土壤慢生根瘤菌属相对丰度为8%~25%,LUO等[33]在不同施肥的农田中检测到高比例的慢生根瘤菌属,该菌属能通过增加碱性磷酸酶活性和磷转运速率响应磷胁迫[34]。此外,不同土地利用类型的假单胞菌属丰度呈现显著性差异,且假单胞菌属是农田、油茶的phoD基因细菌优势类群。陈定安等[35]从油茶树根际分离培养了假单胞菌属,该细菌有利于改善油茶磷素供应,促进油茶生长;陈倩等[36]从山西煤矿区分析筛选出假单胞菌属,发现其具有良好的解磷能力及较强的环境适应能力。链霉菌属为农田、脐橙地、蔬菜地、弃耕地优势属,该细菌在农田、脐橙地中占比显著高于其他用地,包含有既能分解无机磷,又能分解有机磷的菌种。优势属的明显差异可能受到植物凋落物以及根系分泌物的影响较大,同时土壤性质也是影响phoD基因细菌群落结构的重要原因。RDA分析和相关性热图均显示,pH值、有效磷和硝态氮含量等土壤理化性质与优势类细菌存在显著正相关性。
从不同土地利用类型phoD基因细菌群落显著性差异分析结果看,Gemmata、Stella、Rhodoplane、假单胞菌属、链霉菌属、Kineococcus、Paludisphaera等丰度在不同土地利用类型土壤中呈显著差异(P<0.05)。 其中,经矿区尾水灌溉的农田、弃耕地土壤Gemmata、Stella、Rhodoplanes、Kineococcus、Paludisphaera等细菌丰度显著性低于蔬菜地、油茶地、脐橙地等(P<0.05)。相关性分析结果表明,pH值与Gemmata、Stella、Rhodoplanes菌属丰度呈显著正相关,有效磷含量与Gemmata、Stella、Paludisphaera、Kineococcus丰度呈显著相关。由此可见,pH值和有效磷含量是影响不同土地利用类型phoD基因细菌差异的重要因子。尾水灌溉加剧了土壤的酸化程度,进而导致这些细菌丰度下降,同时也影响了土壤中有机磷矿化。丁翠等[37]研究了酸性矿山废水对稻田土壤微生物细菌群落的影响,发现土壤pH值是影响细菌群落结构的主要环境因子。也有研究发现,矿山废水污染程度可造成稻田土壤有效磷的显著淋失[38],可见酸性矿山pH值、有效磷含量、解磷功能细菌存在紧密的联系,仍需要长期且深入的调查研究。此外,综观现有文献报道,慢生根瘤菌属、假单胞菌属均包含有较强解磷功能细菌,在该研究中也分别是弃耕地、农田土壤中的绝对优势菌属,但2种细菌未同有效磷含量呈相关性,且弃耕地、农田有效磷含量较低。一方面,土壤理化性质与功能基因丰度的相关性比较复杂,几种因子相互作用抵消或受限样本数量,均可能影响有效磷与功能细菌丰度相关性。另一方面,慢生根瘤菌属仅在特定供磷水平下才与有机磷含量呈现出相关性[29],假单胞菌属通常在pH值为7时解磷能力最强[39],这表明解磷功能细菌对有机磷矿化的同时还受pH值、供磷水平等多种因素影响。在今后的研究中,应结合灌溉频率、灌溉时间、酸性磷酸酶活性和酸性磷酸酶基因细菌、土壤有机磷形态等数据进行综合分析,以期为酸性灌溉水源土地利用磷调控提供更多重要信息。
4 结论
稀土矿区不同土地利用类型土壤均呈现一定程度酸化现象,弃耕地和农田土壤pH值、有效磷含量低于蔬菜地、油茶地和脐橙地。phoD基因细菌丰度、多样性排序为蔬菜地>油茶地>脐橙地>农田>弃耕地。弃耕地与农田主导菌属分别为慢生根瘤菌和假单胞菌属,不同土地利用类型土壤假单胞菌属、链霉菌属等细菌相对丰度存在明显的差异性,尾水灌溉后Gemmata、Stella、Paludisphaera等细菌丰度显著低于泉水灌溉,pH值、有效磷含量与这些菌属丰度呈正相关。弃耕地与农田土壤phoD基因细菌群落较为类似,pH值、有效磷和硝态氮含量是影响不同土地利用类型phoD基因细菌群落结构的主要影响因子(P<0.05)。有效磷含量与phoD基因细菌丰度呈显著正相关,矿区尾水灌溉通过影响土壤中pH值、有机磷含量等理化性质,进而影响phoD基因细菌多样性与群落结构,研究结论可为离子型稀土矿区生态修复提供科学指引。
