长链非编码RNA C16orf89在胃癌组织中的表达及对胃癌细胞增殖和侵袭的影响
2023-06-21王长亮田文静黄景荣计春燕
王长亮,田文静,黄景荣,计春燕,王 慧,赵 莉,孙 俊
(1.湖北省中西医结合医院消化内科,湖北 武汉 430010;2.广州医科大学附属第一医院肿瘤科,广东 广州 510120)
胃癌是消化系统最常见的恶性肿瘤,病死率较高[1]。胃癌发病率在全球呈现增长趋势,同时其发病年龄逐渐年轻化[2]。尽管胃癌的诊断和治疗有了较大发展,但胃癌患者的5年生存率依然较低[3]。长链非编码RNA(Long non-coding RNA,lncRNA)在人体细胞中广泛表达,通过直接或者间接与染色体、微小RNA(microRNA,miR)、蛋白结合,调控相应基因的转录和表达,在维持染色体稳定方面发挥重要作用[4]。相关文献[5-6]证实,甲状腺癌、胸腺癌、淋巴瘤以及胃癌等组织中存在lncRNA的异常表达,其通过影响各种分子信号通路转导参与肿瘤的发生及进展。lncRNA C16orf89是一个由582个碱基组成的长链非编码RNA,其在胃癌组织及细胞系中表达模式和功能尚不清楚。因此,本研究探讨lncRNA C16orf89在胃癌中的表达模式,通过体外实验分析过表达lncRNA C16orf89对胃癌细胞增殖和侵袭的作用,通过生物信息学技术进一步分析lncRNA C16orf89可能的下游机制。
1 材料与方法
1.1 实验细胞 胃癌细胞系BGC823、AGS、NCI-N87细胞(购自美国ATCC公司)在含10%胎牛血清的RPMI-1640培养基中培养,胃癌细胞系SGC7901、HS-746T和永生化胃上皮细胞GES-1细胞(购自美国ATCC公司)在含10%胎牛血清的DMEM培养基中培养,在37 ℃、5% CO2条件下加湿培养。
1.2 主要试剂 胎牛血清(FBS)(批号:10100-147)购自美国Gibco公司;DMEM培养基和RPMI-1640培养基(批号:GT65-165、GT65-128)购自日本TOYOBO公司;lncRNA C16orf89过表达质粒pcDNA3-lncRNA C16orf89、阴性对照质粒pcDNA3-NC、miR-NC、miR-95-3p(批号:GNM25-113、GNM25-114、GNM25-115、GNM25-116)购自北京中杉金桥生物公司;噻唑蓝(MTT)试剂盒、实时定量聚合酶链反应(qRT-PCR)试剂盒、双荧光报告基因检测试剂盒(批号:E6819、E1520、E1910)购自美国Promega公司;Lipofectamine 2000、双荧光报告载体野生型pmirGLO-C16orf89-wt和突变型pmirGLO-C16orf89-mut(批号:11668019、17306001、17306002)购自美国Invitrogen公司;磷酸化磷脂酰肌醇3-激酶(p-PI3K)、磷酸化蛋白激酶B(p-AKT)、磷酸化雷帕霉素靶蛋白(p-mTOR)、细胞周期蛋白D1(CCDN1)、β-肌动蛋白(β-actin)、磷酸酯酶与张力蛋白同源物(PTEN)抗体(批号:ab278545、ab38449、ab109268、ab230947、ab8226、ab267787)购自美国Abcam公司。
1.3 研究方法
1.3.1 数据库分析:应用基因表达谱交互分析(GEPIA)数据库分析lncRNA C16orf89在胃癌组织及癌旁组织中的表达。应用Oncolnc数据库在线分析胃癌患者总生存期和lncRNA C16orf89表达的相关性。通过检索Starbase V3.0数据库筛选可能与lncRNA C16orf89结合的miRNA。
1.3.2 质粒转染:将生长状态良好且处于对数生长期的AGS细胞接种于细胞板,在培养箱内常规培养,保证细胞汇合度达到90%。分别将阴性对照质粒pcDNA3-NC和lncRNA C16orf89过表达质粒pcDNA3-lncRNA C16orf89转染至AGS细胞,根据Lipofectamine 2000试剂盒操作说明进行操作,定义为NC组和lncRNA C16orf89组。
1.3.3 实时定量聚合酶链反应(qRT-PCR)检测lncRNA C16orf89及miR-95-3p表达:采用TRIzol试剂盒提取各个细胞系总RNA,反转录为cDNA。以U6或GAPDH作为内参照,采用qRT-PCR检测lncRNA C16orf89及miR-95-3p表达。GAPDH正向引物序列为5’-TGCACCACCAACTGCTTAGC-3’,反向引物序列为5’-GGCATGCACTGTGGTCATGAG-3’。lncRNA C16orf89正向引物序列为5’-CCCTCCACTACCTCAAGCTG-3’, 反向引物序列为5’-TTTCCAGCAAATAGGGCAAG-3’。miR-95-3p正向引物序列为5’-ACACTCCAGCTGGGTTCAACGGGTATTTAT-3’, 反向引物序列为5’-TGGTGTCGTGGAGGAGTCG-3’。以2-ΔΔCt计算目的基因相对表达量。
1.3.4 MTT法检测AGS细胞增殖:以每200 μl即4×103个AGS细胞接种于96孔板。在AGS细胞贴壁后1、2、3、4、5 d时,分别在每孔中加36 μl(400 mg/L)MTT试剂,在37 ℃、5% CO2条件下加湿培养160 min。应用酶标仪检测各孔在波长490 nm处的吸光度值。
1.3.5 Transwell实验检测AGS细胞侵袭:培养箱内消化两组AGS细胞,加入无血清培养基中和消化酶并调整细胞密度,以4×104个/孔接种于Transwell小室上腔。在Transwell小室下腔中加500 μl含20%胎牛血清的DMEM培养基,在37 ℃、5% CO2条件下加湿培养25 h。采用5%多聚甲醛固定,在0.2%结晶紫染液中染色。干燥处理后,在倒置显微镜选择6个随机视野计数。
1.3.6 双荧光素酶报告基因分析lncRNA C16orf89靶基因:将双荧光报告载体野生型pmirGLO-C16orf89-wt或突变型pmirGLO-C16orf89-mut和miR-NC或miR-95-3p共转染AGS细胞,在37 ℃、5% CO2条件下加湿培养53 h。采用裂解液收集各组AGS细胞,通过双荧光素酶报告基因检测系统分析各组AGS细胞的相对荧光素酶活性,实验重复4次。
1.3.7 Western blot检测目的蛋白表达:采用细胞裂解液提取两组AGS细胞总蛋白,测定蛋白浓度后采用十二烷基硫酸钠聚丙烯酰胺凝胶电泳,电转至硝酸纤维素膜上,通过4%牛血清白蛋白溶液封闭3 h,裁剪硝酸纤维素膜,加入一抗p-PI3K(1∶1000稀释)、p-AKT(1∶2000稀释)、β-actin(1∶5000稀释)、p-mTOR(1∶2000稀释)、PTEN(1∶3000稀释)、CCDN1(1∶4000稀释),低温孵育13 h。加入二抗(1∶5000稀释),室温下反应3.5 h。加入化学发光液,通过凝胶成像仪拍照。

2 结 果
2.1 数据库分析结果 GEPIA数据库分析结果显示,与癌旁组织相比,lncRNA C16orf89在胃癌组织中表达下调(P<0.01)。Oncolnc数据库分析结果显示,与lncRNA C16orf89低表达胃癌患者相比,lncRNA C16orf89高表达的胃癌患者总生存期较长(P<0.01)。
2.2 胃癌细胞系及永生化胃上皮细胞中lncRNA C16orf89表达量比较 见图1。qRT-PCR结果显示,永生化胃上皮细胞GES-1和胃癌细胞系SGC7901、AGS、BGC823、HS-746T、NCI-N87中lncRNA C16orf89的表达量分别为0.99±0.13、0.39±0.08、0.17±0.05、0.58±0.12、0.71±0.04、0.64±0.09,胃癌细胞系中lncRNA C16orf89表达量低于GES-1细胞,且AGS细胞中lncRNA C16orf89表达量最低(均P<0.01)。
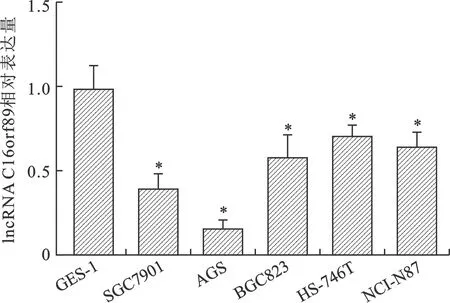
注:与GES-1细胞比较,*P<0.01
2.3 lncRNA C16orf89过表达质粒转染效率 qRT-PCR检测转染效率显示,NC组和lncRNA C16orf89组AGS细胞lncRNA C16orf89表达量分别为0.17±0.07和1.01±0.25,结果提示lncRNA C16orf89过表达质粒转染后AGS细胞中lncRNA C16orf89表达量升高(P<0.01)。
2.4 过表达lncRNA C16orf89对AGS细胞增殖能力的影响 见图2。MTT实验结果显示,与NC组比较,lncRNA C16orf89组AGS细胞从第2天起增殖能力明显降低(均P<0.05),因此过表达lncRNA C16orf89可明显降低AGS细胞的增殖能力。

注:与NC组比较,*P<0.05
2.5 过表达lncRNA C16orf89对AGS细胞侵袭能力的影响 见图3。Transwell侵袭实验显示,NC组和lncRNA C16orf89组AGS细胞侵袭数分别为(140.30±30.31)个和(49.48±19.07)个。与NC组比较,lncRNA C16orf89组AGS细胞侵袭能力明显下降(P<0.01)。

注:A图为两组细胞结晶紫染色结果(×100);B图中,与NC组比较,*P<0.01
2.6 生物信息学数据库Starbase V3.0预测结果 Starbase V3.0预测结果显示,miR-95-3p可能是lncRNA C16orf89的潜在靶基因,野生序列为“CCGUUGAA”,突变序列为“GGCAACUU”,见图4。

图4 lncRNA C16orf89与miR-95-3p的结合位点
2.7 双荧光素酶报告载体实验结果 见图5。在AGS细胞中,过表达miR-95-3p明显降低野生型lncRNA C16orf89-wt载体的相对荧光素酶活性(P<0.01),对突变型lncRNA C16orf89-mut载体的相对荧光素酶活性无明显影响(P>0.05)。

注:与miR-NC比较,*P<0.01
2.8 过表达lncRNA C16orf89对miR-95-3p的调控作用 qRT-PCR结果显示,NC组和lncRNA C16orf89组AGS细胞miR-95-3p相对表达量分别为6.54±0.89和1.02±0.48,因此过表达lncRNA C16orf89可明显降低AGS细胞中miR-95-3p的表达(P<0.01)。
2.9 过表达lncRNA C16orf89对PI3K/AKT信号通路蛋白表达的影响 见图6。Western blot结果显示,与NC组比较,lncRNA C16orf89组PI3K/AKT信号通路蛋白p-PI3K、p-AKT、p-mTOR、CCDN1表达量显著降低,PTEN表达量显著升高(均P<0.01)。
3 讨 论
胃癌的发生和发展涉及多基因、多分子信号通路的调控,伴随原癌基因的异常高表达和抑癌基因的异常低表达[7]。lncRNA在RNA的转录、加工和翻译过程中具有重要功能,影响细胞的应激、增殖、凋亡、自噬等过程[8-9]。研究[10-12]表明,特定lncRNA表达改变与胃癌的发生、演进具有相关性,同时与胃癌的淋巴结转移、大小、浸润程度密切相关。文献[13]报道,胃癌细胞系中lncRNA POT1-AS1高表达,其表达水平与胃癌患者临床病理学特征以及较短的无病生存期和总生存期有关,lncRNA POT1-AS1通过海绵化miR-497-5p促进胃癌细胞的增殖、迁移。文献[14]报道,lncRNA SNHG3在胃癌细胞系和组织中表达明显增加,其表达上调与胃癌临床分期、患者低生存率有关,敲低lncRNA SNHG3可在体内抑制异种移植胃癌的生长。lncRNA C16orf89基因定位于人16号染色体5060224-5065919区域,具有3个外显子,但其在胃癌细胞中发挥作用的分子机制尚不清楚。
本研究发现,lncRNA C16orf89在胃癌组织及胃癌细胞系中的表达均显著下调。与lncRNA C16orf89低表达的胃癌患者相比,lncRNA C16orf89高表达的胃癌患者总生存期较长。细胞功能学实验显示,过表达lncRNA C16orf89可在体外明显降低AGS细胞的增殖和侵袭能力。以上结果提示,lncRNA C16orf89在胃癌的发生、演进过程中可能发挥抑癌基因功能。文献[15]报道,lncRNA发挥功能的主要机制是通过海绵吸附miRNA,从而竞争性结合miRNA,引起miRNA的表达下调。例如,lncRNA LET通过海绵吸附miR-548k抑制胃癌细胞的增殖、侵袭和迁移,并促进细胞凋亡[16]。
本研究通过公共数据库预测发现,lncRNA C16orf89与miR-95-3p有潜在的结合位点。miR-95-3p在前列腺癌、肝癌、胶质瘤等肿瘤组织和细胞系中高表达,其分布在细胞质中,可明显促进肿瘤细胞增殖、侵袭,且在肿瘤进程中扮演原癌基因,与肿瘤患者的不良预后密切相关[17-19]。miR-95-3p在顺铂抗性的胃癌组织和细胞系中高度表达,其可有效促进胃癌细胞的增殖、侵袭、致瘤能力,在胃癌细胞中发挥促癌基因作用[20]。双荧光素酶报告基因检测证明lncRNA C16orf89与miR-95-3p间存在结合作用。同时,本研究表明lncRNA C16orf89过表达显著降低AGS细胞中miR-95-3p的表达水平。以上结果证实lncRNA C16orf89可以通过海绵吸附miR-95-3p发挥作用。PI3K/AKT信号通路能够促进胃癌细胞的增殖、转移并抑制其分化,该信号通路的持续激活显著促进胃癌的发生和演进。研究[20]证实,miR-95-3p能够在胃癌细胞中直接靶向激活PI3K/AKT信号通路。本研究证明,lncRNA C16orf89竞争性结合miR-95-3p后,PI3K/AKT信号通路蛋白表达量降低,亦证实了lncRNA C16orf89/miR-95-3p调控轴的存在。
综上所述,lncRNA C16orf89在胃癌组织和细胞系中表达下调,过表达lncRNA C16orf89能够抑制胃癌AGS细胞增殖及侵袭,其分子机制是通过靶向抑制miR-95-3p表达干扰PI3K/AKT信号通路的持续激活。lncRNA C16orf89异常表达可能参与影响胃癌患者的恶性进程。
