石山苣苔属的潜在适生区特征及其环境驱动变量
2023-06-18黄红温放李美君张金权周雨露李佺渊白新祥
黄红 温放 李美君 张金权 周雨露 李佺渊 白新祥


摘 要: 石山苣苔属(Petrocodon Hance)是著名的观赏花卉之一,但气候动荡和人类活动的强烈干扰,使其绝大部分被评估为极危(CR),至少也是易危(VU)以上。为重建末次间冰期以来石山苣苔属潜在适生区的时空变化,探讨适生区对环境变化的响应关系,为石山苣苔属的起源、地理分化研究和中国特有种质资源保护、园林开发利用提供理论指导,该研究结合120个分布记录和17个环境变量,应用优化的MaxEnt模型和地理信息技术(ArcGIS)对石山苣苔属在中国及中南半岛的适生区及其分布格局进行模拟,并基于逐步多元线性回归分析、冗余分析和蒙特卡洛检验评估影响石山苣苔属当前地理分布的主导变量。结果表明:(1)优化的MaxEnt模型的预测精度高,AUC值大于0.96;石山苣苔属当前适生区从中国西南部连续分布至越南北部,零散分布于中国中部和南部、块状分布于缅甸北部,其中以中国云贵高原南部为最佳适生区。(2)制约石山苣苔属当前地理分布的主导环境变量为最干月降雨量(bio14)、最热季度平均降雨量(bio18)、最湿季降雨量(bio16)、温度变化方差(bio4)、最冷月最低温度(bio6)、海拔(alt)。(3)在气候变化情景下,石山苣苔属适宜生境的扩张和收缩区位于当前潜在分布区的北部和东北部,是易受气候变化影响的敏感区。末次间冰期,石山苣苔属适生区大规模扩张,但末次盛冰期干冷的环境中几乎没有石山苣苔属适生区。之后,石山苣苔属的适宜生境向高纬度急速增加,而低纬度的适宜生境减少。(4)石山苣苔属适生区的质心从广西永福县(110.10° E、24.69° N)向北迁移至湖南城步县(110.29° E、26.05° N)。综上认为,全球气候变暖对石山苣苔属潜在分布区具有一定积极影响,但是极度的增温会造成石山苣苔属的适宜生境缩减、生态位变窄,而具有成熟喀斯特地貌优势的中国西南部至越南北部可能是其避難的主要场所。
关键词: 潜在适生区, 气候变化, 环境变量, MaxEnt 模型, 分布质心
中图分类号: Q984 文献标识码: A 文章编号: 1000-3142(2023)05-0799-18
Abstract: Petrocodon Hance is one of the famous ornamental flowers, but most of them are assessed as extremely dangerous (CR), or at least above vulnerable (VU) due to climate turbulence and strong interference of human activities. To reconstruct the temporal and spatial changes of the potential suitable areas of Petrocodon since the last interglacial period, and explore the response of the suitable areas to environmental changes, which provided theoretical guidance for the origin of Petrocodon, the study of geographical differentiation, the conservation of unique germplasm resources in China, and the development and utilization of gardens. In this paper, combined with 120 distribution records and 17 environmental variables, the optimal MaxEnt model and geographic information technology (ArcGIS) were used to simulate the suitable areas and distribution pattern of Petrocodon in China and Indo-China Peninsula. Based on stepwise multiple linear regression analysis, redundancy analysis and Monte-Carlo test, the dominant variables affecting the current geographical distribution of Petrocodon were evaluated. The results were as follows: (1) The prediction accuracy of the optimized MaxEnt model was high, and the AUC value was greater than 0.96. The potential suitable areas of Petrocodon for the present distribution are continuous from Southwest China to North Vietnam, scattered in central and southern China and blocky in North Myanmar, of which the South Yunnan-Guizhou Plateau in China was the most suitable. (2) The environmental variables which restrict the geographical distribution of Petrocodon included precipitation of the driest month (bio14), precipitation of the warmest quarter (bio18), precipitation of the wettest quarter (bio16), SD of the temperature seasonality (bio4), min temperature of the coldest month (bio6) and altitude (alt). (3) Under the climate change scenario, the expansion and contraction areas of the suitable habitat of Petrocodon were located in the north and northeast of the current potential distribution area, which were sensitive areas susceptible to climate change. During the last interglacial period, the suitable area of Petrocodon expanded on a large scale, but there was almost no suitable distribution area of Petrocodon in the dry and cold environment during the last glacial maximum. After that, with the aggravation of climate warming, the suitable habitats of Petrocodon increased rapidly to high latitudes, while the suitable habitat in low latitudes decreased. (4) The centroid position of the suitable area for Petrocodon migrated northwards from Yongfu County, Guangxi (110.10° E, 24.69° N) to Chengbu County, Hunan (110.29° E, 26.05° N). To sum up, global warming has a positive impact on the potential distribution area of Petrocodon, but extreme warming will cause the suitable habitat of Petrocodon to shrink, and the niche of Petrocodon will narrow. Southwest China to North Vietnam, which has the advantage of mature karst landform, may be its main refuge.
Key words: potential suitable area, climate change, environmental variable, MaxEnt model, distribution centroid
全球气候变化不断影响着物种的生活习性、地理分布、群落组成、生态系统结构(Doxford & Freckleton, 2012;赵泽芳等,2016; Matías et al., 2017),导致某些对生境专属性要求较强的植物迁移无路,加速物种灭绝;同时也改变物候节律引发生态紊乱(Sandel et al., 2011; Li & Chen, 2014)。气候变化引起地表温度和降水的重新分配,而地表温度和降水决定了植被的形成和演化(Lambert et al., 2010; Wang et al., 2014),进而显著影响扩散能力低、分布范围窄的稀有物种的丰富度格局(Walther et al., 2002; Hewitt, 2004; Svenning & Skov, 2007)。光照是植物生命活动的基础和能量来源,水分是植物生理活动的来源和基本成分(Fang, 1991)。因此,关注物种过去和未來对气候变化的响应,不仅有助于了解物种形成的历史原因和地理分布的变化,而且有助于制定科学的种质资源管理策略。
根据当前文献报道,石山苣苔属(Petrocodon Hance)主要分布于中国及中南半岛的喀斯特地貌区域(卢永彬等,2017),共计47种(含1变种),其中中国报道44种(GRC, 2022; 温放等,2022b)。该属植物花部变异几乎涵盖了所有苦苣苔科植物的花部特征(葛玉珍,2012;卢永彬等,2017),体态多样,花色艳丽,花冠局部还常兼有不同颜色的色晕、条纹、斑点或网纹,具有良好的产业开发应用前景(卢永彬等,2017)。该属植物分布于阴湿的喀斯特地貌(少量种类见于丹霞地貌)的峡谷、石缝、岩壁或溶洞口弱光带,对石灰岩钙化土的基质具有专一性,局限分布于一个大区域内生态环境最为优越的位置(温放,2021)。并且,该属植物多数物种数量少,分布范围窄,对环境的要求严格且环境适应脆弱,是评价当地生态环境优劣的重要植物类群之一(韦毅刚,2018;辛子兵等,2019)。一旦它们赖以生存的生态位发生变化,就意味着这个物种或这个地区某个群体的消失(葛玉珍,2012;温放,2021)。值得注意的是,由于气候动荡和人类活动的强烈干扰,近年发表的类群中除小黄花石山苣苔(P. luteoflorus)(Fan et al., 2020 )外,其余至少也是易危(VU)以上,多数被评估为极危(CR)(Li et al., 2019; Su et al., 2019; Zhang et al., 2019; Fan et al., 2020; Li et al., 2020; Xin et al., 2021; Nong et al., 2021)。
目前,对于石山苣苔属的研究主要集中在新分类群的发表、系统发育和濒危现状的再评估(葛玉珍,2012;温放等,2022a)。对石山苣苔属潜在适生区特征及其环境驱动因子还未进行系统和深入的研究,诸多价值还未挖掘出来,该属植物就已经处于极危的状态。虽已开展大量野外调研和濒危现状的再评估,但野外调查获得的数据和濒危等级不足以充分反映该属植物的整体分布格局。Liu等(2017)认为石山苣苔属物种分布与地理环境有着直接关系,受到诸多环境变量的制约。石山苣苔属是在特定地区或某种局部特殊生境下形成的物种,具有空间信息特征(温放等,2022b)。因此,重建不同气候条件下石山苣苔属在中国及中南半岛的分布格局,分析其生态适宜性意义重大。
物种能够在特定地区或某种局部特殊生境中生存和繁衍后代的所有条件总和称为生态位(乔慧捷等,2013)。生态位的研究可以通过生态模拟实现(朱耿平等,2013; Ahmed et al., 2015)。生态模拟是通过收集物种已知分布数据和生物环境变量,再结合统计学的运算方法推算研究物种的生态需求,并将运算结果投射到不同的时空中用以预测物种的潜在地理分布(朱耿平等,2013)。当前,应用最大熵模型(MaxEnt)模拟植物潜在地理分布已经成为一种研究趋势,并得到了较好的研究结果(Higgins et al., 2012; Cory et al., 2013; Higginset al., 2020; Zhao et al., 2021)。特别是将MaxEnt、ArcGIS、R语言及ENMTools多软件相结合与其他物种分布模型比较时,结果更接近于已实现的生态位(Higgins et al., 2012),并且预测的结果便于分析(Ahmed et al., 2015),准确度也相对较高(Phillips & Dudík, 2008; Elith et al., 2011),尤其在研究物种分布广泛和位置信息不确定中有很大的优势(Farashi et al., 2013)。因此,MaxEnt模型是目前使用最为广泛的生态位模型(Ahmed et al., 2015; Barbosa & Schneck, 2015; Vaz et al., 2015)。MaxEnt模型有助于推断物种已知分布范围内的潜在分布,但由于生物相互作用、扩散限制和在不适环境中的持久性(Higgins et al., 2020),可能会限制模型的外推能力,对物种已知分布范围外的分布预测存在局限性(Sillero, 2011)。因此,本研究选用MaxEnt模型来模拟不同气候条件下石山苣苔属在中国及中南半岛的潜在分布区。
当前,关于石山苣苔属适生区及其与气候变化关系的研究还很薄弱,该研究资料的缺乏不利于该属植物的价值评估和资源管理。本研究以中国和中南半岛为研究区域,选取17个环境变量,依托石山苣苔属地理分布信息,采用生态学模型和地理信息技术的方法,利用MaxEnt模型、ArcGIS、R语言及ENMTools多软件相结合,通过模拟预测石山苣苔属的潜在分布区和适宜生境,拟探讨以下问题:(1)预测石山苣苔属适生区的时空分布格局;(2)探索限制当前石山苣苔属地理分布的重要环境变量,综合分析该属植物地理分布与环境变量之间的关系;(3)提出保护和开发利用石山苣苔属植物的科学依据。
1 材料与方法
1.1 研究区概况
石山苣苔属集中分布中国及中南半岛,分布范围为98.8°—117.2° E、10.7°—33.3° N,分布海拔为120~1 700 m。从地形分布来看,研究区呈现北高南低的格局(钱满亿,2021),是亚洲热带的北缘,也是南亚季风、东亚季风和西北太平洋季风的叠加影响区,正是高山大河与喀斯特地貌集中分布的中国西南及中南半岛一带(姜超等,2017)。在当地复杂地形地貌及多种气候条件的影响下,形成了多种多样的局域性小气候和地带性植被(Zhu, 2013; Ren, 2015),是热带植物适应分化形成大量区域特有物种的一个热点地区(Myers et al., 2000)。
1.2 数据来源
1.2.1 物种分布的收集与整理 石山苣苔属的分布记录数据主要来源:(1)课题组野外调查;(2)植物标本和数据库,包括中国数字植物标本馆CVH(https://www.cvh.ac.cn/)、全球生物多样性信息网络GBIF(https://www.gbif.org/)、世界植物在线POWO(https://powo.science.kew.org/);(3)文献记载。为了避免因记录点之间的距离过近引起过度拟合。具体步骤如下:(1)将收集的分布数据按照物种名,经度(X),纬度(Y),并以*. csv文件类型进行保存,共计得到180个分布数据;(2)为了保证每一个物种有1分布点,先将分布记录小于3的物种直接用于模型预测,再将分布记录大于3的种进入下一步筛选;(3)以10 km范围内的多个样本点只保留一个点的原则,在ENMTools软件中利用除冗余数据(trim duplicate occurrences)工具删除10 km × 10 km栅格内重复记录数据(Bian & Shi, 2019);(4)經过以上筛选,最终获得120份石山苣苔属样本分布信息。
1.2.2 环境变量来源及筛选 本研究共计35个环境变量(表1)。其中,19个生物气候变量图层来自世界气象数据库(https://www.worldclim.org/),气候系统模型为CCSM4,该气候模型应用于气候模拟具有较大优势(肖建华等,2021)。气候数据共计6个时期,分别为末次间冰期 [last interglacial(LIG),距今120 000~140 000年]、末次冰盛期 [last glacial maximum(LGM),大约22 000年前]、全新世中期 [mid-holocene(MH),大约6 000年前]、当前时期(1950—2000年)、2050年(2041—2060年平均)和2070年(2061—2080年平均)。13个土壤变量数据来自粮农组织(联合国粮食及农业组织)(https://www.fao.org/land-water/databases-and-software/hwsd/en/)的世界土壤数据库(HWSD)。
地形变量中的海拔变量来自世界气象数据库(https://www.worldclim.org/data/worldclim21.html),坡度变量和坡向变量通过海拔变量在ArcGIS中利用表面分析工具获得。矢量地图数据来自自然资源部(http://bzdt.ch.mnr.gov.cn/)。本研究为了预测模型的顺利进行,首先需要将所有环境变量利用统一行政边界进行裁剪,并具有统一的栅格数据像元大小(0.008 3,0.008 3)、地理坐标系(WGS 1984)以及投影坐标系(WGS 1984 EASE Grid Global)。
在进行物种地理分布模拟时,为了避免各环境变量多重共线性而导致模型预测结果的过度拟合问题,首先需要预先运行MaxEnt模型,通过Jackknife(刀切法)检验环境变量的重要性及贡献率,剔除贡献率为0的变量(Zhu & Qiao, 2016)。再将分布数据和环境变量导入ArcGIS中,利用提取工具中的sample工具进行采样,把点带图层提取为Excel表格。然后在SPSS中分析石山苣苔属分布与环境变量之间的相关性,相关系数大于0.95的环境变量得以保留。最后利用ENMTools对环境变量进行皮尔森相关性分析,并在R中绘制皮尔森相关性热图(图 1 ),当相关系数大于0.8时,删减对预测贡献率较小的变量。经筛选得到9个气候变量(bio2、bio3、bio4、bio6、bio14、bio15、bio16、bio18、bio19)、5个土壤变量(t_texture、t_caco3、t_oc、t_sand、s_gravel)和3个地形地貌变量(alt、aspect、slope)。
1.3 模型参数优化
MaxEnt模型参数设置的复杂程度影响模型的转移能力(Merow et al., 2013; Qiao et al., 2015),而转移能力低的模型会造成预测结果不可靠或很难解释(朱耿平和乔慧捷,2016)。 研究表明,MaxEnt模型的复杂度与正则化乘数(regularization multiplier, RM)和特征组合(feature combination, FC)参数密切相关(Muscarella et al., 2015;朱耿平和乔慧捷,2016)。可以通过调用Kuenm数据包调控RM和FC,分析各模型参数的复杂程度,选取复杂度低而转移能力强的模型参数(朱耿平和乔慧捷,2016)。MaxEnt模型默认参数有5种特征,分别为线性(linear-L)、二次型(quadratic-Q)、片段化(hinge-H)、乘积型(product-P)和阈值性(threshold-T);RM的值为1。
在R中,调用Kuenm数据包优化MaxEnt模型(Cobos et al., 2019),将正则化乘数设置为0.1~4,每次间隔0.1,一共40种调控倍频;采用6个特征组合,即L、LQ、H、LQH、LQHP、LQHPT(朱耿平和乔慧捷,2016)。Kuenm数据包对上述31*40种参数组合进行测试。当满足pval_pROC显著,遗漏率(omission rates)<5%,W_AICc<2,delta_AICc=0,则特征组合模型为最优(Phillips et al., 2017; Cobos et al., 2019; Zhuo et al., 2020)。
1.4 模型模拟
将经过筛选得到的17个环境变量和120个分布记录分别导入MaxEnt 3.4.1中。模型参数设置如下:随机测试集为25%;绘制环境响应曲线和预测分布图;利用刀切法评估环境变量的重要性和贡献率;输出格式为Logistic;RM和FC根据模型优化结果设置。模型重复运行10次,最终输出结果文件为10次的平均值。除此之外,其他均保持默认参数。先利用ArcGIS 10.4对模型输出的栅格地图进行再次编辑,每个栅格像元的值表示存在概率,范圍从0到1,然后利用自然分割法(Jenks)对分布预测图进行重分类,将潜在分布区分为4个等级:不适生区(0~0.1)、低适生区(0.1~0.3)、高适生区(0.3~0.5)、最适生区(0.5~1),再利用手动法输入中断值。本研究将高适生区和最适生区定义为总适生区。
1.5 石山苣苔属自然分布对环境变量的响应关系
利用 SPSS 20.0 软件进行描述性统计来分析石山苣苔属适生区的气候特征,并采用变异系数(CV)来衡量各气候变量的变异程度(Fang & Yoda, 1991)。分别将17个环境变量与石山苣苔属的存在概率进行逐步多元线性回归分析,并在CANOCO 5.0 软件中采用气候变量与经纬度进行冗余分析(RDA),并采用蒙特卡洛(Monte-Carlo)检验量化各气候变量对石山苣苔属地理分布的贡献率,以此来判断影响石山苣苔属自然分布的主导气候变量。
1.6 适宜分布区空间格局
物种存在概率值≥0.30的空间单元定义为石山苣苔属适生区,物种存在概率值<0.30的空间单元被认为是不适生区。建立了当前、过去和未来气候情景下石山苣苔属潜在地理分布存在/不存在(0,1)矩阵。适宜生境被赋予表示存在的值(1),不适宜生境被赋予表示不存在的值(0)。以矩阵表0、1为基础,进一步分析了过去和未来气候情景下石山苣苔属适生区空间格局的变化。定义了4类适生区变化:新增适生区、缩减适生区、保留适生区和不适生区。过去和未来的面积变化是相对当前石山苣苔属适宜面积计算而得。
1.7 质心转移分析
首先,利用质心位置的变化来反映物种适宜生境的迁移方向和距离(Zhao et al., 2021)。然后,利用R软件中的Kuenm包计算了末次间冰期、末次冰盛期、全新世中期、当前和未来适宜区域的空间变化和适宜区域的几何质心位置(Bateman et al., 2016; Laurent et al., 2018)。最后,我们使用SDMs工具跟踪质心的经纬度,以检查不同时期石山苣苔属的质心位置,计算质心经纬度坐标并进一步得出迁移距离(Cong et al., 2020)。
2 结果与分析
2.1 模型优化及准确性评价
基于石山苣苔属的120个分布点和17个环境变量,利用MaxEnt模型模拟和预测该属植物的潜在适生区。根据模型优化结果(表 2 ),满足pval_pROC=0具有统计显著性、遗漏率=0.0333%小于5%,也小于MaxEnt模型默认值、W_AICc=1小于2,与其对应的RM=0.1和FC=L。
受试者工作特性曲线(receiver operating characteristic curve,ROC)面积为0.5~1.0。一般认为AUC值在0.5~0.7时,模型预测可靠性较低;在AUC 0.7~0.9时,模型预测可靠性中等;AUC大于0.9时,模型预测可靠性最高(Hosmer & Lemeshow, 2000)。我们的研究结果表现为各时期AUC值均>0.9(图 2 ),表明模型预测可信度高。
2.2 限制当前潜在分布区格局的环境变量
石山苣苔属存在概率与环境变量之间的逐步线性回归关系如表3所示:影响石山苣苔属存在概率的水分变量有最干月降雨量(bio14)、最冷季度平均降雨量(bio19)、最热季度平均降雨量(bio18)、最湿季降雨量(bio16);能量变量有昼夜温差月均值(bio2)、昼夜温差与年温差比值(bio3)、温度变化方差(bio4)、最热月份最高温(bio5);地形变量有海拔(alt)、坡向(aspect);土壤变量有顶层土壤碳酸钙(t_caco3)、底层土壤砾石含量(s_gravel)、顶层土壤砂含量(t_sand)、顶层土壤质地(t_texture)。以上主导变量解释率高达98.1%,且均达到极显著水平(P<0.01)。坡度(slope)、顶层有机碳(t_oc)、湿度变化方差(bio15)3个变量在拟合过程中被剔除,表明这3个变量对石山苣苔属的存在概率影响较小。
为了进一步解释石山苣苔属空间变化与环境变量的关系,将以上筛选得到14个环境变量和120分布记录构建矩阵,在CANOCO 5.0软件中进行冗余分析(RDA),经前向选择后,利用蒙特卡洛检验14个环境变量对地理分布的解释力排序(表 4 )。当P<0.005时,则该环境变量为影响石山苣苔属地理分布格局的主导环境变量。其中,贡献率达到极显著水平(P<0.005)的有6个环境变量,分别为最干月降雨量(bio14)、最热季度平均降雨量(bio18)、最湿季降雨量(bio16)、温度变化方差(bio4)、最冷月份最低温(bio6)、海拔(alt)。其中,能量变量bio4和bio6累计解释率和累计贡献率最高,分别为75.5%、79.4%;水分变量bio14、bio18、bio16累计解释率和累计贡献率为7.4%和8%;地形变量的解释率和贡献率分别为10.7%和11.7%;土壤变量的P值均大于0.05,未达到显著水平。因此,影响石山苣苔属地理分布的主导环境变量有bio4、alt、bio14、bio16、bio18、bio6。各类环境变量对石山苣苔属地理分布格局的影响力排序为能量变量>地形变量>水分变量。
对6个显著影响石山苣苔属地理分布的气候变量与地理分布数据进行RDA排序(图 3),结果表明前2个主分轴的解释率分别为73.32%和90.27%。其中,经度(X)与bio14、bio4呈正相关关系,与alt、bio6、bio18呈负相关关系,与bio16不相关;纬度(Y)与bio4、bio14呈正相关关系,与bio16、bio6呈负相关关系,与alt、bio18不相关。
为了解释6个主导环境变量与石山苣苔属地理分布的关系,通过绘制环境响应曲线来解释物种存在概率和环境变量之间的關系(图 4 )。其中,适生区的温度变化方差(bio4)的范围为98~920、最热季度平均降雨量(bio18)的范围是185~1 207 mm、海拔(alt)的范围是70~2 084 m、最干月降雨量(bio14)的范围是1~54 mm、最湿季降雨量(bio16)的范围是406~1 207 mm、最冷月最低温度(bio6)的范围是-6.17~3 ℃。6个主导变量的变异系数为17.20%~79.37%,最冷月份最低温(bio6)的变异系数最大,最湿季降雨量(bio16)的变异系数最小。综合各气候变量特点来看,石山苣苔属适宜分布区整体上具有湿热的气候特点(表5)。环境变量描述详见表 1。
2.3 石山苣苔属当前潜在地理分布特征
对120个地理信息在ArcGIS中进行定位,结果显示,石山苣苔属的实际分布点主要连续分布在中国西南部至中南半岛北部,零散分布于华南沿海和华中地区。预测分布(图5,图6)表明,120个石山苣苔属分布记录点中有103个点被包含在潜在的适宜区内,表明预测结果与实际分布基本保持一致。在当前气候情景下,总适生区面积为1.87×107 km2,占中国和中南半岛陆地面积的11.66%;高适生区面积为9.98×106 km2,占研究区总陆地面积的6.21%;最适生区面积为8.76×106 km2,占总面积的5.45%。结果表明,当前石山苣苔属总适生区范围为104°—112° E、 18°—32° N,A. 末次间冰期(LIG,距今120 000~140 000年); B. 末次冰盛期(LGM,大约22 000年前); C. 全新世中期(MH,大约6 000年前); D. 当前(1950—2000年); E. 2050年(2041—2060年平均); F. 2070年(2061—2080年平均)。
从中国西南部连续分布至越南北部,零散分布华南沿海和华中地区,块状分布于缅甸北部。
2.4 气候变化下石山苣苔属分布格局和面积的时空变化
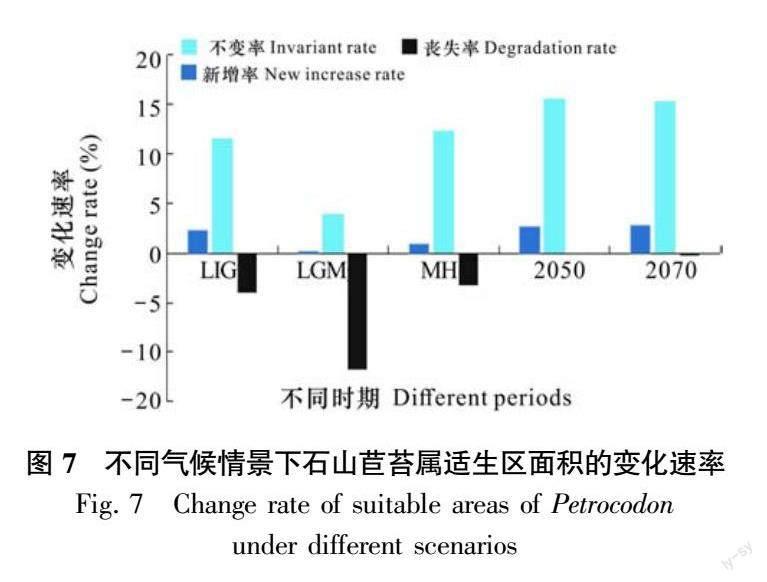
根据当前的地理分布记录和环境数据,绘制了石山苣苔属潜在分布区预测图(图5,图6)。末次间冰期适生区面积占总面积的10.73%(表6),在当前时期总适生区面积的基础上新增率为2.26%,丧失率为3.96%,新增率小于丧失率。当前时期适生区面积占比为11.66%,其占比高于末次间冰期。但是,当前适宜生境的分布中心相比末次间冰期向高纬度山区扩散,并且适宜区破碎化增强。比较末次冰盛期与当前潜在分布区的格局表明,末次冰盛期几乎没有该属植物适宜的分布区,丧失率高达11.71%,远大于新增率,表明气候寒冷干燥不利于石山苣苔属的生长。全新世中期与当前相比,该属植物的适宜生境轮廓与当前相似,适生区面积占比为10.33%,新增率0.86%,丧失率3.26%,新增率小于丧失率,表明该时期气候条件仍然不利于石山苣苔属的生长。在历史气候条件下,自末次间冰期以来,中国云贵高原至越南北部高度适生区均明显缩减与破碎化。
在未来气候情景下(图 7 ),于2050年石山苣苔属的适宜生境占比(14.66%)达到峰值,新增率2.61%,丧失率0.04%,新增区域位于当前时期适生区中心的东部和北部。2070年适宜生境占比为14.17%,新增率2.72%,丧失率0.24%。未来两个时期新增区域均位于现代时期适宜生境的北部和东部,这是物种对未来气候变化反应的敏感区域,这些区域应为重要保护区域。同时,表明在2050气候变暖的环境条件下,高适生区呈现集中分布,并且石山苣苔属植物的分布在一定的温度升高范围内处于积极状态,直到2070将会随着温度的升高适生区面积有所降低。
2.5 气候变化背景下石山苣苔属潜在分布中心的迁移路线
末次间冰期适宜区质心(图 8)位于广西永福县(110.10° E、24.69° N),末次间冰期向西北迁移到贵州丹寨县(107.94° E、26.16° N),末次冰盛期向东南部迁移至广西三江县南部(109.60° E、25.54° N),全新世中期再向东北方向迁移至湖南省通道县东部(109.95°E、26.24°N),之后向东部迁移至湖南城步县西北部(110.29° E、26.05°N),2070年质心点向东南迁移湖南城步县南部(110.19° E、26.30° N)。末次间冰期相比当前向南迁移179 km;末次冰盛期相比当前向西迁移194 km;全新世中期相比当前向西南移动87 km。此外,2050 年向东北迁移25 km,2070年东南方向迁移40 km。自末次间冰期以来,石山苣苔属适宜生境总体向高纬度地区迁移,迁移至湖南省南部为最北界,并且各时期迁移距离存在较大差异。
3 讨论与结论
3.1 模型精度分析
MaxEnt模型的参数默认设置是通过对来自6个地区的226种物种的不同数据集进行测试而生成的 (Phillips & Dudík, 2008), 应用ROC曲线评A. 温度变化方差(bio4); B. 海拔(alt); C. 最冷月最低温度(bio6); D. 最干月降雨量(bio14); E. 最湿季降雨量(bio16); F. 最热季度平均降雨量(bio18)。
价模型精度。但是对于特定物种的研究,使用专门的模型参数设置比使用默认设置更有效(Anderson & Gonzalez, 2011),可以有效地避免过度拟合(Zhu & Qiao, 2016)。优化的MaxEnt模型预测结果得到平滑的响应曲线且AUC≥0.96,表明微调模型可以合理地反映该属植物地理分布对气候变量变化的响应(朱耿平和乔慧捷,2016),并可以准确预测其潜在分布。这表明此模型用于石山苣苔属潜在分布区的预测可信度高。
3.2 石山苣苔属潜在适生区现状分析
MaxEnt模型预测结果表示石山苣苔属分布的最大可能性,不能准确得出物种现实的分布范围,而是物种的潜在分布区。本研究结果表明:从末次间冰期(LIG)开始至2070年经过6个时期,石山苣苔属潜在区从中国西南部连续至越南北部,零散分布中国南部和中部及块状分布缅甸北部。A. 末次间冰期(LIG,距今120 000~140 000年); B. 末次冰盛期(LGM,大约22 000年前); C. 全新世中期(MH,大约6 000年前); D. 当前(1950—2000年); E. 2050年(2041—2060年平均); F. 2070年(2061—2080年平均)。
因此,中国西南部至越南北部的地形地貌优势极有可能是石山苣苔属避难的主要场所。这些地区为石山苣苔属适生区,也是石山苣苔属种质资源热点地区,是未来石山苣苔属重点保护的区域。
如前文所述,季风是石山苣苔属从典型热带地区扩散至热带北缘的中国西南地区并存活下来的先决条件(Ren, 2015),且高山河谷与喀斯特地貌是该属植物的主要分布区域。研究区是亚洲热带的北缘以及南亚季风、东亚季风和西北太平洋季风的叠加影响区,也是高山河谷与喀斯特地貌集中分布区(姜超等,2017),该区域局域高度异质性的地形地貌造成多样的生境有利于石山苣苔属的生长与繁殖,使得扩散到这里的石山苣苔属个体逐渐适应季节性干湿气候、石灰岩高钙土壤等特殊环境。从季节角度分析发现,南亚季风在5—7月强度最大(周浙昆等,2017),东亚季风6—8月强度最大(吴国雄等,2013),西北太平洋季风5—9月强度最大(黎磊等,2014),在季风作用最强的阶段正好为该属植物生长发育季节。因此,在石山苣苔属的引种驯化实践中,在选择引种植基地时着重考虑当地季风情况。
3.3 环境变量与石山苣苔属潜在地理分布变化的关系
由于石山苣苔属本身的生理特性和所在生境的特殊性,使其仅分布于特定地区或某种局部特殊生境。在不同时期内,影响石山苣苔属分布的土壤变量和地形地貌基本相同,不会有较大波动,而致使石山苣苔属分布区变化的可能是由于气候变量的剧烈波动以及人类活动的强烈干扰。当前,制约石山苣苔属地理分布的主导环境变量为最干月降雨量(bio14)、最热季度平均降雨量(bio18)、最湿季降雨量(bio16)、温度变化方差(bio4)、最冷月最低温度(bio6)、海拔(alt)。可见,温度和降水作为重要的环境变量显著影响着石山苣苔属的生理特征、分布范围、多样性和丰富度等,海拔变量也具有显著影响作用是通过重新分配降水和温度来实现(Lambert et al., 2010; Wang et al., 2014)。
石山苣苔属5—9月开花,10—12月形成成熟蒴果,整个营养生长和开花结果阶段正好与研究区最暖季度重合。因此,干旱和寒冷季度可能会阻止该属植物的繁衍。同时,温度变化方差(bio4)和最冷月最低温(bio6)为主导环境变量合理性在于一定温差和最低温是植物的生长、发育和开花最重要调控因子之一(Khodorova et al., 2014),但是,低于-6.17 ℃极端低温不利于石山苣苔属的生长,从而影响其在地理尺度上的分布格局。因为石山苣苔属从热带或湿热地区进化而来,可能不具备抵御寒冷的机制(Li, 1996)。因此,在石山苣苔属的引种栽培中,在选取作为选育优质种源的种植基地时要注意当地温度变化方差和最冷月最低温,应优先选择最冷月最低温为-6.17~3 ℃、温度变化方差为98~920的区域。
此外,Liu等(2017)研究表明降水和海拔都不会显著影响苦苣苔科植物地理分布模式,但是本研究表明,最干月降雨量(bio14)、最热季度平均降雨量(bio18)、最湿季降雨量(bio16)和海拔(alt)对石山苣苔属的地理分布有着显著的影响,可能是由于石山苣苔属主要生长在喀斯特岩壁上,最热季度平均降雨量(185~1 207 mm)可加速石灰岩的矿化速率,可为其生长发育提供了更多的养分(徐翔等,2018)。石山苣苔属的存在概率随海拔的上升而减少的趋势,且峰值出现在中低海拔段(0~1 500 m),与实际分布海拔(120~1 700 m)相比预测分布区范围更广,但是超过1 500 m的高海拔区域不利于大多数石山苣苔属的生长。正如水分能量假说所认为,水分和热量的共同作用影响着植物物种分布的格局(Allen, 2002; Hawkins et al., 2003; Bradford et al., 2003;王志恒等,2004a)。所以,我们推测,高海拔区域存在概率低主要是由于低温环境降低了石灰岩的矿化速率;喀斯特地貌特殊的双层地质结构,致使水分迅速流失,从而形成了干旱低温缺水的喀斯特地貌环境不利于石山苣苔属的生存(Hawkins et al., 2003;王志恒等,2004b)。因此,在石山苣苔属未来的生产实践中,要注意当地降水量,尤其要重点关注最热季度平均降雨量的适宜取值为185~1 207 mm、最干月降雨量适宜取值为1~54 mm、最湿季降雨量适宜取值为406~1 207 mm的地域。
综上所述,各项环境变量通过控制石山苣苔属植物的生长和发育,从而进一步影响其分布格局。首先,能量变量、海拔变量、水分变量在塑造石山苣苔属生态适應方面发挥着主要作用,而土壤变量对其影响较小,可能是因为石山苣苔属多在喀斯特地区岩石表面半附生至附生生活,在大尺度区域条件下土壤的理化性质不足以代表石山苣苔属生长发育的土壤机制。其次,在进行地理分布数据统计以及查阅相关研究发现,石山苣苔属植物地理分布信息均来自喀斯特地貌(除星萼石山苣苔),因此,石山苣苔属(除星萼石山苣苔,该种目前仅已知其分布在广西资源县和湖南新宁县两县交界的丹霞地貌上)所有种均在相同的喀斯特地貌中。在此条件下,能够反映喀斯特地貌背景的土壤变量的差异不能从该属植物种间的地理分布中表现出来。
3.4 石山苣苔屬适宜区空间分布格局对气候变化的响应
根据模型结果,从末次间冰期开始到未来气候(2070年)6种气候情境中,石山苣苔属在中国及中南半岛的适生区面积在末次冰盛期断崖式降低,全新世中期成倍增加,之后缓慢增加,直至2050年停止增加,于2070年出现缓慢降低趋势,且整体有向高纬度地区移动的趋势。根据气候稳定性假说,气候波动会导致物种濒危,而稳定的气候往往会增加一个地区的物种丰富度(Stevens, 1989)。在末次间冰期间,东亚季风增强、降雨增加,全球温暖程度与当今的增暖相当,适生区分布接近当代分布格局。但是适生区面积相比当前较少,可能是因为在末次间冰期植被的反馈作用对华南地区降水的影响不显著(张琼和陈婕,2020)。在末次冰盛期,气候寒冷干燥、波动大,不利于石山苣苔属的生长发育,导致在该时期适生区面积丧失率高达11.71%。在末次冰盛期之后全球气候进入了温度急剧上升的冰消期,全球气候逐步进入了相对稳定的全新世暖期(况雪源等,2021),该时期适生区的北部和东北部急速向外缘扩增,适生区面积占比达到10.33%。当前时期至2050年,温度的逐渐升高促使了石山苣苔属适生区的扩展,使其适生区面积占比达到最大值14.29%,说明一定范围的气候变暖有利于石山苣苔属的扩散。但是,在2070年,适生区面积相对有所减少,说明全球气候变暖也可能致使该属物种实际生态位变窄。因为石山苣苔属对石灰岩土壤基质有特殊要求(Liu et al., 2017),它们难以跟踪适宜气候的北移。
总体看来,石山苣苔属在中国及中南半岛相对稳定的高适生区主要为中国西南部(广西、贵州、云南东南部)和越南北部。在这些地区加强保护力度,对自然保护区进行“禁止滥采滥挖”法律宣传。并且,石山苣苔属适生区整体向高纬度地区移动的现象,这与前人研究结果保持一致(Sekercioglu et al., 2008;方精云等,2018)。适宜生境中的收缩和扩张位于适宜区域的北部和东北部,这是物种对气候变化反应的敏感区域(Thuiller et al., 2005; Diamond et al., 2011),这些区域应该加强监测力度,针对性制定更好的指导保护策略。
综上所述,鉴于石山苣苔属大多数面临濒危的现状和具有较高的观赏价值,建议基于石山苣苔属自然群落在高适生区实施系统详细调查,摸清其致濒机制;基于潜在适生区,开展石山苣苔属引种、开发利用研究,为园林观赏植物的应用做好基础准备。
致谢 在这里非常感谢中国科学院昆明植物研究所税玉民研究员提供兔儿风叶石山苣苔(Petrocodon ainsliifolius)精确的经纬度地理信息;也非常感谢江西省中国科学院庐山植物园的助理研究员张涛为本文提供中国行政区矢量图。
参考文献:
AHMED SE, MCINERNY G, O′HARA K, et al., 2015. Scientists and software-surveying the species distribution modelling community [J]. Divers Distrib, 21(3): 258-267.
ALLEN AP, GILLOOLY JF, SAVAGE VM, et al., 2006. Kinetic effects of temperature on rates of genetic divergence and speciation [J].Proc Natl Acad Sci USA, 103(24): 9130-9135.
ANDERSON, RP, GONZALEZ I, 2011. Species-specific tuning increases robustness to sampling bias in models of species distributions:An implementation with MaxEnt [J]. Ecol Model, 222 (15): 2796-2811.
BARBOSA FG, SCHNECK F, 2015. Characteristics of the top-cited papers in species distribution predictive models [J]. Ecol Model, 313(C): 77-83.
BATEMAN BL, PIDGEON AM, RADELOFF VC, et al., 2016. The pace of past climate change vs. potential bird distributions and land use in the United States [J]. Glob Change Biol, 22: 1130-1144.
BIAN X, SHI FM, 2019. Predicting the potential distribution of Rhaphidophorinae (Orthoptera, Rhaphidophoridae) in China [J]. Zootaxa, 4683(1): 120-128.
BRADFORD AH, RICHARD F, HOWARD VC, et al., 2003. Energy, water, and broad-scale geographic patterns of species richness [J]. Ecology, 84(12): 3105-3117.
CEN HF, FU LF, WEN F, 2017. Petrocodon urceolatus sp. nov. (Gesneriaceae) from Zhangjiajie City, northwestern Hunan Province, China [J]. Nord J Bot, 35(3): 334-338.
COBOS ME, PETERSON AT, BARVE N, et al., 2019. Kuenm: an R package for detailed development of ecological niche models using MaxEnt [J]. Peer J, 7(1): e6281.
CONG MY, XU YY, TANG LY, et al., 2020. Predicting the dynamic distribution of Sphagnum bogs in China under climate change since the last interglacial period [J]. PLoS ONE, 15(4): e0230969.
CORY M, MATTHEW JS, JOHN AS, 2013. A practical guide to MaxEnt for modeling species distributions: what it does, and why inputs and settings matter [J]. Ecography, 36(10): 1058-1069.
DIAMOND SE, FRAME AM, MARTIN RA, et al., 2011. Species traits predict phenological responses to climate change in butterflies [J]. Ecology, 92(5): 1005-1012.
DOXFORD SW, FRECKLETON RP, 2012. Changes in the large-scale distribution of plants: Extinction, colonisation and the effects of climate [J]. J Ecol, 100(2): 519-529.
ELITH J, PHILLIPS SJ, HASTIE T, et al., 2011. A statistical explanation of MaxEnt for ecologists [J]. Divers Distrib, 17(1): 43-57.
FAN ZW, CAI L, YANG JW, et al., 2020. Petrocodon luteoflorus (Gesneriaceae), a new species from karst region in Guizhou, China [J]. PhytoKeys, 157: 167-173.
FANG JY, YODA K, 1991. Climate and vegetation in China V. Effect of climatic factors on the upper limit of distribution of evergreen broad leaf forest [J]. Ecol Res, 6: 113-125.
FANG JY, ZHU JL, SHI Y, 2018. The responses of ecosystems to global warming (in Chinese) [J]. Chin Sci Bull, 63(2): 136-140. [方精云, 朱江玲, 石岳, 2018. 生态系统对全球变暖的响应 [J]. 科学通报, 63(2): 136-140.]
FARASHI A, KABOLI M, KARAMI M, 2013. Predicting range expansion of invasive raccoons in northern Iran using ENFA model at two different scales [J]. Ecol Inf, 15: 96-102.
GE YZ, 2012. A study on phylogenetics of Petrocodon s.l. (Gesneriaceae) [D]. Nanning: Guangxi University. [葛玉珍, 2012. 廣义石山苣苔属(苦苣苔科)的系统发育研究 [D]. 南宁: 广西大学.]
GE YZ, XIN ZB, LI S, et al., 2020. Study on endangered degree and conserved levels for plants of Gesneriaceae in Guangxi [J]. Guihaia, 40(10): 1491-1504. [葛玉珍, 辛子兵, 黎舒, 等, 2020. 广西苦苣苔科植物濒危程度和优先保护序列研究 [J]. 广西植物, 40(10): 1491-1504.]
GRC, 2022. The Gesneriaceae Resource Centre. Royal Botanic Garden Edinburgh [EB/OL]. [2021-04-28].https://padme.rbge.org.uk/GRC.
HAWKINS BA, FIELD R, CORNELL HV, et al., 2003. Energy, water, and broad-scale geographic patterns of species richness [J]. Ecology, 84(12): 3105-3117.
HEWITT GM, 2004. Genetic consequences of climatic oscillations in the Quaternary. Philosophical transactions of the Royal Society of London [J]. Phil Trans Roy Soc B-Biol Sci, 359(1442): 183-195.
HIGGINS SI, LARCOMBEMJ, BEETON NJ, et al., 2020. Predictive ability of a process-based versus a correlative species distribution model [J]. Ecol Evolut, 10(20): 11034-11054.
HIGGINS SI, O′HARA RB, RMERMANN C, 2012. A niche forbiology in species distribution models [J]. J Biogeogr, 39: 2091-2095.
HOSMER DW, LEMESHOW S, 2000. Applied logistic regression [M]. New Jersey: John Wiley and Sons, Inc: 89-151.
JIANG C, TAN K, REN MX, 2017. Effects of monsoon on distribution patterns of tropical plants in Asia [J]. Chin J Plant Ecol, 41 (10): 1103-1112. [姜超, 谭珂, 任明迅, 2017. 季风对亚洲热带植物分布格局的影响 [J]. 植物生态学报, 41(10): 1103-1112.]
KHODOROVA NV, BOITEL CM, 2014. The role of temperature in the growth and flowering of geophytes [J]. Plants, 2(4): 699-711.
KUANG XY, HAN YC, WANG ZY, 2021. Dynamic down scaling simulation of millennial climate in China since the Last Glacial Maximum-Climate comparison of three typical periods [J]. Quat Sci, 41(3): 842-855. [況雪源, 韩跃超, 王志远, 2021. 末次冰盛期以来中国千年尺度气候的动力降尺度模拟——3个特征时期气候对比 [J]. 第四纪研究, 41(3): 842-855.]
LAMBERT AM, MILLER RJA, INOUYE DW, 2010. Changes in snowmelt date and summer precipitation affect the flowering phenology of Erythronium grandiflorum (glacier lily; Liliaceae) [J]. Am J Bot, 97 (9): 1431-1437.
LAURENT P, MOUILLOT F, YUE C, et al., 2018. FRY, a global database of fire patch functional traits derived from space-borne burned area products [J]. Sci Data, 5: 180132.
LI CR, PU F, GADAGKAR SR, et al., 2019. Petrocodon longitubus (Gesneriaceae), a new species from Guizhou, China [J]. Phytotaxa, 408(4): 1179-3163.
LI JY, CHANG H, LIU T, et al., 2019. The potential geographical distribution of Haloxylon across Central Asia under climate change in the 21st Century [J]. Agric For Meteorol, 275(C): 243-254.
LI L, CHEN JK, 2014. Influence of climate change on wild plants and the conservation strategies [J]. Biodivers Sci, 22(5): 549-563. [黎磊, 陈家宽, 2014. 气候变化对野生植物的影响及保护对策 [J]. 生物多样性, 22(5): 549-563.]
LI S, PAN B, XIN ZB, et al., 2020. Petrocodon ionophyllus, a new species of Gesneriaceae from the limestone areas of South China [J]. Rheedea, 30(1): 150-158.
LI ZL, QIN WH, WEN F, et al., 2020. Petrocodon wenshanensis, a new species of Gesneriaceae from southwestern China [J]. PhytoKeys, 157: 183-189.
LI ZY, 1996. The geographical distribution of the subfamily Cyrtandroideae Endl. Emend. Burtt (Gesneriaceae) [J]. Acta Phytotax Sin, 34(4): 341-360.
LIU YP, SHEN ZH, WANG QG, et al., 2017. Determinants of richness patterns differ between rare and common species: implications for Gesneriaceae conservation in China [J].Divers Distrib, 23(3): 235-246.
LU YB, HUANG YS, XU WB, et al., 2017. Floral evolutionary trend of Petrocodon (Gesneriaceae) and its significance in classification [J]. Guihaia, 37(10) : 1227-1239. [盧永彬, 黄俞淞, 许为斌, 等, 2017. 石山苣苔属(苦苣苔科)花形态演化及分类学意义 [J]. 广西植物, 37(10): 1227-1239.]
MATTEO P, MAURIZIO M, MARCO M, et al., 2020. Potential impact of climate change on the forest coverage and the spatial distribution of 19 key forest tree species in Italy under RCP4.5 IPCC trajectory for 2050s [J]. Forests, 11(934): 1-19.
MEROW C, SMITH MJ, SILANDER JA, 2013. A practical guide to MaxEnt for modeling species distributions: what it does, and why inputs and settings matter [J]. Ecography, 36(10): 1058-1069.
MUSCARELLA R, GALANTE PJ, SOLEY-GUARDIA M, et al., 2015. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MaxEnt ecological niche models [J]. Method Ecol Evolut, 5(11): 1198-1205.
MYERS N, MITTERMEIER RA, MITTERMEIER CG, et al., 2000. Biodiversity hotspots for conservation priorities [J]. Nature, 403(6772): 853-858.
NONG DX, HUANG BY, NONG SY, et al., 2021. Petrocodon albinervius, a new species of Gesneriaceae from limestone areas in southwestern Guangxi, China [J]. Taiwania, 66(2): 135-140.
PHILLIPS SJ, ANDERSON RP, DUDK M, et al., 2017. Opening the black box: an open-source release of MaxEnt [J]. Ecography, 40(7): 887-893.
PHILLIPS SJ, DUDK M, 2008. Modeling of species distributions with MaxEnt: new extensions and a comprehensive evaluation [J]. Ecography, 31 (2): 161-175.
PHILLIPS SJ, DUDK M, SCHAPIRE RE, 2021. MaxEnt software for modeling species niches and distributions (Version 3.4.1) [EB/OL]. [2021-11-28]. Available from url: http://biodiversityinformatics.amnh.org/open_source/maxent/.
POWO, 2021. Plants of the World Online [EB/OL]. [2021-11-28]. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet; http://www.plantsoftheworldonline.
QIAN MY, 2021. TRMM observation and regional model study of precipitation diurnal variations in summer over Indo-China Peninsula and South of China [D]. Nanjing: Nanjing University of Information Science and Technology. [钱满亿, 2021. 中南半岛地区植被与气候的联系及其未来预估 [D]. 南京: 南京信息工程大学.]
QIAO H, JORGE S, PETERSON AT, 2015. No silver bullets in correlative ecological niche modelling: insights from testing among many potential algorithms for niche estimation [J]. Method Ecol Evolut, 6(10): 1126-1136.
REN MX, 2015. The upper reaches of the largest river in Southern China as an “evolutionary front” of tropical plants: Evidences from Asia-endemic genus Hiptage (Malpighiaceae) [J]. Collect Bot, 34: e003.
SANDEL B, ARGE L, DALSGAARD B, et al., 2011. The influence of late quaternary climate-change velocity on species endemism [J]. Science, 334(6056): 660-664.
SEKERCIOGLU CH, SCHNEIDER SH, FAY JP, et al., 2008. Climate change, elevational range shifts, and bird extinctions [J]. Conserv Biol, 22(1): 140-150.
SILLERO N,2011. What does ecological modelling model?A proposed classification of ecological niche models based on their underlying methods [J]. Ecol Mod, 222: 1343-1346.
STEVEN JP, ROBERT PA, MIROSLAV D, et al., 2017. Opening the black box: an open-source release of MaxEnt [J]. Ecography, 40(7): 887-893.
STEVENS GC, 1989. The latitudinal gradient in geographical range: how so many species coexist in the tropics [J]. Amer Nat, 133(2): 240-250.
SU LY, PAN B, HONG X, et al., 2019. Petrocodon jiangxiensis (Gesneriaceae), a new species from Jiangxi, China [J]. Ann Bot Fennic, 56(4-6): 1797-2442.
SVENNING JC, SKOV F, 2007. Ice age legacies in the geographical distribution of tree species richness in Europe [J]. Glob Ecol Biogeogr, 16(2): 234-245.
TAO S, CHEN L, 1987. A review of recent research on the East Asian summer monsoon in China [M]//CHANG CP, KRISHNAMURTI TN. Monsoon meteorology. Oxford: Oxford University Press: 60-92.
THUILLER W, LAVOREL S, ARAJO MB, 2005. Niche properties and geographical extent as predictors of species sensitivity to climate change [J]. Glob Ecol Biogeogr, 14: 347-357.
VAZ UL, CUNHA HF, NABOUT JC, 2015. Trends and biases in global scientific literature about ecological niche models [J]. Brazilian J Biol, 75(4 Suppl. 1): 17-24.
WALTHER GR, POST E, CONVEY P, et al., 2002. Ecological responses to recent climate change [J]. Nature, 416(6879): 389-395.
WANG SP, MENG FD, DUAN JC, et al., 2014. Asymmetric sensitivity of first flowering date to warming and cooling in alpine plants [J]. Ecology, 95(12): 3387-3398.
WANG ZH, CHEN AP, FANG JY, 2004a. Relationship between species richness of seed plants and topography in Hunan Province [J]. Acta Geogr Sin, 59(6): 889-894. [王志恒, 陳安平, 方精云, 2004a. 湖南省种子植物物种丰富度与地形的关系 [J]. 地理学报, 59(6): 889-894.]
WANG ZH, CHEN AP, PIAO SL, et al., 2004b. Pattern of species richness along an altitudinal gradient on Gaoligong Mountains, Southwest China [J]. Biodivers Sci, 12(1): 82-88. [王志恒, 陈安平, 朴世龙, 等, 2004b. 高黎贡山种子植物物种丰富度沿海拔梯度的变化 [J]. 生物多样性, 12(1): 82-88.]
WARREN DL, MATZKE NJ, CARDILLO M, et al., 2021. ENMTools 1.0: an R package for comparative ecological biogeography [J]. Ecography, 44(4): 504-511.
WEN F, 2021. Gesneriaceae depends on the weak light at the mouth of the cave [J]. For Humankind, (8): 66-81. [溫放, 2021. 苦苣苔——依赖洞口弱光带 [J]. 森林与人类, (8): 66-81.]
WEN F, FU LF, XIN ZB, et al., 2022a. Endangered status and biodiversity conservation of Chinas Gesneriaceae [J].Guihaia.https://kns.cnki.net/kcms/detail/45.1134.Q.20220425.1936.007.html. [温放, 符龙飞, 辛子兵, 等, 2022a. 中国苦苣苔科植物濒危现状与多样性保护 [J]. 广西植物.https://kns.cnki.net/kcms/detail/45.1134.Q.20220425.1936.007.html.]
WEN F, WEI YG, FU LF, et al., 2022b. The Checklist of Gesneriaceae in China [EB/OL]. http://gccc.gxib.cn/cn/about-68.aspx. [温放, 韦毅刚, 符龙飞, 等, 2022b. 中国苦苣苔科植物名录 [EB/OL]. Available download from http://gccc.gxib.cn/cn/about-68.aspx.]
WU GX, DUAN AM, LIU YM, et al., 2013. Recent advances in the study on the dynamics of the Asian summer monsoon onset [J]. Chin J Atmos Sci, 37(2): 211-228. [吴国雄, 段安民, 刘屹岷, 等, 2013. 关于亚洲夏季风爆发的动力学研究的若干近期进展 [J]. 中国大气科学杂志, 37(2): 211-228.]
XIAO JH, DING X, CAI CN, et al., 2021. Simulation of the potential distribution of Phoebe bournei with climate changes using the maximum-entropy (MaxEnt) model [J]. Acta Ecol Sin, 41(14): 5703-5712. [肖建华, 丁鑫, 蔡超男, 等, 2021. 闽楠(Phoebe bournei, Lauraceae)地理分布及随气候变化的分布格局模拟 [J]. 生态学报, 41(14): 5703-5712.]
XIN ZB, DO TV, FU LF, et al., 2021. Petrocodon vietnamensis sp. nov. (Gesneriaceae) from karst caves in northern Vietnam [J]. Nord J Bot, 39(4): e02996.
XU X, ZHANG HY, XIE T, et al., 2018. Elevational pattern of seed plant diversity in Xishuangbanna and its mechanisms [J]. Biodivers Sci, 26(7): 678-689. [徐翔, 张化永, 谢婷, 等, 2018. 西双版纳种子植物物种多样性的垂直格局及机制 [J]. 生物多样性, 26(7): 678-689.]
ZHANG Q, CHEN J, 2020. Enhanced East Asian summer monsoon precipitation due to vegetation feedback during the last interglacial 127 ka [J]. Quaternary Sci, 40 (6): 1499-1512. [张琼, 陈婕, 2020. 末次间冰期127 ka时期植被反馈增强东亚夏季风降水的数值模拟研究 [J]. 第四纪研究, 40(6): 1499-1512.]
ZHANG RB, DENG T, FU LF, et al., 2019. Petrocodon tongziensis (Gesneriaceae), a new species from limestone areas in Guizhou, China based on morphological and molecular evidence [J]. Nord J Bot, 37(2): e01774.
ZHANG RL, FU LF, LI S, et al., 2018. Petrocodon asterocalyx, a new species of Gesneriaceae from Guangxi, China [J]. Phytotaxa, 343 (3): 259-268.
ZHANG RL, LI S, MACIEJEWSKI S, et al., 2020. Petrocodon rubiginosus, a new species of Gesneriaceae from Guangxi, China [J]. PhytoKeys, 157: 175-181.
ZHAO GH, CUI XY, SUN JJ, et al., 2021. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models [J]. Ecol Indic, 132: 108256.
ZHAO ZF, WEI HY, GUO YL, et al., 2016. Potential distribution of Panax ginseng and its predicted responses to climate change [J]. Chin J Appl Ecol, 27(11): 3607-3615. [赵泽芳, 卫海燕, 郭彦龙, 等, 2016. 人参潜在地理分布以及气候变化对其影响预测 [J]. 应用生态学报, 27(11): 3607-3615.]
MATAS L, LINARES JC, SNCHEZ M, et al., 2017. Contrasting growth forecasts across the geographical range of Scots pine due to altitudinal and latitudinal differences in climatic sensitivity [J]. Glob Change Biol, 23(10): 4106-4116.
ZHOU ZK, HUANG J, DING WN, 2017. The impact of major geological events on Chinese flora [J]. Biodivers Sci, 25(2): 123-135. [周浙昆, 黄健, 丁文娜, 2017. 若干重要地质事件对中国植物区系形成演变的影响 [J]. 生物多样性, 25(2): 123-135.]
ZHU GP, QIAO HJ, 2016. Effect of the MaxEnt models complexity on the prediction of species potential distributions [J]. Biodivers Sci, 24(10): 1189-1196. [朱耿平, 喬慧捷, 2016. MaxEnt模型复杂度对物种潜在分布区预测的影响 [J]. 生物多样性, 24(10): 1189-1196.]
ZHU H, 2013. Geographical elements of seed plants suggest the boundary of the tropical zone in China [J]. Palaeogeogr Palaeoclimatol, 386(15): 16-22.
ZHUO ZH, XU DP, PU B, et al., 2020. Predicting distribution of Zanthoxylum bungeanum Maxim. in China [J]. BMC Ecol, 20(1): 46.
(责任编辑 周翠鸣)
